
Abstract
Plantation forestry continues to intensify and grow in area, with a concomitant increase in fertilizer demand. Virtually no fertilizer is used on non-plantation forest systems. The scale of fertilizer use per ha per year in a small proportion of plantation systems is now similar to some agricultural production systems, but the total area of plantation forestry remains only a few percent of that used for agriculture. Hence, in a global context, forestry is a minor user of fertilizers. In relation to the knowledge base for fertilizer management, forestry and agriculture have similar practical questions that drive research, i.e. nutritional diagnosis and the development of fertilizer prescriptions that optimize production, environmental and economic goals. Much of this research is soil-climate-species-management specific. During the past few decades, solution culture methods were developed that maintain stable internal nutrient concentrations, which were essential for improving our understanding of nutrient-growth relationships. The development of plant production models that include the mechanistic simulation of nutrient supply and uptake are at an early stage of development. Plantation forestry and agriculture lack a mechanistic basis for evaluating base cation availability that accounts for Al-pH-root interactions. Further developments in this field could assist in rationalizing the use of lime. There is a lack of resources available in plantation forestry, and probably also in agriculture in some countries, to develop and refine calibrations of traditional types of soil and foliar analyses. Further testing of soil solution approaches is warranted. Further research on resource use efficiency, wood quality, rhizosphere relations, and mixed-species systems in relation to fertilization is also warranted.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Background
Fertilizer use in forestry is almost entirely restricted to plantations, of which there were 140 Mha world-wide in 2005 when this land use was increasing by 2.8 Mha annually (FAO 2006). The plantation area is likely to continue to expand, and almost all plantations receive fertilizer at some stage of their development. Hence, an increasing area of plantations leads to an increase in the demand for fertilizers in forestry. Another factor driving increased fertilizer usage in forestry is the level of intensification of management, with the objective of increasing the average rate of wood production per unit of land. Intensification reduces the duration of the crop cycle and increases the frequency of harvesting, and, because younger trees have higher concentrations of nutrients, it increases the average rate of nutrient export.
Until about three decades ago forest plantations had a rotation cycle of at least two decades, which was many times longer than most agricultural crops, but increasing demand globally has intensified management and led to rotation lengths of 4- to 7-years in some countries for growing several Eucalyptus and Acacia species. Demand for wood for traditional uses (paper, sawn timber, veneers, charcoal and firewood) drove this intensification, and short-rotation tree crops are emerging as a possible option for producing bioenergy (Gopalakrishnan et al. 2009). Most perennial horticultural crops have much longer rotations than these short-rotation forest plantations. Therefore, in relation to annual fertilizer application requirements, many forest plantations can be considered broadly similar to annual or perennial agricultural crops. However, Asia and South America, where the shortest-rotation forest plantations are grown, together account for only 24% of industrial roundwood production globally (FAO 2005). Most roundwood is still produced in much longer rotations in Europe, North America and Oceania from natural forests and plantations.
Forest nutrition research can be considered to have begun with the first detailed publications by Ebermayer in 1876 and 1882 in Germany, when it was noted that the periodic removal of the forest litter layer for use as animal bedding led to unhealthy forests, and that this effect was nutritionally mediated (cited by Attiwill and Leeper 1987; Rennie 1955 and Tamm 1995). Since Ebermayer, forest nutrition research has bolstered general knowledge of soil-plant nutrient relations in synergy with developments in agriculture and other domains of plant nutrition. Agricultural research with artificial fertilizers began around 1842 when Lawes and Gilbert took out a patent on the production of superphosphate, and in the same year Lawes published the effects on cabbage growth of applying different forms of inorganic nitrogen fertilizers (Johnston 1994). Only a few years later in 1847, inorganic fertilizer experiments on forest soils commenced in France (Baule and Fricker 1970; cited by Pritchett 1979). However, forest fertilization commenced operationally for many plantation types only in the 1950s, after the development of reasonably sound scientific principles (Ballard 1984). The use of fertilizer in forestry increased during the 1960s and 1970s as the economic value of fertilization became apparent; fertilizing to maximize tree growth rate became a priority (Cromer et al. 1977; Schönau and Herbert 1989). As the need developed to refine the management of forest fertilization, which was mainly in a plantation context, forestry began to contribute generally to the science of fertilization.
The objective of this review is to provide an overview of fertilizer usage in forestry internationally and the history of forest nutrition research, and describe developments in specific aspects of the knowledge base of forest fertilization that have implications for other cropping systems. I conclude by identifying major knowledge gaps and likely future trends in the research and practice of forest fertilization.
Goals of fertilization
In agriculture, fertilizers can be used to increase vegetative growth and total biomass production, e.g. for root-crops and pastures, but most fertilizers are used to increase reproductive growth for food production, e.g. cereals, oilseeds, and horticulture. In addition to maximizing yields, fertilizers are managed in many instances to optimize product quality, which is achieved by synchronizing the rates and timing of fertilization with the stage of growth. In contrast, the main goal of fertilization in forestry is to increase biomass production for wood volume and weight (which is the main context of other sections of this review) whilst avoiding serious deteriorations in wood quality. On a small scale, fertilizers are also used in forestry to manage seed production (Williams et al. 2003).
Where it has been of interest, wood quality in relation to fertilization has mainly been assessed for pulp and paper purposes. The principle qualities of interest are wood density, fibre diameter and length, microfibril angle, cell wall thickness, and contents of lignin, cellulose and extractives. The high cost of assessing these attributes has hindered progress, but the use of NIR analyses has recently reduced the cost and increased research in this area. In relation to eucalypts, fertilizer generally either increases or has no effect on wood quality, but it increases the nutrient content of wood and hence nutrient export from sites (Raymond 1998). In conifers, fertilization that increases growth rates also tends to decrease wood density, but overall there is usually an increase in the total value of wood for sawn timber or pulpwood (Antony et al. 2009; Cao et al. 2008; Downes et al. 2002; Nyakuengama et al. 2002; Zobel 1992). Overuse of fertilizers in forestry can result in trees with multiple or contorted stems that greatly reduces the value of the tree for either pulpwood or sawn timber purpose (Turnbull et al. 1994).
Fertilizer use in forestry
Forest plantations are widespread around the globe, with large areas in suitable parts of Asia, the Americas, and Europe, and much smaller areas in Africa and Oceania (FAO 2006). World-wide, forest plantations grown for wood products occupy about 3% of the total land area used for food and fibre production. The same percentage applies to Australia, where the low percentage of land used for plantations compared to agriculture, combined with low average annual rates of fertilizer usage per ha, results in plantations accounting for only 0.12% of N and 0.24% of P fertilizer usage (May et al. 2009b).
While the proportion of total fertilizer usage in plantations, and the average annual rates of application are low, per application the amounts and types of fertilizers are broadly similar to those in agriculture (Table 1). There is far less frequent use of fertilizers in forest plantations, where applications during the first year are common but later applications range from annually to never. The forms of fertilizer used in forestry mirror those used in agriculture, but some forestry-specific blends are available.
Like other cropping systems, the emphasis of most forest fertilization research has been on diagnosis of nutrient limitations in specific contexts (i.e. combinations of soils, climate, species and management) using soil and plant analyses, and the refinement of fertilizer rate, timing, form and placement options (Fox et al. 2007; Gonçalves and Barros 1999; Smethurst et al. 2004a). Fertilizer placement in a plantation considers uptake efficiency and the relative value of placement options. Planting densities are commonly 1,000–2,000 trees per ha, which means plants are several meters apart. Young tree seedlings have only small root systems. Therefore, applications around planting time are usually localized in concentrated spots or bands 10–50 cm from the planting position on or below the soil surface, or over small areas close to the plant (Smethurst and Wang 1998). As the root system develops, the zone of application usually increases in area until totally broadcasting the fertilizer becomes efficient (Attiwill and Adams 1996; Smethurst et al. 2004a), which in most countries is the cheapest and preferred way to apply fertilizer. Modern navigational methods aid the selection and recording of ground or flight paths for fertilizer application, which in-turn improves the uniformity of application, and improves the ability of pilots to avoid buffers and other areas that are not to be fertilized (e.g. McBroom et al. 2008).
Examples of forest fertilization
Slow-grown conifers
Some plantation conifers in Europe are grown in rotations of several decades (e.g. 92 years in central Sweden) and fertilized at planting with multi-nutrient mixes (Evers 1991). Thereafter N fertilizers are applied at various intensities up to 150 kg N ha−1 every 5 years, but more commonly less frequently or never (Eriksson et al. 2007). When these plantations are fertilized, it is commonly by broadcasting calcium ammonium nitrate or NPK mixtures.
