
Abstract
The objective of this study was to investigate how the management practices of prescribed fire and understorey vegetation removal affect water and nutrient relations of old, yet prematurely declining Eucalyptus gomphocephala. Long unburnt sites were established in Yalgorup National Park, Western Australia, adjacent to frequently burnt state forest sites. Trees were allocated to vegetation clearing, prescribed fire or no prescribed fire treatments. Prescribed fire was achieved in only one long unburnt national park site so that the results were pseudoreplicated but analysed accordingly. Soil chemistry, plant nutrient availability and tree foliar carbon and nitrogen isotope ratio and nutrient concentration were investigated. No effects of vegetation clearing were found. Prescribed fire sites were associated with sky exposure and bare ground whereas no prescribed fire sites were associated with shrub and litter cover and litter depth. Foliar carbon isotope ratios were significantly more negative in prescribed fire, relative to no prescribed fire, treatments on long unburnt sites. Soil exchangeable Zn and Mn and plant available (estimated by charged resin beads) Mg were higher on prescribed fire, relative to no prescribed fire, long unburnt sites. Seedling bioassays indicated elevated P and Cu availability on prescribed fire, relative to no prescribed fire, treatments. In overstorey E. gomphocephala, foliar N levels were elevated (but not to excessive levels), and there was a trend toward elevated foliar Mn, in prescribed fire relative to no prescribed fire treatments on long unburnt sites. In the context of our large-scale pseudoreplicated case study, prescribed fire provided a pulse of water and N, (with some indications towards provision of elevated Mn, Cu and Mg) availability to E. gomphocephala in decline on sites with a history of a long absence of fire that may in part underpin observations of elevated tree health on sites that have a history of relatively frequent fire.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Many fire-dependent forests and woodlands of North America now contain more small and less large trees and less diverse understoreys compared to forests pre-European settlement due to changed management of fire exclusion and suppression, livestock grazing and timber extraction (Allen et al. 2002; Youngblood et al. 2006). The poor vigour and longevity of old trees within forests and woodlands of vegetation altered in structure by past European settlement is of critical importance given the increasing rarity and the unique habitat of old trees (Kolb et al. 2007). Fuel accumulation, from moribund trees, and greater density of mid-storey that ‘ladder’ fire into overstorey tree crowns, significantly increase the likelihood of severe wildfires which have been realised across extensive areas of North America (Westerling et al. 2006; Miller et al. 2008; Lutz et al. 2009). This has large ramifications for old trees that are threatened by drought and competition stress (Kolb et al. 2007) and that have inherently low growth rates due to relatively high respiratory demands per unit photosynthetic area (Sillett et al. 2010). Close et al. (2009) have highlighted the parallels in management history, vegetation structure and premature decline in health of old trees between some Australian eucalypt forests and woodlands and those of North America. Similar to North America, extensive wildfires have been realised in Australia (Shakesby et al. 2003; Mills 2005; Mitchell et al. 2006). We suggest that the altered vegetation structure and premature decline of old trees in some Australian forests and woodlands represents a significant threat to old trees in native forests and woodlands, analogous to the decline of isolated and scattered remnant trees within the modified agricultural landscapes of temperate Australia (Manning et al. 2006; Davidson et al. 2007; Close et al. 2008). Whilst overstorey-understorey interactions have been shown to be important in many fire-prone forest systems (Dodson et al. 2008) there has been very little work investigating the consequences of altered vegetation structure on the ecophysiology of old trees in premature decline.
In response to historical woodland and forest management and the consequent elevation of fire risk, managers have developed prescribed fire practices in recent decades. In addition, contemporary effort in North America has been focussed on fire surrogate treatments such as mechanical thinning to change forest structure and enable the use of prescribed fire to restore woodland and forest ecosystems. Major policy (Stephens and Ruth 2005) and research (fire and fire surrogate research program; Stephens et al. 2009) initiatives are building scientific understanding and underpinning landscape scale forest management of prescribed fire and mechanical thinning in North America. However the practice of prescribed burning is socially contentious in Australia (Bell and Oliveras 2006). There has been important research into the development of prescribed fire protocols (McCaw et al. 1997) however, while mechanical thinning has been practiced as a silvicultural tool for improving harvestable yield (Reid 2006) the practice has not been investigated or effected as a fire surrogate. Recently an argument for research into the use of mechanical thinning as a fire surrogate in historically fire-prone woodlands and forests has been proposed (Archibald et al. 2010).
In Australia, studies indicate that in the long absence of fire: 1) a limited range of shade tolerant woody understorey/midstorey species thrive; 2) dominant overstorey eucalypts prematurely decline and; 3) there is a lack of recruitment of the shade intolerant overstorey eucalypts (Ellis 1971, 1985; Withers and Ashton 1977; Lunt 1998; Werkman et al. 2008; reviewed in Close et al. 2009). Furthermore, recovery of premature decline in overstorey E. delegatensis forest has been reported in response to prescribed fire (Ellis et al. 1980). We were interested in the ecophysiology associated with the premature decline of overstorey eucalypts with a history of a long absence of fire and how this might be affected by prescribed fire and understorey/midstorey removal (a proxy for mechanical thinning). We hypothesised that with a long absence of fire the increased development of a woody understorey/midstorey: 1. competes with overstorey eucalypts for soil water and; 2. alters nutrient cycling processes that may lead to phosphorus and/or cation deficiency or excessive uptake of nitrogen (Close et al. 2009).
