
Abstract
Increased tree mortality associated with hotter-drought events has rarely been recorded in mature (>10 years old) commercial plantations. However, we report a recent (2021–2022 warm season) event of this kind that occurred across the Mesopotamian region of Argentina, South America. We aimed to elucidate the mortality timing, rate, and magnitude associated with environmental and stand characteristics at local scale to determine which conditions make these productive systems more vulnerable to extreme climatic events. Using Sentinel-2 satellite images and field-data we estimated the canopy mortality from November 2021 to October 2022, in Pinus spp afforestations (mainly P. taeda). We found that spatio-temporal heterogeneity in canopy mortality was associated with variation in soil and topographic characteristics, but not in stand age and size. We found that canopy mortality occurs at greater extent, earlier and faster in shallow soils (≤ 0.6 m), while the lowest mortality occurred at depressed and waterlogged areas. Intermediate levels of mortality were observed in deep soils (> 1 m), where relatively higher mortality occurred in areas with shallow water table signs, inhibiting deep root development. Our results suggest that sites allowing a deeper rooting system and/or where soil water availability is expected to be higher and long-lasting could represent areas with low-mortality risk for fast-growing pine plantations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
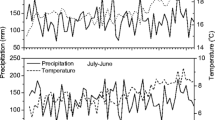
Increased tree mortality is a phenomenon widely reported over the last decades in different species and forests around the world (Allen et al. 2010; Senf et al. 2020). A recent study indicated that about 70% of the global drought-induced tree die-offs were related to the combined effects of drought and heat waves of short duration (i.e. “hotter-drought” events) mainly associated with global warming (Hammond et al. 2022). These hotter-drought events may induce rapid and abrupt tree mortality in drought-sensitive and (unexpectedly) drought-tolerant species in native forests (Kannenberg et al. 2021). Nevertheless, little evidence exists on the impact of these events on commercial tree species introduced outside their native range. Widespread tree mortality in commercial and/or managed plantations has been mainly associated with biotic agents or the combined effect of drought stress and pathogen attack (Černý et al. 2016; Holuša et al. 2018; Ji et al. 2020); but increased and small-scale mortality associated with hotter-drought have also been recently reported in Pinus spp. (Carnegie et al. 2022) and Eucalyptus spp. plantations (Pascual et al. 2022). In contrast, abrupt, simultaneous and regionally distributed tree mortality event of commercial forest species has been recently reported in the Mesopotamian region of Argentina, South America (Fig. S1), which affected about 25,000 ha of plantations out of a total of 300,000 ha planted in a subregion (SAGyP 2022a). This large-scale mortality event was associated with a hotter-drought period that occurred during the 2021–2022 warm season (Fig. 1), raising serious concern in the forestry sector.

(A) Daily accumulated precipitation in the study area. (B) Maximum daily temperature in the same periods as in A. Black lines correspond to the period of 7 years prior to the mortality event. Orange lines correspond to the year of study, starting in November 2021 when the first mortality signs were observed in some stands. Arrows indicate the dates in 2021 and 2022 of satellite images processed for this study. Lightgrey shadow indicates the warm spring-summer season in the study area
Field-based and remote sensing observations indicated that the observed mortality was spatially heterogeneous at regional and local scales (SAGyP 2022a). At the regional scale, spatial variation in tree mortality was mainly related to differences in species planted, being the most affected Pinus taeda, followed by P. elliottii var. elliottii, and to a lesser extent species of the genus Eucalytpus (SAGyP 2022a). Another important driver of spatial variability in tree mortality at the regional scale seems to have been the age of plantations, with higher tree mortality in mature (10 years old or more, close to the rotation period in the area) than in younger plantations. However, at a local scale in which age and species variability was generally low, stands presented variation in mortality proportion giving rise to the question of what did determine that some stands were highly affected and others were not even when located at short distances one from the other.
Landscape heterogeneity, determined by topography, aspect, soil texture and depth, could be a key driver of the observed variability in tree mortality at local scale. In this regard, both in native and plantations forests, landscape characteristics favouring the accumulation, percolation and promoting long-lasting soil water availability have enhanced tree water status (Nardini et al. 2021; Nosetto et al. 2005), reduced tree mortality (Tai et al. 2017) and canopy dieback (Schwantes et al. 2018). This evidence suggests that landscape characteristics (which can be resumed into the site quality concept in the forestry discipline) where tree stands are planted could be key to explaining and predicting stands´ vulnerability to hotter-drought events.
