
Abstract
The extent of resection and the intrinsic biological aggressiveness of the tumor have been repeatedly identified as the primary determinants of meningioma recurrence. Invasive growth limits the resectability of meningiomas. Tumor invasion is mediated by matrix metalloproteases and their inhibitors such as uPA and PAI-1. In some cancers uPA expression is controlled in part by promoter methylation. In the work reported in this paper we investigated the role of uPA/PAI-1 expression and methylation of the uPA promoter in meningiomas. Sixty-five tumor tissue samples (WHO grade I: 26, grade II: 27, grade III: 12) from 58 patients were analyzed for uPA and PAI-1 protein content using a commercially available ELISA kit. For uPA promoter methylation analysis, a 365-bp promoter fragment was amplified by PCR after bisulfite treatment and subjected to a methylation-sensitive restriction digest with AciI. Pertinent clinical data were retrieved from the patients’ charts. uPA and PAI-1 protein expression correlated significantly with WHO grade (uPA: P < 0.033, PAI-1: P < 0.001). High (>6 ng/ml = median) PAI-1 levels were seen more frequently in tumors with brain invasion (P = 0.006) and proved a significant predictor of the patients’ prognosis (Kaplan–Meier estimates of progression-free survival: P = 0.004). Increased methylation of the uPA promoter was found to correlate significantly with lower levels of uPA expression. Our data suggest PAI-1 (and possibly to a lesser degree uPA) as potential prognostic markers in meningiomas. uPA expression in meningiomas might, in part, be controlled by promoter methylation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Meningiomas are the second most common CNS tumors. Their incidence has been reported to be 6 per 100,000 [1]. They are twice as common in females as in males, and the incidence is highest after the fifth decade of life. In comparison with gliomas, clinical and basic science researchers have given relatively little attention to these growths. The WHO classification distinguishes between benign WHO grade I, atypical WHO grade II, and anaplastic WHO grade III tumors. Five-year recurrence of 38–52% has been described for atypical meningiomas, and 5-year recurrence for anaplastic meningiomas may be as high as 50–84% [2]. Five-year overall survival of 78 and 44% has been reported for atypical and anaplastic meningiomas, respectively [3].
Surgical resection is still regarded as standard treatment for most tumors. Radiosurgery is a good option for small tumors or (progressive) residual tumor whenever (repeat) surgery is deemed too risky, and for tumors in specific locations, for example the cavernous sinus. Conventional radiotherapy is usually prescribed for anaplastic (and often also for atypical) meningiomas [4]. Various systemic therapies have been tried for tumors which have failed standard therapeutic regimens. However, these treatments have met with modest success at best. There are some data to suggest a role for targeted therapies in the future [5, 6].
The extent of resection and the intrinsic biological aggressiveness of the tumor have been repeatedly identified as the primary determinants of meningioma recurrence [7]. Invasive growth limits the resectability of meningiomas. Inclusion of brain invasion as a criterion for atypia in the 2007 WHO classification is a significant modification of the preceding version [8]. However, only a few studies have been performed on meningioma invasion. Little is known about the underlying molecular mechanisms.
Tumor invasion is associated with extracellular matrix remodeling and involves various proteases, for example the urokinase plasminogen activator (uPA) system. uPA and its inhibitor PAI-1 are involved in extracellular matrix proteolysis but also in cellular adhesion and migration by interacting with vitronectin and integrins [9, 10]. uPA/PAI-1 have been implicated as important in glioma invasion and malignant progression [11, 12]. Only a few similar investigations in meningiomas have been conducted [13, 14]. uPA gene expression is regulated in part through promoter methylation in breast cancer cell lines and possibly in human breast carcinomas also [15]. The methylation status of the uPA promoter has been shown to affect prostate cancer invasion and growth in vitro and in vivo [16].