In a recent study we investigated overstorey E. gomphocephala in Western Australia and Eucalyptus delegatensis in Tasmania on sites of histories of either a long absence of fire or frequent fire where overstorey tree decline was apparent (Close et al. 2011). We found significantly developed under- and mid-storey vegetation and decreased C isotope discrimination (increased C13:C12 isotope ratio) on sites of long unburnt history, consistent with greater water limitation (Farquhar et al. 1982; Farquhar and Richards 1984) in overstorey trees on sites of long unburnt history in both E. gomphocephala in Western Australia and in E. delegatensis in Tasmania. Further that foliar Zn, Cu and P concentrations were negatively correlated with tree decline severity in overstorey E. gomphocephala in Western Australia and in E. delegatensis in Tasmania, respectively (hypothesised to be due to deficiency of these nutrients in trees in decline), and that there was no evidence for luxury nitrogen uptake as a result of soil N accumulation in the absence of fire (Close et al. 2011). In light of these findings we posed the following hypotheses towards the question: ‘can land management redress the decline of old, overstorey eucalypts?’
-
Hypothesis 1
That water stress in old, overstorey trees, induced by competition with understorey/midstorey developed in the long absence of fire, can be reduced through prescribed fire and/or vegetation clearing treatments. Whilst responses have varied as a result of thinning understory vegetation to reduce within-stand competition on remaining trees (see Kolb et al. 2007 for a review), McDowell et al. (2003) elegantly demonstrated that stand density reductions that resulted in increased growth of old Pinus ponderosa trees effected increased stomatal conductance during the period that stand leaf area was not fully re-established. Diffusion of CO2 into leaves is a function of stomatal conductance, thus increased stomatal conductance led to increased photosynthetic assimilation. This is expressed in the C13:C12 isotope ratio, given that C13 is discriminated against due to relatively low diffusivity into stomata and low reactivity with rubisco (Farquhar et al. 1982; Farquhar and Richards 1984). Warren et al. (2001) recommend that the C13:C12 isotope ratio is a useful indicator of drought stress in seasonally dry climates where variation in other environmental factors can be accounted for. Thus we expected in this study that thinning and prescribed fire treatments would increase the availability of soil water to trees that would consequently be reflected in more negative foliar C13:C12 isotope ratio signatures.
-
Hypothesis 2
That pH-dependent micronutrients (Fe, Mn, Zn, Mg and Cu) may become deficient due to being plant un-available in soil and/or being locked up in biomass in the long absence of fire and that micro-nutrients increase in plant-availability due to mineralisation following fire (Raison et al. 1985; Tomkins et al. 1991; Khanna et al. 1994).
E. gomphocephala woodlands and forests are restricted to the coastal plain of south-west Australia. Approximately 27% of the original distribution remains and of this only 31% is reserved (CALM 2003). Clearing, for urban expansion, is an increasing threat thus management that maintains the composition and health of reserved E. gomphocephala woodlands and forests is critical. However the health of E. gomphocephala trees has declined rapidly across Yalgorup National Park (Longman and Keighery 2002) that represents 50% of the area of reserved E. gomphocephala woodlands. Thus the research question ‘can land management redress the decline of old, overstorey eucalypts?’ is particularly relevant to the management of E. gomphocephala woodlands and forests. The objective of the current study was to investigate how the management practices of prescribed fire and understorey removal affect water and nutrient relations of old, overstorey E. gomphocephala. We allocated half of each long unburnt national park and recently burnt state forest site (Close et al. 2011) to prescribed fire treatment and prior to burning we slashed and dropped the understory vegetation to a radius of 20 m from the base of half the study trees.
Methods
Experimental sites
Sites (ranging 50 to 155 ha in extent) were established in woodland of different fire history approximately 130 km south of Perth. Four sites were in Yalgorup National Park (had not been burnt since at least 1972, considered ‘long unburnt’ given that historical grasstree data indicate that fire occurred every 5–10 years; Ward et al. 2001; 115°39′00″ E, 32°50′00″ S), and six sites were within adjacent state forest (subject to prescribed fire every 5–15 years for the past 6 decades, considered ‘frequently burnt’; 115°43′00″ E, 32°55′00″ S). Sites comprised Eucalyptus gomphocephala dominated woodland on the coastal Yoongarillup Plain, of grey-brown sandy soils with limestone bedrock generally within 1 m of the soil surface. The climate is Mediterranean with less than 10% of the mean annual rainfall of 882 mm falling between November and March and a mean annual temperature of 17.7°C.
Vegetation clearing and prescribed fire
Ten trees were randomly selected at each site and permanently labelled as a basis for subsequent sampling for foliage, and for soil and soil solution chemistry from within the dripline of the individual rees. Each of the ten trees was then randomly allocated to understorey vegetation clearing or no vegetation clearing treatments. Clearing treatment consisted of mechanical understorey/midstorey removal (primarily Agonis flexuosa and Spyridium globulosum and excluding Xanthorrea preissii) and stump treatment with glyphosate herbicide to prevent re-sprouting within a 20 m radius of the treatment tree. Subsequent to drying of the slashed vegetation, one of the four long unburnt and three of the six frequently burnt sites were subjected to prescribed fire as weather and resources allowed. Decisions on which sites were subjected to prescribed fire were made by the fire management authority (Department of Environment and Conservation, Western Australian State Government) according to internal protocols of flora, fauna, indigenous and ecosystem values, asset protection and public safety. In all cases the edges were ignited to 30 m on the wind-lee aspects of sites (‘back-burned’) at 1130–1200 h before core ignition at 1300 h via multiple ignitions from drip-torch every 30 m along transects. Prescribed fire was conducted in April 2007 (one frequently burnt site) and November 2007 (two frequently burnt and the long unburnt site) when surface moisture content of litter was 12.5–14.0%.