Medium-growth-rate conifers
Coniferous plantations in Australia and the south-eastern USA grown in 15- to 30-year rotations are commonly fertilized at establishment with NPK mixtures applied close to the planting position, and application rates are 10–50 kg ha−1 each of N and P (Fox et al. 2007; May et al. 2009b). Established stands receive nutrients at high rates of 208–324 kg N and 50–112 kg P ha−1 broadcast every 5–15 years.
High-growth-rate eucalypts
Some eucalypt plantations in Brazil are grown on 6-year rotations that receive 2–3 applications of 20 kg N and 53 kg P ha−1 (mainly as monoammonium phosphate and diammonium phosphate) during the first 2 years, plus K and B (Barros et al. 2004; Gonçalves et al. 2008). Eucalypt rotations of about twice the length are used in Australia, with generally higher N and lower P rates than those in Brazil.
Liming
The use of lime (including dolomite) has increased in forestry during the past few decades in response to a perception that soil acidification and concomitant nutrient deficiencies threaten forest ecosystems. Liming of forests occurs predominantly in Central and Northern Europe (Kreutzer 1995; Meiwes et al. 2002). In Europe, soil acidification has accompanied the atmospheric deposition of nitrogen and sulphur generated from coal-fired power stations and from automobiles, i.e. the ‘acid rain’ effect. The realization of its potential consequences for forestry developed in the 1970s and 1980s, with the expectation that as a predisposing stress it would contribute to an increased incidence of toxicities and nutrient imbalances that would slow forest growth or in some cases lead to mortality (Hüttermann 1985; Matzner and Ulrich 1985). The accompanying critical load concept (Sverdrup and de Vries 1994) influenced many forest managers in Europe to commence or increase the use of lime (including dolomite). However, data on forest growth later indicated faster rather than slower growth of forests (Binkley and Högberg 1997), suggesting that the beneficial N- and S-fertilizer effects of atmospheric depositions outweighed any negative effects of acidification for several decades at least.
Liming can also have negative consequences associated with humus decay, nitrate concentrations in seepage water, mobilization of heavy metals, shallower root systems, boron deficiency and pathogens (Kreutzer 1995). Further, liming has had little effect on acidity in soil per se, because it is applied to established forests with a litter layer, and most of the chemical changes occur only in the litter layer. Changes in litter chemistry can result in higher rates of nitrification and increased rates of nitrate leaching at a catchment scale, and other adverse outcomes (Löfgren et al. 2009). Far more important to countering the effects of acid deposition than liming are the trends in electricity production during the past few decades that have reduced S deposition by 90% and N deposition by 50% in southern Sweden, for example (Löfgren et al. 2009).
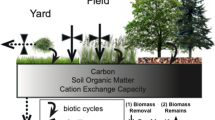
Liming is also used routinely by some plantation forestry companies in Brazil to counter soil acidity, but its main purpose is to correct Ca and Mg deficiencies. This practice is not universal amongst plantation companies in that country, and experimental results indicate that liming to correct acidity is not warranted when planting with Eucalyptus or Pinus species, which are tolerant of acid soils (Gonçalves et al. 1997). What appears to be an unnecessary use of lime in Europe, and in some cases in Brazil, underscores the risks associated with acting on expected soil chemistry and plant growth effects without confirmation of the latter under normal field conditions. While some progress has been made in our understanding of the effects of forest liming, we particularly need to separate the effects of liming to alleviate the toxic effects of H and Al from those on nutrient availability at root membrane to stand scales. There is a larger knowledge base for some agricultural species, but in that domain too we still lack an adequate quantitative, mechanistic, understanding of the interactive effects of liming across a range of spatial and temporal scales. A schematic of the main pools and processes involved is presented in Fig. 1. A number of these aspects have been studied in detail for some soil-plant systems, but quantitative links have not been established for any plant-soil system. Until such understanding and quantification is available, we will have to rely on calibrated, empirical relations between fertilizer or lime applications and ecosystem responses.

Schematic diagram of the main soil-plant processes involved in Al-H-base cation availability responses to liming and fertilization
Fertilizer use in agriculture
There is a very wide range of fertilizer practices used in agriculture due to the variety of natural and socio-economic conditions, crop requirements, nutrient forms and fertilizer availability. Examples are provided in Table 1 for some Australian agricultural systems, which show that relatively high annual rates are used in horticulture and sugarcane, and lower rates in cereals, oilseeds and dairy systems. These latter systems have rates about twice those in intensively managed forest plantations. Annual rates of fertilizer usage in Australian agriculture are within the large range reported for cropping and pastures in the UK (Chalmers 2001), vegetable crops in China (Chen et al. 2004), and also broad-acre and horticultural crops in various regions of the world (Crews and Peoples 2005).
Understanding forest nutrition and growth
Nutrient cycling
Although fertilizer management in forest plantations is similar in principle and practice to many agricultural situations, differences in some aspects of nutrient cycling need to be considered. Prior to 1960, plant nutrition as a discipline had no particular emphasis on forests, but in the period 1960–1990 forest nutrition developed as a discipline of research by focusing on nutrient budgets and nutrient cycling in a variety of natural and managed forest systems (Smethurst 2004). Some of these systems had been fertilized, but not all. Comerford (2002) lists 16 key texts on forest soils published between 1983 and 2000 that document an increasing emphasis on nutritional management. To these can be added Bowen and Nambiar (1984), Nambiar and Brown (1997), and Gonçalves (2004) that specifically develop the topic in a plantation context.
As Comerford (2002) highlighted, there are several aspects of forest soils that are distinct from agricultural systems: time between harvests, nutrient cycling, erosion, topography, fertilization practices, irrigation, soil temperature, utilized soil depth, stoniness, remoteness from markets, and surface soil organic horizons. All of these differences impinge to some degree on fertilizer management, some of which are further explored here.
In trees, the annual requirement for nutrients for new growth is partially met by drawing on internal pools in older tissues rather than meeting nutrient demand entirely by uptake from soil. In this context, internal cycling includes the withdrawal of nutrients from aging or dying tissues and retranslocation of those nutrients to younger tissues. The importance of this process had been recognized by the 1970s (Wells and Jorgensen 1975), and it has been quantified in a number of forest systems since. In a 13-year-old, N-deficient Pinus radiata plantation, N fertilization led to a 45% increase in wood production and 350% increase in N retranslocation, showing increased reliance on retranslocation as growth rate increased (Fife and Nambiar 1997). Internal cycling of nutrients in three Eucalyptus species, Acacia mearnsii and Pinus radiata was studied between 12 and 22 months of age, during which 31–60% of N present in young, green foliage, 54–63% of P and 18–38% of K was retranslocated (Fife et al. 2008). Hence, such dependence on retranslocation probably exists among all or many forest plantation species. In contrast, annual agricultural crops depend almost entirely on uptake, during which sub-annual transfers of carbohydrates and nutrients are important for grain, fruit or tuber production.
Cycling of nutrients external to the plant is also prevalent in forests that have commenced above- or below-ground litter production, and it can account for a significant component of annual nutrient availability to tree roots. For example, tropical plantations of various species accumulate in litter 80–660 kg N and 4–20 kg P ha−1, which is more than in many natural tropical forests (O’Connell and Sankaran 1997) and generally more than the average annual amounts of N and P accumulated in trees of tropical forest plantations (Gonçalves et al. 1997). Hence, retention of this material and harvesting residues between rotations is essential to reduce or alleviate the need for fertilizers during the early phase of the next crop (Nambiar 2008). Conversely, slow decomposition of litter in some systems, particularly in cold climates, immobilizes large pools of N and other nutrients (Tamm 1995). This build-up of nutrients in litter layers limits nutrient losses via leaching from these ecosystems, but it can also contribute to N deficiency in trees.
Links between nutrient cycling and catchment-scale outcomes were made early in forestry because some aspects of forestry practices were controversial, i.e. clearfelling and complete vegetation control using herbicides could lead to high nitrate concentrations in streams and ground water (Vitousek and Melillo 1979). Later studies have shown that N and P fertilizer in forestry leads to generally small and transient increases in N and P concentrations in stream water (Binkley et al. 1999). More recently, practices in agriculture have come under scrutiny in a nutrient context for potential adverse catchment scale and greenhouse gas outcomes (McDowell 2008). Agriculture generally, via fertilizer use, has caused a much larger perturbation of global nutrient cycles than forestry (May et al. 2009b). For example, by about 1990, global N fertilizer production, which is mainly used to produce food, and other anthropogenic influences had doubled the rate of N transfer from atmospheric to terrestrial pools compared to that which would otherwise have occurred due to natural processes (Vitousek et al. 1997). This proportion has probably increased substantially during the past decade.