In the work discussed in this paper we investigated the importance of uPA/PAI-1 expression and uPA promoter methylation in a collection of meningiomas with a high percentage of atypical (WHO grade II) and anaplastic (WHO grade III) tumors. Various clinicopathological data (WHO grade, MIB-1 labeling index, tumor localization, tumor multiplicity, brain invasion, recurrence, progression free and overall survival) were tested for potential associations with uPA and PAI-1 expression.
Materials and methods
Patients and clinical data
For this investigation we studied 65 tissue samples (WHO grade I: 26, WHO grade II: 27, WHO grade III: 12) from 58 patients (66% female) who underwent surgery for an intracranial meningioma in the Department of Neurosurgery at the University of Bonn between 1996 and 2005. Tumors were selected from the Department’s tumor tissue bank. The patients’ age ranged from 17.8 to 80.3 years with a median age of 60 years at the time of surgery. The relative frequency of WHO grade I, grade II, and grade III tumors among 15 patients <50 years was 27, 53, and 20%, among 33 patients 50–70 years 52, 27, and 21%, and among 17 patients >70 years 29, 58, and 12%.
Fifteen tumor samples (23%) were obtained at surgery for recurrent tumor diagnosed between 12 and 516 months after the initial surgery. Tumor tissue from both primary and recurrent meningioma was available in one case. Forty-four patients (76%) were diagnosed with a single tumor, six patients (10%) with 2 or 3, and eight patients (14%) with >3 meningiomas. Twenty-two tumors (34%) were located at the cranial base. Twenty-six meningiomas (40%) were larger than 5 cm. MRI investigations showed significant brain edema (>0.5 cm on T2 weighted MR images) in 22 (34%) patients. There were 38 Simpson grade 1 (59%), 21 Simpson grade 2 (32%), and five Simpson grade 3 (8%) resections. In one case, only a Simpson grade 4 resection was possible.
Pertinent clinical data were retrieved from the patients’ charts. Survival analysis were restricted to patients (n = 47) who were included in the study at the time of their first surgery. Median follow-up after the first surgery in this group was 48 months (mean 44, range 0–137 months).
Neuropathology
All histopathological diagnoses were made at the Department of Neuropathology/German Brain Tumor Reference Center at the University of Bonn. Brain invasion in the absence of histological atypia was considered sufficient to enable diagnosis of an atypical meningioma WHO grade II. Brain invasion and bone invasion were recorded if histologically confirmed. MIB-1 (Ki-67) labeling indices were obtained for 58 (89%) meningiomas, and tumors were assigned to three categories: (1) MIB-1 <5%, (2) MIB-1 = 5–10%, (3) MIB-1 >10%.
uPA/PAI-1 expression analysis
Tumor tissue specimens were obtained at surgery and immediately shock frozen in liquid nitrogen. Samples were later stored at −80°C. Frozen tumor samples (100 mg) were homogenized in tris buffered saline/1% Triton X-100. uPA and PAI-1 protein contents were determined using the Femtelle uPA/PAI-1 Elisa Kit (American Diagnostica, Pfungstadt, Germany) as described by the manufacturer. The Femtelle kit is used routinely as part of the work-up for nodal negative breast cancers treated in the Departments of Gynecology at Hannover Medical School. Protein concentrations were measured using standard methods (BCA kit; Pierce, Rockford, USA).
uPA promoter methylation analysis
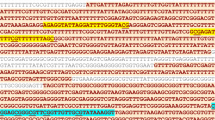
DNA was extracted using standard methods (QIAamp DNA mini kit; Qiagen, Hilden, Germany) from shock frozen tumor tissue and subjected to bisulfite treatment with the EpiTect bisulfite kit (Qiagen) in accordance with the manufacturer’s recommended procedure . A 365 bp fragment of the uPA promoter (−430 to −66; NCBI Reference Sequence: NG_011904.1: 4596–4960) was PCR amplified (forward primer: 5′-AGA TTT TAT AGT TTT ATT AGT-3′, reverse primer: 5′-CTC CCT CCC CTA TCT TAC AA-3′). Primers were designed to bind specifically to bisulfite-modified DNA. Bisulfite treatment of the DNA converts unmethylated cytosine residues into uracil but leaves 5-methylcytosine unaffected. Uracil residues resulting from the previous bisulfite treatment are converted to thymidine during subsequent PCR analysis. Hence, the base sequence of the final 365-bp PCR product will vary with the DNA methylation status of the template. The sequence of the primers did not include CpG dinucleotides, in order to for enable amplification of the DNA fragment irrespective of DNA methylation status.