In this framework, we aimed to contribute to forest mortality assessment at global scale by reporting a large-scale event occurred in commercial pine stands in N.E. Argentina. Beyond the report, our objective was to evaluate the role of the site quality on the temporal trajectory of this unprecedented and abrupt mortality event recorded in managed plantations of Pinus taeda due to extreme hot drought conditions. We used high-resolution (10 m) maps of canopy mortality derived from Sentinel-2 satellite images developed for these forest stands (Gatica et al. 2023). From this map, we extracted values of the mortality proportion at stand level from different site quality areas and for different days after the mortality event started in November 2021. Specifically, our objectives were: (1) to evaluate whether site quality is relatively more important than other forest management characteristics (such as stand age and size) to explain spatial variation in mortality proportion, and (2) to compare temporal mortality trajectory among different site quality conditions.
Methods
Study site
Forest plantations studied were installed on natural grassland and floodplains (i.e. afforestation) in the northeast of Argentina (Corrientes province, Mesopotamia region; Fig. S1). The main tree forestry species in the area was Pinus taeda (loblolly pine), a non-native species from North America. The planted seedlings came from locally genetically improved materials well adapted to local conditions. The stands belong to a private company owning around 20,000 ha. of pine plantations in the study area. No detailed information on geographic coordinates can be publicly provided for confidentiality reasons, but they are available for scientific purposes on request to the authors. The climate is humid subtropical, with a long-term (30 years) average maximum temperature in summer (Oct–Mar) of 30.04 °C and an average minimum temperature in winter (Apr–Sep) of 12.70 °C. The mean annual precipitation averaged during the last 30 years is 1337.56 mm, concentrated (70%) in the austral warm season (Weather Station “EEA INTA Mercedes”; Latitude: -29.1958; Longitude: -58.0443).
In particular, this study area has experienced an anomalous heat event during the warm period between December 2021 and March 2022, where daily maximum temperature for 65 days exceeded by 2 to 10 °C the previous 7-year average daily maximum temperature for the same time period (Fig. 1B). In this period, also daily precipitation was significantly lower than daily records averaged over the past 7 years, accumulating 200 mm less than the expected (Fig. 1A). Long-term records of weather stations closer to the study area (\( {/sim}\) 90 km), indicate that drought events of equal magnitude were experienced in 1986 for this region (a single-year event; SIGA 2023). Therefore, the recent hotter-drought event represents the only anomalous climatic event experienced by the trees studied.
The soils are aquic Argiudols, with clay loam texture (SISINTA, accessed January 2022). The landscape is characterised by depressed plains largely occupied by floodplains, where flooded area changes over time depending on the dynamics of the Paraná and Miriñay rivers (Contreras et al. 2018).
Mortality estimation
The canopy mortality map was generated through Sentinel-2 satellite image classification methods using the Random Forest algorithm. We used a Sentinel-2 image from March 23, 2022, coinciding with the date of field data on the percentage of canopy mortality provided by the technicians of a forestry company. On this image and using the field data, we generated 283 (of 120 × 120 m in size) training and validation polygons with the objective of classifying the scene into classes of healthy (without mortality symptoms), partial canopy mortality (ranging from 20 to 60% of pixels with mortality symptoms) and total canopy mortality (> 80% of pixels with mortality symptoms) forests, as well as harvested forest and areas of non-forest (Fig. S2). For the classification process we used spectral bands and indices previously detected as appropriate to discriminate mortality levels in forests (see review in Huang et al. 2019). Before spectral indices calculation some bands were resampled and thus we homogenized the bands at a resolution of 10 m. The classification process included the training (using 70% of the 283 polygons) of random forest models and the selection of bands or indices using 10 k-fold spatial cross-validation strategy (Meyer et al. 2019). Validation was performed using the confusion matrix method based on the use of the remaining 30% of the training polygons.
The produced map showed validation metrics that suggest almost perfect scene classification (Overall accuracy = 0.97 [CI = 0.96–0.97] and Kappa Index = 0.97). All classes were identified with errors of commission and omission less than 10%, but the total canopy mortality class had errors less than 1% (Table S4). This high precision and accuracy to detect forest patches with total canopy mortality is related to the fact that forests with high mortality have a substantially and significantly different spectral signal from the other classes in the scene (see details in Gatica et al. 2023). Because total mortality can be detected with high precision, we have used this trained Random Forest model to predict total mortality for different scenes before and after March. In total we have obtained total mortality maps for 6 scenes covering the temporal range from November 2021 to October 2022 (indicated with arrows in Fig. 1B and in Fig.S3). It is important to highlight that we focus on total mortality not only because it has a high identification precision but also because we have quantified that patches with intermediate mortality reach total mortality on later dates in 95% of the cases (see maps F, G and H in Fig. S3). Harvested forest and non-forest areas were used as mask for enhance the total canopy mortality quantification.
To quantify the canopy mortality rate we estimated the number of pixels under total canopy mortality class into forest stands which correspond to the observational unit of the National Forestry Inventory of Argentina (SAGyP 2022b; see black-bordered boxes map in Fig. S4). The forest stands studied here (n = 616) have an average area of 4.59 ha (SD ± 0.91) and a range of minimum and maximum area of 0.01 and 10.96 ha, respectively (Table S1).