Nutrient flux density and uptake kinetics
Developing a mechanistic understanding of nutrient uptake processes has led to the realization that nutrient uptake at the cell membrane level of roots and mycorrhizal hyphae was an active process mediated by enzymes. Together with nutrient supply phenomena in soils, supply and uptake could be mathematically modeled (Barber 1995; Tinker and Nye 2000). Between 1970 and 1991, the need to experimentally control plant growth rates and nutrient concentrations led to the development of hydroponic culture techniques that supplied all nutrients at non-limiting concentrations, except for one limiting nutrient (Asher and Blamey 1987; Asher and Cowie 1970; Ingestad 1971). Supply of the limiting nutrient (usually N) conformed to a schedule that directly led to control of the growth rate of the plant. Different exponential growth rates resulted in different but stable internal nutrient concentrations, which greatly assisted in the testing of various hypotheses of nutrient-growth relations in plants (Ericsson et al. 1995; Ingestad 1982).
However, Ingestad came to the conclusion that concentrations of nutrients in hydroponic and soil solutions and uptake kinetics of plant roots were unimportant for controlling plant growth rates (Ingestad 1982). Instead, the rate of nutrient replenishment was critical, which he and his coauthors termed the nutrient flux density approach. This method was also applied to several field experiments in forests around the world (Albaugh et al. 2007; Linder 1995) and was adopted by scientists working on nutrient-growth relations of other forestry, agricultural, and aquatic species (e.g. Groot et al. 2002; Hawkins et al. 2005; Macduff et al. 1993; Pintro et al. 2004; Raven 2001), because of the high level of control it offers over plant growth rates and internal nutrient concentrations.
The nutrient flux density approach presented a dilemma for some scientists working in the more common paradigm where concentrations in growth solutions and uptake kinetics were important (Macduff et al. 1993; Raven 2001). Sands and Smethurst (1995) subsequently demonstrated that these two approaches were not necessarily inconsistent by using uptake kinetic principles to model nutrient uptake and plant growth reported for one of Ingestad’s experiments. A solution concentration of 50 μM inorganic N was required (via periodic or continuous replenishment) to produce a relative growth rate of 0.25. Building on the earlier work of Tinker and Nye (2000) and Barber (1995), nutrient supply and uptake theory has since been used to simulate nutrient uptake and growth of a eucalyptus plantation over several years with simultaneous potential limitations of N, P, light, water or temperature (Smethurst et al. 2004b).
CEC and base cations
Forest soils are generally more acidic and organic than soils used for agriculture. Experience with forest soils has questioned traditional views developed primarily in an agricultural context that use cation exchange capacity, base cation saturation and exchangeable cation concentrations as indicators of base cation availability (Ross et al. 2008). In such forest soils, the source of charge is mainly organic matter, and cation retention cannot be explained by simple exchange phenomena. Ross et al. (2008) argue four salient points that apply to acid soils across agriculture and forestry: (1) new measures of exchangeable Al and H are needed, (2) base saturation should be abandoned as a measure of base cation availability, (3) the paradigm that higher pH accompanies higher CEC does not hold, and (4) CEC should not be used to indirectly infer base cation availability. These observations will become more pertinent as agricultural and forestry soils acidify, which is a long-term trend globally, and as society is forced to use more acid soils to grow food and fibre.
In many parts of the world, salinity or sodicity adversely affects crop growth, and the problem is expanding. During recent decades, Na has been recognized as a functional nutrient for many agricultural plant species, and to some extent it can substitute for the functions of Ca, Mg and K (Subbarao et al. 2003). One implication is that increasing Na availability and uptake can reduce the need for these other base cations, and lower critical foliar concentrations can be used as a guide to fertilizer needs. However, few studies have demonstrated Na substitution of other bases under field conditions. A recent demonstration of this effect in a plantation forestry context comes from Brazil, where K depletion after several decades of eucalypt cropping led to substantial K deficiency (Almeida et al. 2009). Trees responded substantially to K fertilizer applications, and about 40% of that response was also achieved by Na fertilization alone.
Productivity modeling that includes nutrients
Agricultural crop productivity modeling that accounts for species-specific responses to light, water, temperature, and nitrogen was already well developed more than a decade ago (Hanks and Ritchie 1991; Keating et al. 2003), and has since expanded in sophistication and application. A similar model has been developed for forest plantations (Landsberg and Waring 1997), but it considers nutrients by using only a generic fertility factor, and it cannot account for fertilizer applications. A more mechanistic model has been developed that considers some silvicultural operations used in plantation forestry, including N fertilization (Battaglia et al. 2004), which was used to simulate N and P uptake and estimate growth limitation due to light, water, N and P (Smethurst et al. 2004b). Other forest productivity models have been developed that account for one or more nutrients (usually N), but none combine a high level of silvicultural flexibility with detailed nutrient dynamics and uptake by trees and weeds (Smethurst 2007). These process-based plantation forestry models also do not yet have the sophistication to account for all the options available for managing fertilizers, e.g. fertilizer forms and placement, but such developments are possible. Such detailed fertilizer management options can instead be compared using more empirical models or decision support systems based on experience and financial information (Fox et al. 2007; May et al. 2009a).
Nutrient use efficiency
Forest researchers have had an interest in nutrient use efficiency (NUE) for several decades. Based on litterfall as a surrogate for growth and net primary productivity (NPP), and litter N content as a surrogate for N supply, it was asserted that the more nutrient that was used the less was its NUE, i.e. biomass produced per unit of resource supply decreased with an increase in supply (Vitousek 1982). With actual measures of above-ground NPP (ANPP), Binkley et al. (2004) sought evidence that this was the case for the resources of light, water and nitrogen, and found that the few data available supported the opposite hypothesis, i.e. resource use efficiency increased with increasing resource capture. For example, across 14 Eucalyptus stands in Brazil, Stape et al. (2004) found that ANPP per unit of N uptake, increased about 30% with a 200% increase in N uptake, while litterfall per unit of N uptake and litterfall proportion of ANPP both decreased by about 50%, with a 300% increase in ANPP.
A key concept in agriculture is that resource use efficiency is based on product yield per unit of resource supply (e.g. Passioura 2004 for water). But for nutrients, the contribution of both fertilizer and soil sources needs to be considered, and resource capture needs to be separated from resource use within the plant. There are few data in agriculture or forestry that allow a full analysis of nutrient use efficiency, but instead product yield can be expressed per unit of external resource input (e.g. irrigation or fertilizer). This latter index has also been referred to as the partial factor productivity (PFP) of an applied nutrient (Dobermann 2007). Nutrient use efficiency can also be expressed as biomass or product yield per unit of nutrient taken up regardless of the nutrient source, which approximates to the inverse of the average concentration of a nutrient in a plant. It is important that such concepts are more fully understood by including below-ground productivity and measures of resource supply and resource capture, and that the implications for fertilizer practices are clarified.
Various fuel sources are used or considered for biofuel production, but these can have a very wide range of PFP values (Table 2). These data indicate that corn, cereals and pasture have very low PFP values (0.6–144) compared to some eucalypt plantations that had a low requirement for N fertilizer (2–14 k), but similar to pine plantations that had a high N fertilizer requirement. Biofuel production requiring little or no fertilizer input would have a PFP value approaching infinity, e.g. in riparian plantings of trees and grasses that utilize nutrients in wastewater or polluted groundwater (Gopalakrishnan et al. 2009) or in parts of Europe and North America with high atmospheric deposition. As biofuel technologies develop to better cope with woody materials, it is expected that more of this feedstock will be used and thereby increase the N-use efficiency of biofuel production (Galloway et al. 2008), but it will be important to consider the N fertilizer requirement of the particular plantation system under consideration.
Production systems with less fertilizer
There is a concern that the availability of inorganic N and P fertilizers will diminish as the surplus of supply over demand decreases (FAO 2008; Huang 2009) and as mineral reserves of P-containing rocks are depleted (Cordell et al. 2009). However, others argue that world reserves and resources for N and P appear adequate for at least the next 2 years (FAO 2008) and the foreseeable future (Fixen 2009). Under both scenarios, the price of fertilizers could increase relative to the cost of other farm inputs. Lack of availability, or economic pressures could therefore reduce the per ha use of these fertilizers and necessitate food and fibre production using lower nutrient input systems. This trend could in-turn expanded interest in the use of rhizosphere and mixed-species technologies that potentially add N and better utilise existing sources of soil N and P otherwise unavailable to crop plants. These technologies include enhanced P uptake by roots and mycorrhizae via organic acid and phosphatase production (Jones 1998; Richardson et al. 2009), symbiotic (Forrester et al. 2006) and non-symbiotic biological N-fixation, and improved plant root nutrient uptake kinetics (Bassirirad 2000; Raghothama 1999). There is scope for (1) using existing genotypes that can up-regulate these mechanisms, (2) selecting and breeding for new genotypes of plants and microbes, and (3) genetic engineering to induce or enhance these processes in important crop species (Schachtman and Shin 2007; Richardson et al. 2009). There will probably be a greater need for these developments in agriculture than in forestry, so important opportunities might develop for plantation forestry by being aware of such trends in agriculture. However, studies of forest and other non-agricultural crop species have contributed significantly to our current understanding of these processes, because of their importance in low-fertility, non-agricultural systems.