The 365 bp uPA promoter fragment contains 19 potential CpG methylation sites. Sites 2–19 correspond to the CpG dinucleotides termed 1–18 by Pakneshan et al.[15] Five of the CpG dinucleotides (sites 1, 2, 14, 15, and 16) are part of AciI recognition sites, i.e. these AciI sites are destroyed by bisulfite treatment unless protected by CpG methylation. Hence, the band pattern after AciI restriction will reflect the degree of methylation of this region. Increasing levels of DNA methylation will result in more complete AciI digests, i.e. will increase the relative proportion of smaller DNA fragments.
Agarose gel electrophoresis and sequence analysis identified the following major bands in controls (normal brain and normal brain after CpG methylation with M.SssI) and in the meningioma samples: 365 and 342 bp (original uncut PCR product and NCBI Reference Sequence: NG_011904.1: 4619–4960/CpG site 1 to 3′ end), 298 bp (NCBI Reference Sequence: NG_011904.1: 4663–4960/CpG site 2 to 3′ end), 257 and 247 bp (NCBI Reference Sequence: NG_011904.1: 4663–4909/CpG sites 2 to 14 and 4663–4919/CpGH sites 2 to 16), and several smaller fragments. On the basis of the relative densitometric intensities of the bands, samples could be assigned to three groups reflecting increasing levels of methylation (Fig. 1):
-
1
strong 365/342 bp doublet and all other bands <30%;
-
2
strong 365/342 bp doublet ≥ strong 298 bp band, all other bands <30%; and
-
3
365/342 bp doublet <298 bp band, all other bands <30% or small bands (257, and 247 bp etc.) >50%.

Differential methylation of a 365 bp uPA promoter fragment in meningiomas. Agarose gel electrophoresis identified the following major bands after bisulfite treatment, PCR amplification, and methylation-sensitive AciI digest: 365 and 342 bp (original uncut PCR product and NCBI Reference Sequence: NG_011904.1: 4619–4960/CpG site 1 to 3′ end), 298 bp (NCBI Reference Sequence: NG_011904.1: 4663–4960/CpG site 2 to 3′ end), 257 and 247 bp (NCBI Reference Sequence: NG_011904.1: 4663–4909/CpG sites 2 to 14 and 4663–4919/CpGH sites 2 to 16). On the basis of the relative densitometric intensities of the bands, samples could be assigned to three categories reflecting increasing levels of methylation: (1) strong 365/342 bp doublet and all other bands <30% (tumors 2582 and 2200); (2) strong 365/342 bp doublet ≥strong 298 bp band, all other bands <30% (tumor 2517); (3) 365/342 bp doublet <298 bp band, all other bands <30% (tumors 2107 and 2098) or small bands (257, and 247 bp etc.; tumors 2292 and 2324) >50%. M 100 bp marker; N normal brain; M, after methylation with M.SssI
Statistical analysis
Statistical analysis was supported by commercially available software (SPSS 17.0). For survival analysis Kaplan–Meier estimates and log-rank tests were used. The level of significance was set at P < 0.05.