Vegetation survey
Ten (2 × 2 m) understorey/midstorey vegetation plots were randomly located and permanently labelled at each site prior to prescribed fire (Close et al. 2011) and surveyed approximately three months after prescribed fire. The number of surveys and area surveyed were necessarily limited by time and resource given the difficulty of traversing and surveying extremely dense understorey that was developed on long unburnt sites. Cover % of understorey/midstorey vegetation guilds were assessed by the Domin Scale (Mueller-Dombois and Ellenberg 1974). Understorey/midstorey cover estimates of vegetation guilds were transformed in PATN (Belbin 2004) by [(Value-Minimum Value)/Range of Values]. Ordinations were applied using a semi-strong-hybrid multidimensional scaling based on a Bray-Curtis dissimilarity coefficient. Vectors, tested using a Monte-Carlo approach, were considered significant at p < 0.01.
Soil and plant nutrient and isotope analysis
Soil samples (4 sub-samples, 2 m from the trunk and bulked per tree) were collected (from the top 5 cm of the soil) in January 2008. Samples were stored in sealed plastic bags, before oven drying at 110°C. Soils from individual tree samples from the same prescribed fire and understorey clearing treatment combinations were bulked so that there were three replicate samples per treatment combination. Soil carbon, nitrogen and phosphorus analysis was as described in Close et al. (2008). Soils were extracted in diethylenetriaminepentaacetic acid (DTPA) for two hours prior to analysis by atomic absorption spectroscopy for Cu, Zn, Mn, and Fe (Rayment and Higginson 1992). Soils were extracted in 0.1M BaCl2 and 0.1M NH4Cl for analysis of exchangeable Ca, Mg, Na and K (Rayment and Higginson 1992).
Soil solution nutrient availability was estimated using cation- and anion-exchange resins (Amberlite, Sigma-Aldrich). Pre-washed bags made of stocking nylon were filled with 5 g (saturated weight) of resin beads. Resin bags were buried at 5 cm depth approximately 1 m apart in pairs, four pairs under the dripline of each tree at three randomly selected sites per fire history and collected approximately six months later (spring 2007 for one frequently burnt site and winter 2008 for the remainder of the sites). Anions and cations were extracted and analysed as described by Giblin et al. (1994). One foliar sample per tree (30–40 young, fully expanded leaves) was collected from the outer crown on a northerly aspect early July 2008. A total of 15 trees were sampled for each treatment combination except for the prescribed fire treatment on long unburnt sites in which only five trees were sampled as prescribed fire was achieved on only one long unburnt site. Leaf samples were dried at 40°C before sub-sampling for nutrition and 13C and 15N isotope analyses as described in Close et al. (2008).
Seedling bioassay
Seeds of E. gomphocephala were collected from trees within Yalgorup National Park. Seeds were germinated on 20 December 2006 and raised in ‘hiko’ trays in a potting mix with no fertiliser added in a controlled temperature glasshouse (15–25°C) and watered as required. On 3 May 2007, the seedlings were planted four per tree at N-S-E-W under the dripline of the crown of permanently labelled trees within a frequently burnt site, half of which had been subjected to prescribed fire in Autumn 2007. It was not possible to use seedling bioassays at other sites given that these were subjected to prescribed fire in spring 2007 and the seedlings would not have survived the dry, Mediterannean summer. At least one seedling per tree (many had been browsed by kangaroo) was harvested for foliar nutrient analysis on 26 July 2007.
Analysis
Prescribed fire was achieved in three frequently burnt, but only one long unburnt site (three being the original objective but weather and resources did not allow) resulting in an imbalanced data set. In addition, the sampling structure within the experiment consisted of multiple subsamples (individual trees) per experimental unit (site) resulting in pseudoreplication. Due to imbalance we used least square means and we addressed the pseudoreplication issue using linear mixed models in which the subsampling were modelled using random effects (Millar and Anderson 2004). Inference then refers to the site level effects and their interactions rather than subsamples. Site fire history, prescribed fire and understorey clearing were tested as factors in the analysis of soil and tree foliage nutrient status and tree foliage carbon and nitrogen isotope data. There were no effects of understorey/midstorey clearing found so this was dropped from the model. Surviving bioassay seedlings were pooled per tree they were planted around, this being the unit of replication. Inference was conducted using Type 3 analyses of variance and REML (restricted maximum likelihood) estimation using the Mixed Procedure in SAS version 9.2. Beginning with the model outcome = prescribed fire + site fire history + prescribed fire* site fire history, we dropped the interaction if not significant and only then considered the additive model. Where appropriate, least square means and Tukey adjusted P-values (Westfall et al. 1999) of pairwise comparisons were reported. Results were considered significantly different at the p < 0.05 level.
Results
Vegetation
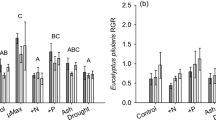
Pattern analysis indicated that prescribed fire had an over-riding effect on understorey vegetation (Fig. 1). Vegetation plots that were not subjected to prescribed fire in either frequently burnt or long unburnt were similarly associated with shrub and litter cover and litter depth. Conversely vegetation plots that had been subjected to prescribed fire in either frequently burnt or long unburnt were similarly associated with bare ground cover and sky exposure.