Mycorrhizae, fungal symbionts that infect the root system, act as a very fine extension of the root system with a high surface area to biomass ratio that enhances under some conditions nutrient and water uptake and the ability of a plant to cope with biotic and abiotic stresses. In return for these potential benefits, the plant supplies carbon to the fungus for growth and metabolism. Many of these functions are genetically mediated (Graham and Miller 2005). In relation to nutrient uptake, plants benefit most from the mycorrhizal symbiosis in terms of biomass growth and survival under conditions of low availability of poorly-mobile nutrients, e.g. P in highly P-limited natural and man-made ecosystems (Chen et al. 2008). In contrast, under high nutrient availability, infection and growth of mycorrhizae can be negligible, or the cost in terms of carbon can reduce plant growth where nutrient uptake is not enhanced by mycorrhizae. These principles have been amply demonstrated in controlled environment and field conditions using a variety of plant species (e.g. perennial versus annual, woody versus non-woody, domesticated versus non-domesticated species) (Graham and Miller 2005). However, field studies can be difficult due in-part to a limited ability to control and measure the level of infection by inoculated and endemic species of fungi. Organic acid and phosphatase enzyme exudates increase the availability of inorganic and organic soil P, respectively, by converting solid-phase P to liquid-phase phosphate, which is the main form of P taken up by roots and mycorrhizal hyphae of crops (Richardson et al. 2009).
Nitrogen-fixing plants have been of interest in plantation forestry for many years as primary crop species or as nurse/companion crops to other main species in plantation forests or agro-forestry systems (Forrester et al. 2006). Increased N availability benefits in such systems have been well-demonstrated (Binkley and Giardina 1997), but major deterrents to their adoption on a large scale have been management complexity and economic viability. An overall financial benefit is not obvious unless fertilizer costs are very high or the wood value gained is very high (Turvey and Smethurst 1983). These financial benefits will probably become more obvious if fertilizer and fibre shortages develop. If or when that situation eventuates, plantation forestry and agriculture will need to seriously consider mixed-species systems, for which there are many generic and system-specific research and management questions remaining to be answered.
Ion flux and membrane transporter technologies
In recent years, the use of ion flux and membrane transporter technologies have started to expand our understanding of the nutritional physiology of trees. Wood formation has a particular dependence on potassium (K), the supply of which to cambial cells is regulated by K+ channels (Langer et al. 2002). In Populus tremula, one transporter was continuously present at a low level, suggesting a house-keeper function, but the levels of two others followed the annual variation in plant growth (Langer et al. 2002). Functioning of theses transporters (and hence uptake of K into cambial and expanding xylem cells) depends on the necessary H+-gradient being generated by a H+-ATPase in the plasma membrane, and this response can be initiated within a few hours of the addition of auxin to dormant twigs (Arend et al. 2002). Similar transporter and ion flux studies in tree roots also indicate a key role for H+-pumping to maintain the electrochemical gradient that drives the fluxes of K and Na (Knowles 2007; Sun et al. 2009). However, when measuring fluxes of these cations, care needs to be exercised in the use of the non-invasive, ion-selective flux microelectrode technology, because one needs to correctly account for the non-ideal, ion-selective behavior of the resins used to make the microelectrodes (Knowles and Shabala 2004).
Using Populus, Eucalyptus, and agricultural species (Escalante-Pérez et al. 2009; Knowles 2007; Sun et al. 2009), these technologies are now elucidating the genetic and physiological basis to salt sensitivity and tolerance. Regulation of the proton gradient across the plasma membrane reduces Na+ influx via non-specific cation channels and simultaneously reduces K+ efflux through depolarization-activated channels. The addition of Ca2+ markedly enhances these processes and thereby assists in maintaining K+/Na+ homeostasis.
Ion-flux technologies are also available for nitrogen, but not yet for phosphorus. Simultaneous measurements of ammonium, nitrate and proton fluxes around roots of Eucalyptus nitens revealed a preference for ammonium, spatial and temporal variations in fluxes in the 20–60 mm region from the root tip, no affect of proximity to root hairs or root laterals, and Michaelis-Menten-style uptake kinetics (Garnett et al. 2001, 2003). Ammonium preference is a more common observation in forest ecosystems than in agricultural systems (Kronzucker et al. 1997; Min et al. 2000). Such results encourage the continued use of ammonium-based fertilizers in forestry. Increased development and use of ion-flux technologies will be needed as we seek to further expand our understanding of plant nutritional physiology and improve fertilizer management, but we need to be cognizant of some current limitations. For example, these methods are mainly suitable for young plants in controlled environments that experience very different growing conditions to field-grown plants, and the lower limits of concentration detection are not as low as the concentrations at which uptake occurs in many forestry and agricultural cropping systems.
Indicators of plantation response to fertilization
There is a well-established method of calibrating soil and plant indicators of potential growth (or yield) response to fertilization (McLaughlin et al. 1999; Smith and Loneragan 1997). This method requires that many fertilizer experiments be established in time and space to capture climatic and landscape variability. Yield of the unfertilized treatment is expressed relative to maximum growth with fertilizer, and these data plotted as a function of the indicator, e.g. a soil or leaf analysis. The value of the indicator at the point where relative yield decreases significantly from below 1.0 (generally taken as 0.90 or 0.95 relative yield) is referred to as the critical value for that indicator. Instead of relative yield, other yield response criteria can also be used, e.g. percent increase in growth due to fertilization.
Fertilizer needs in agriculture commonly take into account soil and plant analyses that are well calibrated. As plantation forestry intensifies and the relative cost of fertilizer increases there will be a need to further develop and apply suitable soil and plant analyses. Appropriate calibrations and their use will not only guide fertilizer use in low- to medium-input systems, but they can also be useful for avoiding the over-use of fertilizers and concomitant environmental problems such as those that occur already in parts of Europe, USA, China and Mexico (Vitousek et al. 2009). Fertilizer practices in forestry should be developed while avoiding over-use.
This protocol for calibrating potential indicators of nutrient deficiency is rarely fully applied in forestry, because resources have often not been available to cater for the long crop cycles and large plot and plant sizes. Some examples are available, and critical indicator values are also inferred, but less-reliably so, from growth and nutrient indicator surveys across operational plantations. Such inferences have also been developed from growth and nutrient relations within just one or a few fertilizer experiments. In the following paragraphs, examples of nutrient limitations in several major plantation regions of the world are described along with the criteria used to predict fertilizer responses.
Pinus plantations, south-eastern USA
A large concentration of forest plantations of Pinus elliottii and P. taeda (13 M ha) are grown in their native range in the south-eastern USA, and fertilization has been a key component of management intensification during the past five decades (Fox et al. 2007). Nutritionally, these plantations are severely limited by low P and N supply if unfertilized. P-fertilizer is needed at planting (Pritchett et al. 1961), and thereafter a combination of N and P is far better than either nutrient alone (Amateis et al. 2000). The concentration of P in an acid extract (Bray2-P) best discriminated between responsive and non-responsive sites at planting (Ballard and Pritchett 1975). Once established, leaf area index (LAI) is the main diagnostic. A fully stocked stand with stem cross-sectional areas at 1.3 m height greater than 22.9 m2 ha−1 should have an LAI of at least 3.5, unless there are other obvious problems that have altered LAI, e.g. fire, ice, insects etc. (Fox et al. 2007). It is possible to use these criteria, because water is generally less limiting than low N and P supply at this stage of the crop.
Pinus and eucalyptus plantations, Australia
Almost 2 M ha of Pinus and Eucalyptus plantations are grown in Australia. Where grown on ex-native forest sites that do not have a history of P-fertilization for agricultural production, applications of P fertilizer at planting are essential (Boomsma 1949). Various indices of soil, plant and litter P have been sought for these plantations (May et al. 2009b). Although a common basis for assessment is still lacking, some of these indexes have been calibrated for specific soil-climate-species-management contexts, e.g. CaCl2-extractable P for Eucalyptus globulus and E. nitens in temperate Australia (Mendham et al. 2002) and total P concentration in litter for Pinus radiata in south-east South Australia (May et al. 2009a).