Results
uPA/PAI-1 protein expression and uPA promoter methylation in meningiomas
uPA and its inhibitor PAI-1 have been implicated in the expression of an invasive phenotype and malignant progression of various cancers including some brain tumors. To study the involvement of uPA/PAI-1 in meningiomas in more detail we investigated 65 of these tumors for uPA/PAI-1 protein expression. Tumors were found to contain variable amounts of uPA (mean: 0.41 ng/mg ± 0.61, median: 0.2 ng/mg, range: <0.1–4.58 ng/mg) and PAI-1 protein (mean: 30.2 ng/mg ± 102.3, median: 5.7 ng/mg, range: 0.7–625.0 ng/mg). There was a significant correlation between uPA and PAI-1 expression (Pearson correlation coefficient: 0.676, P < 0.001). However, very high levels of expression of the uPA inhibitor PAI-1 were also observed in some tumors with low or moderate levels of expression of uPA. Analysis of paired primary and recurrent tumor tissue samples was possible in one patient only (primary vs. recurrent tumor: 0.16 vs. 1.4 ng uPA/mg protein, and 22 vs. 11 ng PAI-1/mg protein).
Pakneshan et al. have provided correlative and functional data linking methylation of the uPA promoter to uPA expression in breast and prostate cancer [14, 15]. We therefore investigated whether uPA expression also varies with uPA promoter methylation in meningiomas. uPA promoter methylation was assessed semiquantitatively on the basis of the relative intensities of the bands observed after bisulfite treatment, PCR, and a methylation-sensitive restriction digest. Overall, uPA levels decreased with increased levels of uPA methylation. Mean/median uPA levels for categories 1, 2, and 3 were 0.61 ± 0.43/0.5, 0.37 ± 0.79/0.2, and 0.21 ± 0.12/0.2 ng/mg protein, respectively (Fig. 2, ANOVA, n.s.). Using the overall median (i.e. 0.2 ng uPA/mg protein) as a cut-off we found a significant inverse correlation between the degree of uPA promoter methylation and uPA expression (Spearman’s rho: −0.348, P = 0.007). These data may indicate that uPA expression in some meningiomas is indeed down-regulated in part by promoter methylation.

Relationship between uPA protein expression and uPA promoter methylation. Mean and median uPA levels decrease with increasing methylation (i.e. from methylation category 1–3, see Fig. 1). This association proved statistically significant when comparing the relative proportion of tumors with high (>median = 0.2 ng/mg protein) levels of uPA between the three methylation categories (Spearman’s rho: −0.348, P = 0.007)
Clinicopathological correlations
Next, we analyzed our series for potential correlations between uPA/PAI-1 expression and various clinicopathological data. Importantly, mean uPA and PAI-1 protein amounts were found to increase with the WHO grade of the tumors (mean/median for WHO grade I: 0.23 ± 0.15/0.2, and 6.75 ± 8.72/4.15, grade II: 0.42 ± 0.33/0.3, and 8.30 ± 6.21/7.80, grade III: 0.78 ± 1.27/3.5, and 130.20 ± 217.21/22.85 ng uPA and PAI-1/mg protein; ANOVA, P = 0.033, PAI-1: P <0.001, see Fig. 3a, b). Using the respective median (0.2 ng uPA/mg protein, and 6 ng PAI-1/mg protein) as a cut-off, the proportion of tumors with high levels of PAI-1 (but not uPA) was found to increase significantly with tumor grade (uPA, Spearman’s rho: 0.228, n.s.; PAI-1, Spearman’s rho: 0.251, P = 0.044). Atypical meningiomas of WHO grade II had intermediate levels of uPA compared with WHO grade I and III tumors. PAI-1 expression was relatively low in benign and atypical, but high in anaplastic meningiomas. Mean uPA and PAI-1 levels were found to increase with the MIB-1 labeling index of the tumors, however, findings were statistically significant for PAI-1 only (MIB-1 <5%: 0.27 ± 0.11, and 6.88 ± 8.67, MIB-1 = 5–10%: 20.88 ± 51.81, and 0.47 ± 0.38, MIB-1 >10%: 32.76 ± 108.04, and 0.69 ± 1.22 ng uPA/mg and PAI-1 protein; ANOVA, uPA: n.s., PAI-1: P = 0.046). The proportion of tumors with high (i.e. >median) uPA/PAI-1 levels did not increase significantly with MIB-1 labeling index.