Centroids (bars represent one standard error) of 3-dimensional ordinations of site attributes (□ = long unburnt not subjected to prescribed fire, ■ = frequently burnt not subjected to prescribed fire, ○ = long unburnt subjected to prescribed fire, ● = frequently burnt subjected to prescribed fire) with significant (p < 0.01) vectors and r2 values shown for E. gomphocephala woodland, Western Australia (overall stress = 0.137)
Soil chemistry
Soil organic carbon was significantly higher in long unburnt/no prescribed fire soils at 4% relative to other treatments that were around 2% (Table 1a; fire history x prescribed fire interaction F1,22 = 12.91, p = 0.0016). Soil pH was significantly lower (around 7.0) in frequently burnt/no prescribed fire soils than in long unburnt and frequently burnt sites subject to prescribed fire (around 8.0; Table 1a; fire history x prescribed fire interaction F1,22 = 12.99, p = 0.0016), but was not different to long unburnt/no prescribed fire soils due to high variability of the data. There was a significant effect of prescribed fire but not of fire history with soil zinc (Table 1a; F1,21 = 6.26, p = 0.0207) and manganese (Table 1a; F1,21 = 14.76, p = 0.0009) being higher in soils of prescribed fire sites. Soil magnesium (fire history effect; F1,22 = 18.58, p = 0.0498) and iron (fire history effect; F1,21 = 9.881, p = 0.0049) were significantly higher in long unburnt than frequently burnt sites. There were no significant effects of fire history or prescribed fire on soil phosphorus or copper.
Plant nutrient availability
Soil solution nutrition
Soil solution nitrate was significantly higher in prescribed fire than no prescribed fire sites (Table 1b; F1,49 = 4.21, p = 0.0456) with no effect of fire history. Soil solution ammonium was highest in long unburnt sites, intermediate in frequently burnt/prescribed fire and lowest in frequently burnt/no prescribed fire (Table 1b; fire history x prescribed fire interaction effect: F1,49 = 5.53, p = 0.0227). Soil solution Mg was higher in long unburnt/prescribed fire soils, than in soils of other sites (Table 1b; fire history x prescribed fire interaction F1,49 = 11.67, p = 0.0013). Soil solution copper was highest in long unburnt/no prescribed fire, intermediate in frequently burnt sites, and lowest in long unburnt/prescribed fire (fire history x prescribed fire interaction effect: F1,31 = 5. 71, p = 0.0231). There were no significant differences in levels of soil solution iron, phosphorus or manganese.
Seedling bioassay
Seedling foliar total N was similar but P (F1,10 = 4.00, p = 0.0733; Table 1c) and Cu (F1,10 = 4.37, p = 0.0632; Table 1c) showed a trend towards being higher in seedlings planted into a site subjected to prescribed fire than a no prescribed fire site. There were no significant differences in seedling foliar K, Zn or Mn.
Tree nutrition
Foliar N was highest in trees on long unburnt/prescribed fire, intermediate in frequently burnt/no prescribed fire and lowest in long unburnt/no prescribed fire and frequently burnt/prescribed fire (Table 1d; fire history x prescribed fire interaction effect: F1,94 = 56.36, p < 0.0001) sites. Foliar P was highest in trees on long unburnt/prescribed fire and frequently burnt/no prescribed fire, intermediate in trees of frequently burnt/prescribed fire and lowest in trees on long unburnt/no prescribed fire sites (Table 1d; F1,94 = 29.42, p < 0.0001). There was a significant effect of fire history (F1,94 = 11.62, p = 0.0010), but not of prescribed fire, with magnesium being lower in trees of frequently burnt sites. There was a significant interaction effect of fire history x prescribed fire for copper (F1,94 = 7.64, p = 0.0069) but no pair-wise comparisons indicated significant difference after Tukey adjustment. There were no significant differences between treatments of tree foliar Mn, Zn or Fe.
Tree foliar isotope ratios
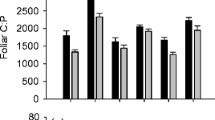
There was a significant fire history x prescribed fire interaction effect where foliage of long unburnt/prescribed fire treatment trees had more negative δ13C than foliage of other treatments (Table 1e; F1,92 = 5.64, p = 0.0196). δ15N was most negative in trees on frequently burnt/prescribed fire and long unburnt/no prescribed fire, intermediate in trees on frequently burnt/no prescribed fire and least negative in trees on long unburnt/prescribed fire sites (Table 1e; fire history x prescribed fire effect, F1,92 = 36.75, p < 0.0001).
Discussion
In E. gomphocephala woodlands of south-west Australia prescribed fire impacted significantly on understorey structure: the soil surface litter layer and understorey shrubs were combusted, whereas non-prescribed fire sites, irrespective of fire history, were characterised by relatively high levels of shrub and litter cover and litter depth and significantly elevated soil organic carbon. Thus prescribed fire can alter the vegetation structure that is associated with premature decline of overstorey eucalypts (Ellis 1971; Werkman et al. 2008; Close et al. 2011) despite the fact that prescribed fire on frequently burnt sites was conducted in autumn on one site and spring on the other three sites.