As for Pinus plantations in the south-eastern USA, LAI has been recognized as a key determinant of potential growth response to N fertilization in Pinus radiata plantations in south-east South Australia (May et al. 2009a) and in Eucalyptus nitens plantations in Tasmania (Smethurst et al. 2003). Soil and litter N analyses have also been examined as indicators for these plantations; litter N concentrations in Pinus radiata were significantly correlated with response to N-plus-P fertilization (May et al. 2009a). The critical concentration of total N in surface soil was 6 mg g−1 for Eucalyptus nitens plantations in Tasmania (Smethurst et al. 2004a) and 2 mg g−1 for E. globulus plantations in Western Australia (White et al. 2009), which illustrates the contextualization required for many critical concentrations based on soil analyses.
Deficiencies of K in Pinus radiata occur under some circumstances in Australia, e.g. on ex-farmland where decades of K removals in agricultural products have not been replaced by fertilization (Smethurst et al. 2007). Although foliar analysis appears useful as a diagnostic tool in these circumstances, further refinement of the critical concentrations is warranted to account for soil type, management and climate (Smethurst et al. 2007).
Eucalyptus and Pinus plantations, Brazil
The area of eucalypt and pine plantations in Brazil has increased rapidly during the past two decades, and now totals 4 Mha for eucalypts and 2 Mha for pines. Applications of P and K were recognized early as necessary in many regions (Barros et al. 2004). Little or no N fertilization was needed for the first rotation. The incidence and severity of N deficiency on these sites is expected to increase with subsequent rotations, and relative growth of 0.74 to 0.98 has recently been documented in later rotations by age 2 years at 11 sites in São Paulo State (Pulito 2009). At three of these sites, trees had already reached or were close to harvest age of 7 years, and in each of these cases relative growth had increased from 0.74–0.83 at 2 years of age to 1.0 at around 7 years, indicating that the earlier response to N fertilizer had disappeared. The reason for this change was not investigated, and raises the hypothesis that N or another resource became limiting between 2 and 7 years. Specific for either Eucalyptus or Pinus, critical concentrations of organic matter had earlier been proposed as an indicator of the need for N fertilization (Barros et al. 2004), but this criterion was not supported by the Pulito (2009) data. Resin-P for P fertilization, and exchangeable K for K fertilization are still a current recommendation and critical concentrations of these indicators depend on clay content. (Barros et al. 2004).
Soil solutions as indicators of nutrient supply
In agriculture and forestry, the development of soil and foliar analyses as indicators of nutrient deficiency has largely been the responsibility of public organizations like universities and state or national departments of research and extension. However, during the past decade it has become difficult for these organizations to resource this activity, and in many cases it has been dropped as an important objective unless the user-pay principle is applied. Meanwhile, crop genotypes, climate, crop management, and soil conditions have changed. Such changes would be expected to in-turn change the critical concentrations of soil and foliar diagnostics. With food and fibre shortages increasing globally, and system inputs becoming more expensive, including fertilizers, there is a mismatch between the need to use resources more efficiently, and the knowledge base for making fertilizer management decisions. Either traditional systems for developing critical nutrient concentrations need to be re-built, or new technologies need to be developed that are less expensive. This dilemma developed in plantation forestry some years ago and led to consideration of nutrient concentrations in soil solutions as potential indictors that were more generic than traditional, soil-type-specific measures, which rely on strong acid, alkaline or salt extracts (Smethurst 2000).
The desire to interpret nutrient availability from nutrient concentrations in soil solution also motivated others to seek appropriate methods. For example, Barraclough (1989) demonstrated how critical soil solution concentrations for agricultural crops in the UK might be used to derive more accurate critical concentrations of more traditional soil indices by accounting for volumetric soil water content (θv). A soil of θv = 0.34 was estimated to have a critical Olsen P value of 7 μg g−1, in comparison to a drier soil (θv = 0.30) requiring a higher critical Olsen P value of 23 μg g−1. This motivation and operational simplicity were also behind development of a dilute calcium chloride extract (0.01 M CaCl2) as an indicator of nutrient availability in European agricultural soils; this solution mimics the ionic strength and pH of soil solutions (Houba et al. 2000). Such an extract was particularly useful for identifying P deficiency in pastures (Dear et al. 1992) and temperate eucalypt plantations (Mendham et al. 2002).
In a research context, soil solution technology has proven more useful than traditional measures of soil fertility where well-established calibrations are lacking (e.g. Smethurst et al. 2001, 2007). Can this technology be developed also as an operational method? The principle of using soil solutions for inferring nutrient deficiencies relies on three main steps. Firstly, the concentration of the inorganic nutrient needs to be measured in a paste extract. Secondly, this concentration may need adjusting for potential dilution effects that can occur during preparation of the paste. The adjusted concentration is an estimate of the concentration in bulk soil solution. Thirdly, the bulk soil solution concentration is interpreted on the basis of that which is required at root surfaces to maintain near-optimum growth.
There is a possibility that the soil solution method could be adapted to low-cost, portable equipment by using simple centrifugation or suction methods to extract the paste solution and by analyzing the solution using portable water analysis equipment (Osborne et al. 2001). The essential aspects of this method were demonstrated in a survey of forest plantations in four regions of Australia, where a large part of the variation in soil fertility is attributed to fertilizer history, in particular whether there had been a pasture phase accompanied by fertilizer inputs. Paste samples were prepared using de-ionized water, and solution was extracted using porous ceramic tips (Rhizon© solution samplers purchased from Eijkelkamp) to which a partial vacuum had been applied. Solutions were analyzed for NH4, NO3, PO4 and K on a portable spectrophotometer designed for water analysis (Spectroquant Nova 60 Photometer© purchased from Merck). Due to buffering by the solid phase, concentrations of NH4, PO4 and K in soil solution were assumed to be altered little by the addition of water during preparation of the paste, but NO3 concentrations were adjusted to account for the dilution effect.
Frequency distributions of concentrations of NH4, NO3, and PO4 indicated that higher values were more common on ex-pasture sites than on ex-forest sites (Fig. 2). In a separate study, soil solution NH4 and NO3 correctly discriminated between 6 responsive and 2 non-responsive E. nitens plantations when fertilized with N. This level of discrimination was better than that achieved by the commonly used KCl extract (Smethurst et al. 2004a). Soil solution P and its surrogate CaCl2-P were also good discriminators of P responsive sites (Mendham et al. 2002). Even if the soil solution approach is adopted, it will not eliminate the need for field experiments; instead, its potential to reduce the need for field experiments and to provide a more generic approach to soil fertility assessment warrants further testing.

Frequency distribution of surface soil NO3 (top), NH4 (middle) and PO4 (bottom) concentrations measured in Australian forest plantations grown on ex-pasture and ex-forest sites. Concentrations were measured using a low-cost, portable nutrient analysis system (Osborne et al. 2001)
Conclusions and knowledge gaps
This review places several aspects of forest fertilization developments in an agricultural context, and identifies knowledge gaps:
-
1.
Fertilizer use in intensive plantation forestry does not approach the usage seen in the most intensive pasture (dairy) or annual cropping systems on a per ha per year basis.
-
2.
However, intensive plantation forestry systems are similar to some biennial or perennial agricultural systems in the length of the crop cycle and the rates of fertilizer applied per application.
-
3.
Less intensive plantations systems have much longer rotations than in agriculture and much lower rates of fertilizer use, and plantation forestry overall occupies a small portion of the landscape compared with agriculture.
-
4.
Fertilization strategies in forestry consider internal and external nutrient cycling and deep rooting.
-
5.
As an option for improving nutrient use efficiency, both forestry and agriculture will need to further develop rhizosphere and mixed-species technologies.
-
6.
Forest scientists have contributed significantly to developing methods of plant culture that maintain stable internal nutrient concentrations. As a research tool, this development should continue to improve our understanding of nutrient-growth relationships.
-
7.
The development of plant production models that include the mechanistic simulation of nutrient supply, uptake and weed competition are at a similar early stage of development in both agriculture and forestry. Nitrogen modeling is more advanced than other nutrients.
-
8.
Plantation forestry lacks a mechanistic basis for evaluating base cation availability that accounts for Al-pH-root interactions, which is also the situation in agriculture. Further developments in this field could assist in rationalizing the use of lime.
-
9.
Traditional concepts of nutrient use efficiency in forestry have recently been challenged and need clarifying in the context of fertilizer management for food, fiber and biofuel production.
-
10.