Relationship between uPA A and PAI-1 B protein levels and the WHO grade of meningiomas (ANOVA, uPA: P = 0.033, PAI-1: P < 0.001). Atypical meningiomas of WHO grade II had intermediate levels of uPA compared with WHO grade I and III tumors. PAI-1 expression was relatively low in benign (WHO grade I) and atypical (WHO grade II), but high in anaplastic (WHO grade III) meningiomas
Brain invasion was histologically verified in 15 cases, and bone invasion in nine tumors. Mean PAI-1 and uPA levels did not differ significantly between tumors with and without brain or bone invasion, but brain invasion was more frequent among tumors with high PAI-1 expression (using the median PAI-1 level = 6.0 ng/mg as a cut-off, Spearman’s rho: 0.337, P = 0.006). There were no correlations between uPA or PAI-1 expression and tumor size, tumor location, presence of brain edema, or multiplicity. uPA promoter methylation did not correlate with WHO grade, MIB-1 labeling index, brain or bone invasion, tumor size, tumor location, presence of brain edema, or multiplicity in our series.
We finally investigated uPA/PAI-1 expression and uPA promoter methylation for an association with patient survival. Kaplan–Meier estimates of progression-free (PFS) and overall survival (OS) using the median PAI-1 level (6 ng/mg) as a cut-off showed a highly significant correlation between PAI-1 expression and the patients’ prognosis (log rank test, PFS: P = 0.004, OS: P = 0.031; Fig. 4). No tumor containing <6 ng/ml PAI-1 recurred after a median follow-up of 47 months. PAI-1 expression >6 ng/ml seemed to be a better predictor of tumor recurrence and tumor-related death than extent of resection. There was no significant correlation between Simpson grade and the patients’ prognosis (log rank test, PFS and OS: P = n.s., see Fig. 4). No significant correlations were observed between uPA expression or uPA promoter methylation and patient survival.

Kaplan–Meier estimates of progression-free survival (PFS, upper panel) and overall survival (OS, lower panel) vs. PAI-1 protein expression (left, using the median PAI-1 level = 6 ng/mg as a cut-off), WHO grade (middle), and extent of resection (Simpson grade, right). There was a highly significant correlation between PAI-1 expression and the patients’ prognosis (log rank test, PFS: P = 0.004, OS: P = 0.031). No tumor containing <6 ng/ml PAI-1 recurred after a median follow-up of 47 months
Discussion
Plasminogen activators (PA) and their inhibitors are important in fibrinolysis, cell migration [17–19], angiogenesis, chemotaxis [20, 21] wound healing, embryogenesis, tumor progression, and invasion, and metastasis [22]. Active uPA catalyzes the conversion of inactive plasminogen to its active form plasmin. Plasmin degrades components of the extracellular matrix, for example laminin, fibronectin, fibrin and collagen, and activates MMPs and growth factors. PAI-1 is the most effective inhibitor of uPA.
uPA and PAI-1 expression have been studied in some detail in gliomas [11, 12]. In contrast, only a few similar investigations in meningiomas have been published [12, 13, 23]. Downregulation of uPA using siRNA and low-molecular-weight inhibitors has been shown to reduce migration, invasion, and tumorigenicity of a malignant meningioma cell line [14, 24]. This study details uPA/PAI-1 protein expression in a relatively large cohort (n = 65) including a sizable proportion of atypical (42%) and malignant (18%) meningiomas. Arai et al. [13] (using a different assay) reported similar amounts of uPA but generally lower levels of PAI-1 in their samples compared with our results. Levels of PAI-1 protein in our benign meningiomas and in the meningiomas studied by Sawaya and co-workers were comparable [12].