Consistent with our first hypothesis we detected significantly increased carbon isotope discrimination (δ13C decreased) in overstorey trees where understorey/midstorey had been subjected to prescribed fire, relative to no prescribed fire, on sites with a history of long absence of fire and significantly developed understorey/midstorey cover. Given that all our study trees were old, emergent trees in the woodland overstorey, increased interception of radiation leading to increased rates of photosynthesis that may underpin increased δ13C in trees as a result of our treatments is unlikely. However, we acknowledge that the significantly elevated foliar N observed in trees on long unburnt sites that were subjected to prescribed fire may mask in part the result, given that increased foliar N will support faster rates of photosynthesis and increased δ13C (Warren et al. 2001). The analogous result of increased N one and two years after thinning and prescribed fire, but not in thinned only treatments, after a long history of no fire was reported in P. ponderosa (Feeney et al. 1998; Wallin et al. 2004).
The result of significantly decreased δ13C in long unburnt trees of prescribed fire treatment at levels similar to that found in non water-limited irrigated Corymbia calophylla and E. marginata (−28.8‰ to −29.1‰) in Western Australia (Grigg et al. 2009), may in part explain the empirical observation of reversed crown decline in E. delegatensis in response to understory/midstorey felling and prescribed fire (Ellis et al. 1980). Particularly given that we have recently reported δ13C consistent with water limitation in E. delegatensis on sites with a history of long absence of fire (Close et al. 2011). The implied increase in soil water-availability post prescribed-fire could have significant implications for the maintenance of positive carbon balance in old trees of early successional status that depend on frequent fire for the maintenance of an ecological disclimax (Jackson 1968). Particularly given that leaf-area-based sap flow rates decrease as tree size increases so that water supply to foliage becomes disproportionately limited in older trees relative to younger trees under conditions of soil-water deficit (Hubbard et al. 1999; Ryan et al. 2000; Sala et al. 2001).
In contrast to our observations on sites with a history of long absence of fire, we found no change in tree foliar δ13C between prescribed fire and no prescribed fire treatments on sites of frequent fire history. This is consistent with the broader hypothesis that contemporary fire management that approximates pre-European fire frequency effects a vegetation structure where water-availability is balanced between the understorey and over-storey eucalypts and premature eucalypt decline is less likely to occur (Close et al. 2009).
Consistent with hypothesis 2, prescribed fire had significant effects on soil chemistry and estimates of plant nutrient-availability. Soil pH was significantly elevated by prescribed fire on frequently burnt sites relative to soils of no prescribed fire sites (around 8 and 7 pH units, respectively), likely due to the release of base cations (Raison 1979). In contrast, pH was around 8 pH units in soils of both long unburnt treatments. This may indicate an inherent difference between the sites given that long unburnt sites were closer (500–1000 m) to the ocean and that carbonates of calcareous soils are stable up to 1000°C (Rabenhorst 1988) and buffer pH against fire-induced change. This could have important implications for the availability of pH-dependent micronutrients (Zn, Mn, Fe, Mg and Cu) given that lack of plant-availability when soil pH is higher than 6.5 (Lambers et al. 1998), underpins the ‘lime-induced chlorosis’ phenomenon (Bavaresco et al. 1999; Fernández and Ebert 2005) that has recently been demonstrated in eucalypts (Czerniakowski et al. 2006; Grigg et al. 2009; see Parsons and Uren 2007 for review). Whilst we found elevated soil Zn and Mn (irrespective of fire history), consistent with the release of these plant nutrients through the combustion of biomass (Tomkins et al. 1991), we did not find elevated plant-available (captured on anionic resin beads) Zn or Mn. This is consistent with findings of easily reducible Mn increasing after fire with no concomitant change in exchangeable Mn (Gonzalez Parra et al. 1996). We also found six-fold higher plant-available Mg in soils of sites subjected to prescribed fire, relative to no prescribed fire sites that were long unburnt. Thus prescribed fire may alter the soil environment temporarily to enable plant uptake of a pulse of Mn, Zn and Mg that in the longer-term becomes unavailable due to ‘lime-induced chlorosis’.
Consistent with the notion that prescribed fire may alter the soil environment temporarily to enable plant uptake of a pulse of Mn, Zn and Mg, we found a trend toward lower levels of Mn, but not of Zn or Mg, in trees of long unburnt/no prescribed fire, relative to trees of frequently burnt no prescribed fire sites. This is significant given that in our previous study involving a greater number of trees (Close et al. 2011) we found a positive correlation of E. gomphocephala health with increasing foliar Mn. Further, a study in parkland irrigated with high pH bore water found significant deficiency in foliar Mn of E. marginata and C. calophylla trees in poor health relative to healthy trees in adjacent, non-irrigated bushland (Grigg et al. 2009). Increased plant-uptake of Mn associated with fire has been reported. Mn content of X. preissii annual stem bands decreased linearly with time since fire from 1930–1980 in a long unburnt E. marginata forest whilst only a slight decrease in Mn was found in annual stem bands of a X. preissii in forest that had been regularly burnt during the same period (Swanborough et al. 2003). Thus prescribed fire may provide a pulse of Mn to declining overstorey trees in the context of our previous study where foliar Mn and Zn were positively correlated to tree health (Close et al. 2011), despite the limitation of this study where one frequently burnt site was burnt in autumn and the others were burnt in spring.