There is an on-going need to develop calibrations of traditional types of soil and foliar analyses, but, in some countries, organizations that have developed these calibrations in the past are finding it difficult to resource further work.
-
11.
Testing soil solution approaches of assessing soil fertility are warranted, because they offer a more generic and affordable approach that might reduce the need for field experimentation.
References
Albaugh TJ, Allen HL, Dougherty PM, Johnsen KH (2004) Long term growth responses of loblolly pine to optimal nutrient and water resource availability. For Ecol Manage 192:3–19
Albaugh TJ, Allen HL, Fox TR (2007) Historical patterns of forest fertilization in the southeastern United States from 1969 to 2004. South J Appl For 31:129–137
Almeida JCR, Laclau J-P, Gonçalves JLM, Ranger J, Saint-André L (2009) A positive growth response to NaCl applications in Eucalyptus plantations established on K-deficient soils. For Ecol Manage (in press)
Amateis RL, Liu J, Ducey M, Allen HL (2000) Modeling response to midrotation nitrogen and phosphorus fertilization in loblolly pine plantations. South J Appl For 24:207–212
Antony F, Jordan L, Daniels RF, Schimleck LR, Clark A, Hall DB (2009) Effect of midrotation fertilization on growth and specific gravity of loblolly pine. Can J For Res 39:928–935
Arend M, Weisenseel MH, Brummer M, Osswald W, Fromm JH (2002) Seasonal changes of plasma membrane H+-ATPase and endogenous ion current during cambial growth in poplar plants. Plant Physiol 129:1651–1663
Asher CJ, Cowie AM (1970) Programmed nutrient addition—a simple method for controlling the nutrient status of plants. In: Proceedings of Australian plant nutrition conference, Sept. 1970, Mt Gambier, South Australia, pp 28–32
Asher CJ, Blamey FP (1987) Experimental control of plant nutrient status using programmed nutrient addition. J Plant Nutr 10:1371–1380
Attiwill PM, Leeper GW (1987) Forest soils and nutrient cycles. Melbourne University Press, Melbourne
Attiwill PM, Adams MA (eds) (1996) Nutrition of eucalypts. CSIRO, Collingwood
Ballard R (1984) Fertilization of plantations. In: Bowen GD, Nambiar EKS (eds) Nutrition of plantation forests. Academic, London, pp 327–360
Ballard R, Pritchett WL (1975) Evaluation of soil testing methods for predicting growth and response of Pinus elliottii to phosphorus fertilization. Soil Sci Soc Am J 39:132–136
Barber SA (1995) Soil nutrient bioavailability. Wiley, New York
Barraclough PB (1989) Root growth, macro-nutrient uptake dynamics and soil fertility requirements of a high-yielding winter oilseed rape crop. Plant Soil 119:59–70
Barros NF, Novais RF (1996) Eucalypt nutrition and fertilizer regimes in Brazil. In: Attiwill PM, Adams MA (eds) Nutrition of eucalypts. CSIRO, Collingwood, pp 335–355
Barros NF, Neves JCL, Novais RF (2004) Mineral fertilizer recommendations for eucalypt plantations. In: Gonçalves JLM (ed) Forest nutrition and fertilization. IPEF, Piracicaba, pp 269–284
Bassirirad H (2000) Kinetics of nutrient uptake by roots: responses to global change. New Phytol 147:155–169
Battaglia M, Sands P, White D, Mummery D (2004) CABALA: a linked carbon, water and nitrogen model for forest growth for silvicultural decision support. For Ecol Manage 193:251–282
Baule H, Fricker C (1970) The fertilizer treatments of forest trees. BLV Verlagsgesellschaft mbH, Munich
Binkley D, Giardina C (1997) Nitrogen fixation in tropical forest plantations. In: Nambiar EKS, Brown AG (eds) Management of soil, nutrients and water in tropical plantation forests. ACIAR Monograph No. 43, pp 297–337
Binkley D, Högberg P (1997) Does atmospheric deposition of nitrogen threaten Swedish forests? For Ecol Manage 92:119–152
Binkley D, Burnham H, Allen HL (1999) Water quality impacts of forest fertilization with nitrogen and phosphorus. For Ecol Manage 121:191–213
Binkley D, Stape JL, Ryan MG (2004) Thinking about efficiency of resource use in forests. For Ecol Manage 193:5–16
Boomsma CD (1949) Phosphate for top dressing as a normal plantation operation. Aust For 13:108–112
Bowen GD, Nambiar EKS (eds) (1984) Nutrition of plantation forests. Academic, London
Brouder S, Volenec JJ, Turco R, Smith DR, Gebisa E (2009) Nutrient use efficiency in bioenergy cropping systems: critical research questions. The Proceedings of the international plant nutrition colloquium XVI, Paper 1430, 10p. Available via http://repositories.cdlib.org/ipnc/xvi/1430. Accessed October 2010
Cao T, Valsta L, Härkönen S, Saranpää P, Mäkelä A (2008) Effects of thinning and fertilization on wood properties and economic returns for Norway spruce. For Ecol Manage 256:1280–1289
Chalmers AG (2001) A review of fertilizer, lime and organic manure use on farm crops in Great Britain from 1983 to 1987. Soil Use Manage 17:254–262
Chen Q, Zhang X, Zhang H, Chrisie P, Li X, Horlacher D, Liebig H-P (2004) Evaluation of current fertilizer practices and soil fertility in vegetable production in the Beijing region. Nutr Cycl Agroecosyst 69:51–58
Chen CR, Condron LM, Xu ZH (2008) Impacts of grassland afforestation with coniferous trees on soil phosphorus dynamics and associated microbial processes: a review. For Ecol Manage 255:396–409
Comerford NB (2002) Forest soils. In: Lal R (ed) Encyclopedia of soil science, Marcel Dekker, pp 586–589
Cordell D, Drangert J-O, White S (2009) The story of phosphorus: global food security and food for thought. Global Environ Change 19:292–305
Crews TE, Peoples MB (2005) Can the synchrony of nitrogen supply and crop demand be improved in legume and fertilizer-based agroecosystems? A review. Nutr Cycl Agroecosyst 72:101–120
Cromer RN, Dargavel JB, Henderson VT, Nelson PF (1977) More pulpwood from less land. APPITA 31:49–54
Dear BS, Helyar KR, Muller WJ, Loveland B (1992) The P fertilizer requirements of subterranean clover, and the soil P status, sorption and buffering capacities from two P analyses. Aust J Soil Res 30:27–44
Dobermann A (2007) Nutrient use efficiency—measurement and management. Paper presented at international fertilizer industry workshop on fertilizer best management practices 7–9 March 2007, Brussels, Belgium. Available via http://www.fertilizer.org/ifacontent/download/7334/115716/version/1/file/2007_IFA_FBMP_part_1_dobermann.pdf Accessed 6 Oct 2009
Dobermann A, Arkebauer T, Cassman K, Lindquist J, Specht J, Walters D, Yang H (2002) Understanding and managing corn yield potential. Proceedings of the fertilizer industry round table, Charleston, South Carolina. The Fertilizer Industry Round Table, Forest Hill, Maryland, October. Available via http://www.potafos.org/far/farguide.nsf/926048f0196c9d4285256983005c64de/8a344a0a2ca7d90f86256e8c006a2c82/$FILE/NE-11F%202002%20Annual%20Report.doc. Accessed in 6 Oct 2009
Donald CM, Hamblin J (1976) The biological yield and harvest index of cereals as agronomic and plant breeding criteria. Agron 28:361–405
Downes GM, Nyakuengama JG, Evans R, Northway R, Blakemore P, Dickson RL, Lausberg M (2002) Relationship between wood density, microfibril angle and stiffness in thinned and fertilized Pinus radiata. IAWA J 23:253–265
Ericsson T, Göansson A, van Oene H, Gobran G (1995) Interactions between aluminium, calcium and magnesium—impacts on nutrition and growth of forest trees. Ecol Bulletins (Copenhagen) 44:191–196
Eriksson E, Gillespie AR, Gustavsson L, Langvall O, Olsson M, Sathre R, Stendahl J (2007) Integrated carbon analysis of forest management practices and wood substitution. Can J For Res 37:671–681
Escalante-Pérez M, Lautner S, Nehls U, Selle A, Teuber M, Schnitzler J-P, Teichmann T, Fayyaz P, Hartung W, Polle A, Fromm J, Hedrich R, Ache P (2009) Salt stress affects xylem differentiation of grey poplar (Populus × canescens). Planta 229:299–309
Evers F-H (1991) Forest fertilization—present state and history with special reference to South German conditions. Fert Res 27:71–86