Importantly, this study links increased PAI-1 (and to a lesser degree uPA) expression with the malignant progression of meningiomas (i.e. a higher tumor grade), to brain invasion, and to tumor recurrence. There was a clear correlation between WHO grade and uPA/PAI-1 protein levels in our series. This confirms previous work by Arai et al. [13]. These authors have also observed higher uPA/PAI-1 expression in malignant than in benign meningiomas, albeit in a much smaller cohort (n = 18).
We also provide some evidence linking elevated PAI-1 (but not uPA) levels to brain invasion. Brain invasion was more frequent in tumors containing >6 ng/ml PAI-1. Siddique et al. and Nagashima et al. [23, 24] reported immunohistochemical data suggesting a correlation between increased uPA expression and an invasive phenotype in meningiomas. These data do not necessarily contradict our findings. There seems to be a general correlation between uPA/PAI-1 immunohistochemistry and ELISA data [13]. However, these two experimental techniques may measure different phenomena. Strong uPA staining of brain-invasive meningioma cells may reflect a functional role of uPA in brain invasion [23, 24], but increased uPA expression may be restricted to the invasion zone and may not necessarily translate into overall increased protein levels. Hence uPA ELISA data may not enable prediction of the invasive capacity of the tumor, but the inherent problems with the quantification of immunohistochemical data could still argue for the use of ELISA measurements as a biomarker for clinical use.
The current ASCO (American Society of Clinical Oncology) guidelines recommend uPA/PAI-1 testing for assessment of node-negative breast cancer [25]. This recommendation is supported by class I evidence [25, 26]. Some data also indicate a significant prognostic impact of PAI-1 levels in lung cancer [27, 28]. Our data suggest PAI-1 as a possible prognostic marker for meningiomas also. PFS and OS correlated strongly with PAI-1 levels in our series. No tumor with a PAI-1 protein content <6 ng/ml recurred, whereas 7/23 patients with meningiomas containing >6 ng/ml PAI-1 were diagnosed with recurrent tumor during follow-up. Of note, we used a commercially available test kit which has been approved by the FDA for routine use in breast cancer. Although this is a definite strength of our dataset, we also readily acknowledge that this test only measures protein content and not uPA/PAI-1 activity. PAI-1 levels >14 ng/ml have been associated with adverse prognosis in breast cancer [26]. This cut-off is similar to the 6 ng/ml cut-off used in our study.
On average, uPA levels are higher in breast and other cancers than in meningiomas. In breast cancer, only levels >3 ng/ml uPA have been correlated with an adverse prognosis [26]. Our data imply that high uPA levels (>3 ng/ml) may also carry an adverse prognosis in meningiomas. One of our tumors contained >3 ng/ml uPA. This patient was diagnosed with a malignant meningioma and succumbed to the disease only three months after surgery.
Finally, our study may shed some light on the regulation of uPA expression in meningiomas. There is evidence to suggest that the uPA gene is regulated in part through promoter methylation in breast cancer cell lines and in human breast cancer tissue. Lack of uPA expression in normal mammary epithelial cells is because of transcriptional suppression of uPA gene expression by DNA methylation of the uPA promoter [15]. The methylation status of the uPA promoter also regulates prostate cancer invasion and growth in vitro and in vivo [16]. Our data indicate that the same mechanism may also be involved in meningiomas. The extent of uPA promoter methylation was found to correlate inversely with uPA expression. Promoter CpG hypermethylation resulting in transcriptional silencing is an important epigenetic mechanism which is important in the formation and progression of various tumors [29, 30]. Functional polymorphisms of methionine metabolism which provide the methyl groups required for DNA methylation have been associated with meningioma formation and malignant progression [31]. Somewhat surprisingly, we did not observe any significant correlations between the clinicopathological data investigated and uPA promoter methylation. However, this largely parallels the findings for uPA expression (with the exception of the correlation between WHO grade and uPA protein levels). This may also reflect the role of mechanisms other than promoter methylation in the regulation of uPA expression.