In addition to our previous findings of correlation between tree health and foliar Mn, we also found significant correlation between tree health and foliar Cu (Close et al. 2011). Whilst we did not find evidence for elevated Cu availability with the anion-resin bead method, nor elevated Cu in the foliage of trees, of the prescribed fire treatments, we did find a trend towards higher uptake of Cu in bioassay seedlings planted on prescribed fire, compared with no prescribed fire, sites in the current study. Similarly, we have detected no effect of prescribed fire on levels of soil P and whilst we found a trend towards elevated P uptake in bioassay seedlings, foliar P of overstorey E. gomphocephala was actually lower in the prescribed fire treatment on both frequently burnt and long unburnt sites. The latter result may be due to elevated pH in ash reducing the availability of P and Cu over a longer time-step than that assessed with the bioassay seedlings. Bioassay seedlings were harvested only six weeks following prescribed fire whereas the foliage of overstorey E. gomphocephala trees was harvested six months following prescribed fire. This may be a reflection of only a short window of elevated P and Cu availability. In contrast increased soil pH and consequent plant-availability and uptake of P after fire on acid soils has been reported up to nine years post-fire (Humphreys and Lambert 1965).
No prescribed fire sites, relative to prescribed fire sites (irrespective of site history), contained significantly elevated levels of soil nitrate, indicative of denitrifying soil conditions in the long absence of fire. This finding is consistent with significantly lower foliar δ15N in trees of no prescribed fire, relative to prescribed fire sites with a history of being frequently burnt, given that fractionation by mycorrhizae post nitrate uptake occurs prior to transfer to trees, leading to more negative δ15N (Högberg 1997) and that history of being frequently burnt is favourable to mycorrhizae of eucaypts (B. Horton unpub). δ15N could be lower in the soil of prescribed fire sites with a history of being long unburnt, given that fire can stimulate biological N fixation by understorey legumes (Hamilton et al. 1991). High soil ammonium in long unburnt, relative to frequently burnt, sites is consistent with an expectation of less negative δ15N in foliage of trees on long unburnt sites as ammonium is the preferred N source for temperate eucalypts (Garnett and Smethurst 1999) and is undepleted in δ15N relative to mycorrhizae-provided nitrate-N. However this was observed in trees subjected to prescribed fire, and not in trees subjected to no prescribed fire, on sites with a history of being long unburnt. Cook (2001) argues that, within a particular rainfall regime, foliar δ15N may indicate historical fire frequencies. Further, that the relative importance of ammonium and nitrate ions for plant uptake appears to control the effects of fire on foliar δ15N values. It is difficult to clearly interprete our results of δ15N. Future studies that incorporate invstigation of the contribution of ash are warranted (Cook 2001), but were outside the scope of the current study.
Conclusion
Given the logistical challenge of replicating prescribed fire, results of this study are pseudoreplicated yet provide key information in the context of a large-scale case-study. Thus, on sites with a history of long absence of fire and decline in the E. gomphocephala overstorey, results of this study indicated that prescribed fire temporarily created an environment of elevated soil water-, and N-availability. Prescribed fire may also temporarily increase the availability of Mg, Mn and/or Cu, although the results of this study were equivocal in the case of these plant nutrients, and this warrants further investigation. Low-to-moderate intensity surface fire may therefore effect a fire disclimax (sensu Jackson 1968) that may serve to perpetuate the health and longevity of old, emergent E. gomphocephala trees and perhaps early successional tree species in general, in part through the provision of water and nutrient ‘pulses’ of availability soon after low-moderate intensity surface fire.
References
Allen CD, Savage M, Falk DA, Suckling KF, Swetnam TW, Schulke T, Stacey PB, Morgan P, Hoffman M, Klingel JT (2002) Ecological restoration of southwestern ponderosa pine ecosystems: a broad perspective. Ecol App 12:1418–1433
Archibald RD, Bradshaw J, Bowen BJ, Close DC, McCaw L, Drake PL, St. Hardy GEJ (2010) Understorey thinning and burning trials are needed in conservation reserves: the case of Tuart (Eucalyptus gomphocephala D.C.). Ecol Man Rest 11:108–112
Bavaresco L, Giachino E, Colla R (1999) Iron chlorosis paradox in grapevine. J Plant Nutr 22:1689–1597
Belbin L (2004) PATN Technical Reference. CSIRO Division of Wildlife and Ecology, Canberra
Bell T, Oliveras I (2006) Perceptions of prescribed burning in a local forest community in Victoria, Australia. Env Manage 38:867–878
CALM (Department of Conservation and Land Management), Tuart Response Group, and Ecoscape, 2003. An atlas of tuart woodlands on the Swan Coastal Plain in Western Australia. Department of Conservation and Land Management, Perth
Close DC, Davidson NJ, Watson T (2008) Health of remnant woodlands in fragments under distinct grazing regimes. Biol Cons 141:2395–2402