FAO (2005) The state of food and agriculture. Available via http://www.fao.org/docrep/008/a0050e/a0050e09.htm. Accessed 14 Jan 2010
FAO (2006) Global forest resources assessment 2005. FAO Forestry Paper 147. Available via http://www.fao.org/DOCREP/008/a0400e/a0400e00.htm. Accessed 18 Jan 2010
FAO (2008) Current world fertilizer trends and outlook to 2011/12. FAO, Rome, 2008. Available via ftp://ftp.fao.org/agl/agll/docs/cwfto11.pdf. Accessed 18 Jan 2010
Fife DN, Nambiar EKS (1997) Changes in the canopy and growth of Pinus radiata in response to nitrogen supply. For Ecol Manage 93:137–152
Fife DN, Nambiar EKS, Saur E (2008) Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment. Tree Physiol 28:187–196
Fixen PE (2009) World fertilizer nutrient reserves—a view to the future. Better Crops 93(3):8–11
Forrester DI, Bauhus J, Cowie AL, Vanclay JK (2006) Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: a review. For Ecol Manage 233:211–230
Fox TR, Allen HL, Albaugh TJ, Rubilar R, Carlson CA (2007) Tree nutrition and forest fertilization of pine plantations in the southern United States. South J Appl For 31:5–11
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892
Garnett TP, Shabala SN, Smethurst PJ, Newman IA (2001) Simultaneous measurement of ammonium, nitrate and proton fluxes along the length of eucalypt roots. Plant Soil 236:55–62
Garnett TP, Shabala SN, Smethurst PJ, Newman IA (2003) Kinetics of ammonium and nitrate uptake by eucalypt roots and associated proton fluxes measured using ion selective microelectrodes. Funct Plant Biol 30:1165–1176
Gonçalves JLM (ed) (2004) Forest nutrition and fertilization. IPEF, Piracicaba
Gonçalves JLM, Barros NF (1999) Improvement of site productivity for short-rotation plantations in Brazil. Bosque 20:89–106
Gonçalves JLM, Barros NF, Nambiar EKS, Novais RF (1997) Soil and stand management for short-rotation plantations. In: Nambiar EKS, Brown AG (eds) Management of soil, nutrients and water in tropical plantation forests. ACIAR Monograph No. 43, pp 379–417
Gonçalves JLM, Stape JL, Laclau J-P, Bouillet RJ (2008) Assessing the effects of early silvicultural management on long-term site productivity of fast-growing eucalypt plantations: the Brazilian experience. Southern Forests 70:105–118
Gopalakrishnan G, Negri MC, Wang M, Wu M, Snyder SW, Lafreniere L (2009) Biofuels, land, and water: a systems approach to sustainability. Environ Sci Technol 43:6094–6100
Graham JH, Miller RM (2005) Mycorrhizas: gene to function. Plant Soil 274:79–100
Groot CC, Marcelis LFM, van den Boogaard R, Lambers H (2002) Interactive effects of nitrogen and irradiance on growth and partitioning of dry mass and nitrogen in young tomato plants. Funct Plant Biol 29:1319–1328
Hanks J, Ritchie JT (eds) (1991) Modeling plant and soil systems. Agronomy 31, Am Soc Agron, Wisconsin, pp 287–321
Hawkins BJ, Burgess D, Mitchell AK (2005) Growth and nutrient dynamics of western hemlock with conventional or exponential greenhouse fertilization and planting in different fertility conditions. Can J For Res 35:1002–1016
Houba VJG, Temminghoff EJM, Gaikhorst GA, van Vark W (2000) Soil analysis procedures using 0.01 M calcium chloride as extraction reagent. Commun Soil Sci Plant Anal 31:1299–1396
Huang W (2009) Factors contributing to the recent increase in U.S. fertilizer prices, 2002–08. USDA Economic Research Service Report AR-33
Hüttermann A (1985) The effects of acid deposition on the physiology of the forest ecosystem. Experientia 41:584–590
Ingestad T (1971) A definition of optimum nutrient requirements in birch seedlings. II. Physol Plant 23:118–125
Ingestad T (1982) Relative addition rate and external concentration: driving variables used in plant nutrition research. Plant Cell Environ 5:443–453
Johnston AE (1994) The Rothamsted classical experiments. In: Leigh RA, Johnston AE (eds) Long-term experiments in agricultural and ecological sciences. CAB International, Wallingford, pp 9–37
Jones DL (1998) Organic acids in the rhizosphere—a critical review. Plant Soil 205:25–44
Keating BA, Carberry PS, Hammer GL et al (2003) An overview of APSIM, a model designed for farming systems simulation. Eur J Agron 18:267–288
Knowles AE (2007) Aspects of the nutrition and physiology of Eucalyptus globulus. PhD Thesis, Univ. Tasmania
Knowles A, Shabala S (2004) Overcoming the problem of non-ideal liquid ion exchanger selectivity in microelectrode ion flux measurements. J Membrane Biol 202:51–59
Kreutzer K (1995) Effects of forest liming on soil processes. Plant Soil 168–169:447–470
Kronzucker HJ, Siddiqi MY, Glass ADM (1997) Conifer root discrimination against soil nitrate and the ecology of forest succession. Nature 385:59–61
Landsberg JJ, Waring RH (1997) A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For Ecol Manage 95:209–228
Langer K, Ache O, Geiger D, Stinzing A, Arend M, Wind C, Regan S, Fromm J, Hedrich R (2002) Poplar potassium transporters capable of controlling K+ homeostasis and K+-dependent xylogenesis. Plant J 32:997–1009
Linder S (1995) Foliar analysis for detecting and correcting nutrient imbalances in Norway spruce. Ecol Bull (Copenhagen) 44:178–190
Löfgren S, Cory N, Zetterberg T, Larsson P-E, Kronnäs V (2009) The long-term effects of catchment liming and reduced sulphur deposition on forest soils and runoff chemistry in southwest Sweden. For Ecol Manage 258:567–578
Macduff JH, Jarvis SC, Larsson C-M, Oscarson P (1993) Plant growth in relation to the supply and uptake of NO −3 : a comparison between relative addition rate and external concentration as driving variables. J Exp Bot 44:1475–1484
Matzner E, Ulrich B (1985) Implications of the chemical soil conditions for forest decline. Experientia 41:578–584
May B, Carlyle C, Lowther R, Keeley P, Bernie C, Michael M (2009a) A decision support system for maximizing profit from nutrient management of mid-rotation radiata pine. Forests and Wood Products Australia Project PNO: 03.3907. Available via http://www.fwpa.com.au/environment.aspx?s=2&pn=PN03.3907. Accessed 18 Jan 2010
May B, Smethurst P, Carlyle C, Mendham D, Bruce J, Baillie C (2009b) Review of Fertiliser Use in Australian Forestry. Forest and Wood Products Australia Report PRC072-0708. Available via http://www.fwpa.com.au/latestreports.aspx?s=4&pn=PRC072-0708. Accessed 6 Oct 2009
McBroom MW, Beasley RS, Chango M (2008) Water quality effects of clearcut harvesting and forest fertilization with best management practices. J Environ Qual 37:114–124
McDowell RW (ed) (2008) Environmental impacts of pasture-based farming. CAB International
McLaughlin MJ, Reuter DJ, Rayment GE (1999) Soil testing—principles and concepts. In: Peverill KI, Sparrow LA, Reuter DJ (eds) Soil analysis and interpretation manual. CSIRO, Collingwood, pp 1–21
Meiwes KJ, Mindrup M, Khanna PK (2002) Retention of Ca and Mg in the forest floor of a spruce stand after application of various liming materials. For Ecol Manage 159:27–36
Mendham D, Smethurst P, Holz G, Menary R, Grove T, Weston C, Baker T (2002) Soil analyses as indicators of P status in young Eucalyptus nitens and E. globulus plantations. Soil Sci Soc Am J 66:959–968
Min XJ, Siddiqi MY, Guy RD, Glass ADM, Kronzucker HJ (2000) A comparative kinetic analysis of nitrate and ammonium influx in two early-successional tree species of temperate and boreal forest ecosystems. Plant Cell Environ 23:321–328
Nambiar EKS (ed) (2008) Site management and productivity in tropical plantation forests: proceedings of workshops in Piracicaba (Brazil) 22–26 Nov 2004 and Bogor (Indonesia) 6–9 November 2006. Center for International Forestry Research (CIFOR). Available via http://www.cifor.cgiar.org/Knowledge/Publications/Detail?pid=774. Accessed 14 Jan 2010
Nambiar EKS, Brown AG (eds) (1997) Management of soil, nutrients and water in tropical plantation forests. Australian Centre for International Agricultural Research Monograph No. 43