In summary, this study provides a detailed description of uPA and PAI-1 protein expression and clinicopathological correlations in meningiomas. uPA expression might be controlled in part by promoter methylation. Similar to other cancers PAI-1 (and to a lesser extent uPA) seem to be involved in malignant progression and invasive growth. PAI-1 expression may prove to be a useful prognostic marker in meningiomas.
References
Longstreth WT, Dennis LK, McGuire VM (1993) Epidemiology of intracranial meningiomas. Cancer 72:639–648
Louis DN, Scheithauer BW, Budka B, von Deimling A, Kepes JJ (2000) Meningiomas. In: Kleihues P, Cavenee WK (eds) Pathology and genetics of tumours of the nervous system: World Health Organisation classification of tumours. IARC Press, Lyon, pp 176–184
Durand A, Labrousse F, Jouvet A, Bauchet L, Kalamaridès M, Menei P, Deruty R, Moreau JJ, Fèvre-Montange M, Guyotat J (2009) WHO grade II and III meningiomas: a study of prognostic factors. J Neurooncol 95(3):367–375
Hug EB, Devries A, Thornton AF et al (2000) Management of atypical and malignant meningiomas: role of high-dose, 3D-conformal radiation therapy. J Neurooncol 48(2):151–160
Black PM, Carroll R, Glowacka D, Riley K, Dashner K (1994) Platelet-derived growth factor expression and stimulation in human meningiomas. J Neurosurg 81(3):388–393
Lamszus K, Lengler U, Schmidt NO, Stavrou D, Ergün S, Westphal M (2000) Vascular endothelial growth factor, hepatocyte growth factor/scatter factor, basic fibroblast growth factor, and placenta growth factor in human meningiomas and their relation to angiogenesis and malignancy. Neurosurgery 46(4):938–947
Ho DM, Hsu CY, Ting LT, Chiang H (2002) Histopathology and MIB-1 labeling index predicted recurrence of meningiomas: a proposal of diagnostic criteria for patients with atypical meningioma. Cancer 94(5):1538–1547
Perry A, Louis DN, Scheithauer BW, Budka H, von Diemling A (2007) Meningiomas. In: Louis DN, Ohgaki H, Wiestler OD, Cavanee WK (eds) World Health Organization classification of tumours of the central nervous system, 4th edn. IARC, Lyon, pp 164–172
Deng G, Curriden SA, Hu G, Czekay RP, Loskutoff DJ (2001) Plasminogen activator inhibitor-1 regulates cell adhesion by binding to the somatomedin B domain of vitronectin. J Cell Physiol 189(1):23–33
Bajou K (2002) Role of PAI-1 plasminogen activator inhibitor in tumor invasion and angiogenesis. Bull Mem Acad R Med Belg 157(5–6):313–318
Bindal AK, Hammoud M, Shi WM, Wu SZ, Sawaya R, Rao JS (1994) Prognostic significance of proteolytic enzymes in human brain tumors. J Neurooncol 22(2):101–110
Sawaya R, Yamamoto M, Ramö OJ, Shi ML, Rayford A, Rao JS (1995) Plasminogen activator inhibitor-1 in brain tumors: relation to malignancy and necrosis. Neurosurgery 36:375–380
Arai Y, Kubota T, Nakagawa T, Kabuto M, Sato K, Kobayashi H (1998) Production of urokinase-type plasminogen activator (u-PA) and plasminogen activator inhibitor-1 (PAI-1) in human brain tumours. Acta Neurochirurgica 140:377–385
Kondraganti S, Gondi CS, McCutcheon I et al (2006) RNAi-mediated downregulation of urokinase plasminogen activator and its receptor in human meningioma cells inhibits tumor invasion and growth. Int J Oncol 28(6):1353–1360
Pakneshan P, Tetu B, Rabbani SA (2004) Demethylation of urokinase promoter as a prognostic marker in patients with breast carcinoma. Clinical Cancer Res 10:3035–3041
Pakneshan P, Xing RH, Rabbani SA (2003) Methylation status of uPA promoter as a molecular mechanism regulating prostate cancer invasion and growth in vitro and in vivo. FASEB J 17(9):1081–1088