Close DC, Davidson NJ, Johnson DW, Abrams MD, Hart SC, Lunt ID, Archibald RD, Horton B, Adams MA (2009) Premature decline of Eucalyptus and altered ecosystem processes in the absence of fire in some Australian forests. Bot Rev 75:191–202
Close DC, Davidson NJ, Swanborough P (2011) Fire history and understorey vegetation: water and nutrient relations of overstorey Eucalyptus delegatensis and E. gomphocephala. For Ecol Manage 262:208–214
Cook GD (2001) Effect of frequent fires and grazing on stable nitrogen isotope ratios of vegetation in northern Australia. Aust Ecol 26:630–636
Czerniakowski B, Crnov R, Smith IW, Luck JE (2006) Soil properties associated with the tree decline ‘Mundulla Yellows’. Plant Soil 285:197–206
Davidson NJ, Close DC, Battaglia M, Churchill K, Ottenschlarger M, Watson T, Bruce J (2007) Eucalypt health and agricultural land management within bushland remnants in the Midlands of Tasmania, Australia. Biol Conserv 139:439–446
Dodson EK, Peterson DW, Harrod RJ (2008) Understorey vegetation response to thinning and burning restoration treatments in dry conifer forests of the eastern Cascades, USA. For Ecol Manage 255:3130–3140
Ellis RC (1971) Dieback of alpine ash as related to changes in soil temperature. Aust For 35:152–163
Ellis RC (1985) The relationships among eucalypt forest, grassland and rainforest in a highland area in north-eastern Tasmania. Aust J Ecol 10:297–314
Ellis RC, Mount AB, Mattay JB (1980) Recovery of Eucalyptus delegatensis from high altitude dieback after felling and burning the understorey. Aust For 43:29–35
Farquhar GD, Richards RA (1984) Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust J Plant Physiol 11:539–552
Farquhar GD, O’Leary MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust J Plant Physiol 9:121–137
Feeney SR, Kolb TE, Wagner MR, Covington WW (1998) Influence of thinning and burning restoration treatments on pre-settlement ponderosa pines at the Gus Pearson Natural Area. Can J For Res 28:1295–1306
Fernández V, Ebert G (2005) Foliar iron fertilization: a critical review. J Plant Nutr 28:2113–2124
Garnett TP, Smethurst PJ (1999) Ammonium and nitrate uptake by Eucalyptus nitens: effects of pH and temperature. Plant Soil 214:133–140
Giblin AE, Laundre JA, Nadelhoffer KJ, Shaver GR (1994) Measuring nutrient availability in arctic soils using ion-exchange resins—a field-test. Soil Sci Soc Am J 58:1154–1162
Gonzalez Parra J, Cala Riveroand V, Iglesias Lopez T (1996) Forms of Mn in soils affected by a forest fire. Sci Total Environ 181:231–236
Grigg A, Close DC, Lambers H, Ruthroff KX, Dixon KW (2009) Ecophysiology of Eucalyptus marginata and Corymbia calophylla in decline in an urban parkland. Austral Ecol 34:499–507
Hamilton SD, Lawrie AC, Hopmans P, Leonard BV (1991) Effects of fuel-reduction burning on a Eucalyptus obliqua forest ecosystem in Victoria. Aust J Bot 39:203–217
Högberg P (1997) Tansley Review No. 95: 15N natural abundance in soil-plant systems. New Phytol 137:179–203
Hubbard RM, Bond NJ, Ryan MG (1999) Evidence that hydraulic conductance limits photosynthesis in old Pinus ponderosa trees. Tree Physiol 19:165–172
Humphreys FR, Lambert MJ (1965) An examination of a forest site which has exhibited the ash-bed effect. Aust J Soil Res 3:81–94
Jackson WD (1968) Fire, air, water and earth—an elemental ecology of Tasmania. Proc Ecol Soc Aust 3:9–16
Khanna PK, Raison RJ, Falkiner RA (1994) Chemical properties of ash derived from Eucalyptus litter and its effect on forest soils. For Ecol Manage 66:107–125
Kolb TE, Agee JK, Fulé M, McDowell NG, Pearson K, Sala A, Waring RH (2007) Perpetuating old ponderosa pine. For Ecol Manage 249:141–157
Lambers H, Chapin FSIII, Pons TL (1998) Plant physiological ecology. Springer-Verlag, New York
Longman VM, Keighery BJ (2002) Tuart Issues. In: Keighery BJ, Longman VM (eds) Tuart (Eucalyptus gomphocaphala) and Tuart Communities. Wildflower Society of Western Australia, Nedlands, pp 292–330
Lunt ID (1998) Allocasuarina (Casuarinaceae) invasion of an unburnt coastal woodland at Ocean Grove, Victoria: Structural changes 1971–1996. Aust J Bot 46:649–656
Lutz JA, van Wagtendonk JW, Thode AE (2009) Climate, lightning ignitions and fire severity in Yosemite National Park, California, USA. Int J Wildland Fire 18:765–774
Manning AD, Fischer J, Lindenmayer DB (2006) Scattered trees are keystone structures—implications for conservation. Biol Cons 132:311–321
McCaw WL, Smith RH, Neal JE (1997) Prescribed burning of thinning slash in regrowth stands of karri (Eucalyptus diversicolor) 1. Fire characteristics, fuel consumption and tree damage. Int J Wildland Fire 7:29–40
McDowell N, Brooks JR, Fitzgerald A, Bond BJ (2003) Carbon isotope discrimination and growth response of old Pinus ponderosa trees to stand density reductions. Plant Cell Environ 26:631–344
Millar RB, Anderson MJ (2004) Remedies for pseudoreplication. Fish Res 70:397–407
Miller JD, Safford HD, Crimmins M, Thode AE (2008) Quantitative evidence for increasing forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA. Ecosystems 12:16–32
Mills GA (2005) On the subsynoptic-scale meteorology of two extreme fire weather days during the Eastern Australian fires of January 2003. Aust Met Mag 54:265–290