Nyakuengama JG, Downes GM, Ng J (2002) Growth and wood density responses to later-aged fertilizer applications in Pinus radiata. IAWA J 23:431–448
O’Connell AM and Sankaran KV (1997) Organic matter accretion, decomposition and mineralisation. In: Nambiar EKS, Brown AG (eds) Management of soil, nutrients and water in tropical plantation forests. ACIAR Monograph No. 43, pp 297–337
Osborne R, Smethurst P, Wilkinson A (2001) Soil fertility assessment: survey and training using a quick-test meter. Cooperative Research Centre for Sustainable Production Forestry, Hobart, Tech Rep 61
Passioura J (2004) Water-use efficiency in farmers’ fields. In: Bacon M (ed) Water-use efficiency in plant biology. Blackwell, Oxford, pp 302–321
Pintro JC, Taylor GJ, Matumoto-Pintro T (2004) Optimizing exponential growth of Triticum aestivum by application of the relative addition rate (RAR) technique utilizing a computer-controlled nutrient delivery system. Braz J Plant Physiol 16:163–169
Pritchett WL (1979) Properties and management of forest soils. Wiley, New York
Pritchett WL, Llewellyn WR, Swinford KR (1961) Response of slash pine to colloidal phosphate fertilization. Soil Sci Soc Am J 46:640–644
Pulito AP (2009) Resposta à fertilização nitrogenada e estoque de nitrogênio biodisponível em solos usados para plantações de Eucalyptus [Response to nitrogen fertilization and stock of available nitrogen in soils used in eucalyptus crops]. MSc (Forestry) Thesis, University of São Paulo, Piracicaba
Raghothama KG (1999) Phosphate acquisition. Ann Rev Plant Physiol Plant Mol Biol 50:665–693
Raven JA (2001) An aquatic perspective on the concepts of Ingestad relating plant nutrition to plant growth. Physiol Plant 113:301–307
Raymond CA (1998) Wood properties, silviculture and growth: a review of published literature for eucalypts. Cooperative Research Centre for Temperature Hardwood Forestry, Hobart, Tech Rep 6
Rennie PJ (1955) The uptake of nutrients of mature forest growth. Plant Soil 7:49–95
Richardson AE, Barea J-M, McNeill AM, Prigent-Combaret C (2009) Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 321:305–339
Ross DS, Matschonat G, Skyllberg U (2008) Cation exchange in forest soils: the need for a new perspective. European J Soil Sci 59:1141–1159
Sands PJ, Smethurst PJ (1995) Modelling nitrogen uptake in Ingestad units using Michaelis-Menten kinetics. Aust J Plant Physiol 22:823–831
Schachtman DP, Shin R (2007) Nutrient sensing and signaling: NPKS. Ann Rev Plant Biol 58:47–69
Schönau APG, Herbert MA (1989) Fertilizing eucalypts at plantation establishment. For Ecol Manage 29:221–244
Smethurst PJ (2000) Soil solution and other soil analyses as indicators of nutrient supply: a review. For Ecol Manage 138:397–411
Smethurst P (2004) Nutritional physiology of trees. In: Burley J, Evans J, Youngquist JA (eds) Encyclopedia of forest sciences, Oxford, pp 1616–1622
Smethurst PJ (2007) Nitrogen cycling in forests with inter-specific competition: models and knowledge in relation to temperate eucalypt and pine plantations. Cooperative Research Centre for Forestry Tech Rep 172. Available via http://www.crcforestry.com.au/publications/technical-reports/index.html. Accessed 12 Oct 2009
Smethurst PJ, Wang BP (1998) Soil solution phosphorus and Eucalyptus nitens roots in NP-treated microsites in highly phosphorus-fixing soil. N Z J Forestry Sci 28:140–151
Smethurst PJ, Herbert AM, Ballard LM (2001) Fertilization effects on soil solution chemistry in three eucalypt plantations. Soil Sci Soc Am J 65:795–804
Smethurst PJ, Baillie C, Cherry M, Holz G (2003) Fertilizer effects on LAI and growth of four Eucalyptus nitens plantations. For Ecol Manage 176:531–542
Smethurst P, Holz H, Moroni M, Baillie C (2004a) Nitrogen management in Eucalyptus nitens plantations. For Ecol Manage 193:63–80
Smethurst P, Mendham D, Battaglia M, Misra R (2004b) Simultaneous prediction of nitrogen and phosphorus dynamics in a Eucalyptus nitens plantation using linked CABALA and PCATS models. In: Borralho N et al. (eds) Eucalyptus in a changing world, Proc. IUFRO Conference, Aveiro, Portugal, 11–15 Oct. 2004, pp 565–568
Smethurst PJ, Knowles A, Wilkinson A, Churchill K, Lyons A (2007) Soil and foliar chemistry associated with potassium deficiency in Pinus radiata. Can J For Res 37:1093–1105
Smith FW, Loneragan JF (1997) Interpretation of plant analysis: concepts and principles. In: Reuter DJ, Robinson JB (eds) Plant analysis an interpretation manual. CSIRO, Collingwood, pp 3–33
Stape JL, Binkley D, Ryan MG (2004) Eucalyptus production and the supply, use and efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For Ecol Manage 193:17–31
Subbarao GV, Ito O, Berry WL, Wheeler RM (2003) Sodium—a functional plant nutrient. Crit Rev Pl Sci 22:391–416
Sun J, Dai SX, Wang RG, Chen SL, Li NY, Zhou XY, Lu CF, Shen X, Zheng XJ, Hu ZM, Zhang ZK, Song J, Xu Y (2009) Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol 29:1175–1186
Sverdrup H, de Vries W (1994) Calculating critical loads for acidity with the simple mass balance method. Wat Air Soil Poll 72:143–162
Tamm CO (1995) Towards an understanding of the relations between tree nutrition, nutrient cycling and environment. Plant Soil 168–169:21–27
Tinker PB, Nye PH (2000) Solute movement in the rhizosphere. Oxford University Press
Turnbull CRA, Beadle CL, West PW, Cromer RN (1994) Copper deficiency a probably cause of stem deformity in fertilized Eucalyptus nitens. Can J For Res 24:1434–1439
Turvey ND, Smethurst PJ (1983) Nitrogen fixing plants in forest plantation management. In: Gordon JC, Wheeler CT (eds) Biological nitrogen fixation in forest ecosystems: foundations and applications. Martinus Nijhof, The Hague, pp 233–260
Vitousek PM (1982) Nutrient cycling and nutrient use efficiency. Am Nat 119:553–572
Vitousek PM, Melillo JM (1979) Nitrate losses from disturbed forests: patterns and mechanisms. For Sci 25:605–619
Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler DW, Schlesinger WH, Tilman DG (1997) Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750
Vitousek PM, Naylor R, Crews T, David MB, Drinkwater LE, Holland E, Johnes PJ, Katzenburger J, Martinelli LA, Nziguheba G, Ojima D, Palm CA, Robertson GP, Sanchez PA, Townsend AR, Zhang FS (2009) Nutrient imbalances in agricultural development. Sci 324:1519–1520
Wells CG, Jorgensen JR (1975) Nutrient cycling in loblolly pine plantations. In: Bernier B, Winget CH (eds) Forest soils and forest land management, Les Presses de l’Université Laval, pp 137–158
White DA, Crombie SD, Kinal J, Battaglia M, McGrath JF, Mendham DS, Walker SN (2009) Managing productivity and drought risk in Eucalyptus globulus plantations in south-western Australia. For Ecol Manage 259:33–44
Williams DR, Potts BM, Smethurst PJ (2003) Promotion of flowering in Eucalyptus nitens by paclobutrazol was enhanced by nitrogen fertilizer. Can J For Res 33:74–81
Zobel B (1992) Silvicultural effects on wood properties. IPEF International, Piracicaba 2:31–38
Acknowledgements
Juergen Bauhus, Leonardo Gonçalves, and Tom Fox assisted in sourcing some of the literature. Comments by Daniel Mendham, Shaun Lisson, Barrie May, Christiane Smethurst, and Don White on earlier drafts of the manuscript were much appreciated. The author also appreciates the assistance provided to him to present this paper to the IPI-OUAT-IPNI International Symposium ‘Potassium Role and Benefits in Improving Nutrient Management for Food Production and Reduced Environmental Damages’, 5–7 November 2009, Bhubaneswar, Orissa, India.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Ismail Cakmak.
Rights and permissions
About this article
Cite this article
Smethurst, P.J. Forest fertilization: Trends in knowledge and practice compared to agriculture. Plant Soil 335, 83–100 (2010). https://doi.org/10.1007/s11104-010-0316-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0316-3