Irigoyen JP, Munoz-Canoves P, Montero L, Koziczak M, Nagamine Y (1999) The plasminogen activator system: biology and regulation. Cell Mol Life Sci 56:104–132
Webb DJ, Nguyen DH, Gonias SL (2000) Extracellular signal-regulated kinase functions in the urokinase receptor-dependent pathway by which neutralization of low density lipoprotein receptor-related protein promotes fibrosarcoma cell migration and matrigel invasion. J Cell Sci 113:123–134
Jo M, Thomas KS, Somlyo AV, Somlyo AP, Gonias SL (2002) Cooperativity between the Ras-ERK and Rho-Rhi kinase pathways in urokinase-type plaminogen activator-stimulated cell migration. J Biol Chem 277(14):12479–12485
Blasi F (2001) u-PA and cell migration: urokinase receptor as a ligand for a chemotactic G-protein-coupled receptor. Haemostasis 31:59
Degryse B, Orlando S, Resnati M, Rabbani SA, Blasi F (2001) Urokinase/urokinase receptor and vitronectin/alpha (v) beta (3) integrin induce chemotaxis and cytoskeleton reorganization through different signaling pathways. Oncogene 20:2032–2043
Andeasen PA, Egelund R, Petersen HH (2000) The plasminogen activation system in tumor growth, invasion and metastasis. Cell Mol Life Sci 57:25–40
Siddique K, Yanamandra N, Gujrati M, Dinh D, Rao JS, Olivero W (2003) Expression of matrix metalloproteinases, their inhibitors and urokinase plasminogen activator in human meningiomas. Int J Oncol 22:289–294
Nagashima G, Fujimoto T, Suzuki R, Asai J, Itokawa H, Noda M (2006) Dural invasion of meningioma: a histological and immunohistochemical study. Brain Tumor Pathol 23(1):13–17
Harbeck N, Schmitt M, Paepke S, Allgayer H, Kates RE (2007) Tumor-associated proteolytic factors uPA and PAI-1: critical appraisal of their clinical relevance in breast cancer and their integration into decision-support algorithms. Crit Rev Clin Lab Sci 44(2):179–201
Pappot H, Pedersen AN, Brünner N, Christensen IJ (2006) The complex between urokinase (uPA) and its type-1 inhibitor (PAI-1) in pulmonary adenocarcinoma: relation to prognosis. Lung Cancer 51(2):193–200
Pedersen H, Brünner N, Francis D et al (1994) Prognostic impact of urokinase, urokinase receptor, and type 1 plasminogen activator inhibitor in squamous and large cell lung cancer tissue. Cancer Res 154(17):4671–4675
Jones PA, Baylin SB (2002) The fundamental role of epigenetic events in cancer. Nat Rev Genet 3(6):415–428
Baylin SB, Herman JG, Graff JR, Vertino PM, Issa JP (1998) Alterations in DNA methylation: a fundamental aspect of neoplasia. Adv Cancer Res 72:141–196
Semmler A, Simon M, Moskau S, Linnebank M (2008) Polymorphisms of methionine metabolism and susceptibility to meningioma formation: laboratory investigation. J Neurosurg 108(5):999–1004
Kargiotis O, Chetty C, Gogineni V et al (2008) uPA/uPAR down regulation inhibits radiation-induced migration, invasion and angiogenesis in IOMM-Lee meningioma cells and decreases tumor growth in vivo. Int J Oncol 33(5):937–947
Acknowledgments
R. Mahlberg, Neurochirurgische Universitätsklinik Bonn, provided expert technical support. We are indebted to B. Harzheim for help with the acquisition of clinical data.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kandenwein, J.A., Park-Simon, TW., Schramm, J. et al. uPA/PAI-1 expression and uPA promoter methylation in meningiomas. J Neurooncol 103, 533–539 (2011). https://doi.org/10.1007/s11060-010-0411-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11060-010-0411-6