Mitchell RM, O’Brien DM, Campbell SK 2006. Characteristics and radiative impact of the aerosol generated by the Canberra firestorm of January 2003. J Geophys Res.-Atmospheres 111: Article Number: D02204
Mueller-Dombois DR, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, New York
Parsons RF, Uren NC (2007) The relationship between lime chlorosis, trace elements and Mundella Yellows. Austral Plant Path 36:415–418
Rabenhorst MC (1988) Determination of organic and carbonate carbon in calcareous soils using dry combustion. Soil Sci Soc Am J 52:965–969
Raison RJ (1979) Modification of the soil environment by vegetation fires, with particular reference to N transformations: a review. Plant Soil 51:73–108
Raison RJ, Khanna PK, Woods PV (1985) Mechanisms of element transfer to the atmosphere during vegetation fires. Can J For Res 15:132–140
Rayment GE, Higginson FR (1992) Australian laboratory handbook of soil and water chemical methods. Inkata, Melbourne, pp 110–114
Reid R (2006) Diameter-basal area ratio as a practical stand density measure for pruned plantations. For Ecol Manage 233:375–382
Ryan MG, Bond BJ, Law BE, Hubbard RM, Woodruff D, Cienciala E, Kucera K (2000) Transpiration and whole tree conductance in ponderosa pine trees of different heights. Oecologia 124:553–560
Sala A, Carey EV, Keane RE, Callaway RM (2001) Water use by whitebark pine and subalpine fir: potential consequences of fire exclusion in the northern Rocky Mountains. Tree Physiol 21:717–725
Shakesby RA, Chafer CJ, Doerr SH, Blake WH, Wallbrink P, Humphreys GS, Harrington BA (2003) Fire severity, water repellency characteristics and hydrogeomorphological changes following the Christmas 2001 Sydney forest fires. Aust Geo 34:147–175
Sillett SC, Van Pelt R, Koch GW, Ambrose AR, Carroll AL, Antoine ME, Mifsud BM (2010) Increasing wood production through old age in tall trees. For Ecol Manage 259:976–994
Stephens SL, Ruth LW (2005) Federal forest-fire policy in the United States. Ecol Apps 15:532–542
Stephens SL, Moghaddas JJ, Edminster C, Fieldler CE, Haase S, Harrington M, Keeley JE, Knapp EE, McIver JD, Metlen K, Skinner CN, Youngblood A (2009) Fire treatment effects on vegetation structure, fuels and potential fire severity in western U.S. forests. Ecol Apps 19:305–320
Swanborough PW, Lamont BB, February EC (2003) δ13C and water-use efficiency in Australian grasstrees and South African conifers over the last century. Oecologia 136:205–212
Tomkins IB, Kellas JD, Tolhurst KG, Oswin DA (1991) Effects of fire intensity on soil chemistry in a eucalypt forest. Aust J Soil Res 29:25–47
Wallin KF, Kolb TE, Skov KR, Wagner MR (2004) Seven-year results of the influence of thinning and burning restoration treatments on pre-settlement ponderosa pines at the Gus Pearson Natural Area. Rest Ecol 12:239–247
Ward D, Lamont BB, Burrows C (2001) Grasstrees reveal contrasting fire regimes in eucalypt forest before and after European settlement of southwestern Australia. For Ecol Manage 150:323–329
Warren CR, McGrath JF, Adams MA (2001) Water availability and carbon isotope discrimination in conifers. Oecologia 127:476–486
Werkman T, Davidson NJ, Close DC (2008) Is decline in high altitude eucalypt forests related to rainforest understorey development and altered soil bacteria following the long absence of fire? Austral Ecol 33:880–890
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase in western US forest wildfire activity. Science 313:940–943
Westfall PH, Tobias RD, Rom D, Wolfinger RD, Hichberg Y (1999) Multiple comparisons and multiple tests using the SAS system, SAS Institute Inc.
Withers J, Ashton DH (1977) Studies on the status of unburnt Eucalyptus woodland at Ocean Grove, Victoria. 1 The structure and regeneration. Aust J Bot 25:623–637
Youngblood A, Metlen KL, Coe K (2006) Changes in stand structure and composition after restoration treatments in low elevation dry forests of northeastern Oregon. For Ecol Manage 234:143–1
Acknowledgements
We gratefully acknowledge funding support from the Bushfire Cooperative Research Centre and The Forest Fire Management Group (Department of Environment and Conservation—Western Australia, Department of Environment and Heritage—South Australia, Department of Sustainability and Environment—Victoria, Forestry Tasmania and Forests New South Wales). Darren Harvey managed the vegetation clearing operations, site preparation and prescribed fires. The fire management authority reference group: Rick Sneeuwjagt, Drew Haswell, Tony Blanks, Dr Tim Wardlaw, Craige Brown, Vic Jurskis, and Dr Paul Barber provided advice. We thank Dr Robert Archibald for technical assistance and Daan Lock (Arbor Oxygen) for foliar sampling. Soil and leaf nutrient analyses were conducted by the CSBP Analytical Laboratory, Perth, Western Australia. Isotope ratio analyses were conducted by the Research School of Biological Sciences laboratory, the Australian National University, Canberra.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Tibor Kalapos.
Rights and permissions
About this article
Cite this article
Close, D.C., Davidson, N.J., Swanborough, P.W. et al. Does low-intensity surface fire increase water- and nutrient-availability to overstorey Eucalyptus gomphocephala?. Plant Soil 349, 203–214 (2011). https://doi.org/10.1007/s11104-011-0862-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-011-0862-3