Forest management covariates
We characterised each forest stand with data from a site quality map based on a Site Index adapted by the forestry company (Table S2). Site quality for forestry activities was defined by an average performance of different forestry species at local scale, representing the highest suitability of each site for Pinus (sites P) and Eucalyptus (sites E) species, which depends on the mean annual increment (MAI) at harvest time. Also, this adapted Site Index is accompanied by a qualitative description of natural vegetation, soil type and depth, and waterlogging conditions (see Table S2). Briefly, site quality Rc, which represents the lowest productive sites, is characterised by shallow soils developed over a rocky impediment up to 50 cm depth, located in the highest areas of the landscape. Sites qualities Ba and P15 correspond to relatively low productive forestry situations located on floodplains (areas remaining waterlogged during the rainy seasons). Intermediate levels of forest productivity appear in site qualities P18, P21, and P25, which are characterised by soils with low permeability, slow runoff, and drainage, and with the presence of a textural clay horizon at about 50 cm depth. These site qualities correspond to areas of moderate elevation (flattened and irregular hills), with a hanging water table of variable depth (50 to 80 cm depth). Finally, the most productive sites correspond to the E35 category; these sites occur on elevated areas of the landscape but their soils are deeper (\( {/sim}\) 100 cm) than at Rc.
Site quality description was complemented by quantitative field measurements of soil characteristics and qualitative observation of rooting depth by means of soil excavation in three contrasting site quality levels (categories Rc, P21, and E35, Table S2) carried out in August 2022. At that moment, total (> 90% of trees died), partial (\( {/sim}\) 50% trees died), and null tree mortality was observed in Rc, E35, and P21 site qualities, respectively. Observed maximum depth of roots was around 50 cm in Rc site, corresponding to the rocky impediment, and more than 100 cm deep in P21, where the Bt horizon was at that depth or deepest. In the case of E35, roots were mostly shallow (\( {/sim}\) 30 cm), with variable maximum depth depending on the presence of anoxia signs in the soil due to a shallow water table.
It is worth noting that loblolly pine represents about 90% of the stands evaluated in the present study (Table S1), therefore the patterns observed mostly are driven by this species. Other species present in the study area were Eucalyptus spp, P. halepensis, P. elliottii and hybrids P. elliottii x caribaea (slash pines).
Data analyses
We evaluated the effect of site quality, stand age and size on the proportion of canopy mortality per forest stand using generalized linear mixed models (GLMM) (Zuur et al. 2009) and under multi-model selection inference approach (Burnham and Anderson 2002). We fitted a full model that had time (measured as days after November 19 2021 which coincide with the Sentinel 2 image available without canopy mortality symptoms), site quality (SQ), age, and stand size, and the interactions between time and site SQ, and time and age, as fixed factors. It must be noted that the stand age range differs substantially between site quality categories (Table S3). Hence, we have not included interaction terms between age and site quality because our experimental design was constrained for inference only related to additive effects. In addition, we added a quadratic term of time because we observed that the mortality rate reduces its linear increase after some time since November 19 (beginning of the analyzed period). Finally, a first-order autoregressive error structure (AR1) was included to take into account temporal correlation into forest stands (Zuur et al. 2009). We modelled canopy mortality proportion using a Binomial distribution with a logit link function (Zuur et al. 2009). From this full model, we generated “nested models” removing terms from higher-order to a null model (without covariates). For each model, we calculated the Bayesian Information Criterion (BIC) and the difference in BIC (ΔBIC) between each model and the best model (model with the lowest BIC value) (Burnham and Anderson 2002). Statistical significance of the best model was evaluated using the Analysis of Deviance based on Wald test (Bolker et al. 2009). The inclusion of Species as a random factor explained a small (< 1%) variation in mortality temporal trajectory, suggesting that the pattern observed is independent of Species planted.
Considering that site quality was the most important variable explaining spatiotemporal patterns in mortality proportion (see Results), we tested the impact of the site quality on the temporal evolution of mortality using a non-linear mixed model (NLMM) (Pinheiro and Bates 2000). In this case, we transformed canopy proportion mortality to percentage unit for easier interpretation. We considered fitting the temporal evolution of mortality using a Weibull model of the following form:
where k is the asymptote (representing maximum accumulated mortality); a is a parameter related to the rate of mortality; and b is a parameter related to the magnitude of the inflection point (high b values indicate abrupt increments in canopy mortality). Site quality was included as a predictor of the variation for each Weibull parameter (Pinheiro and Bates 2000). The significance of site quality effect was evaluated using the F-test statistic (Pinheiro and Bates 2000).
Data analyses were carried out using the computational language R version 4.2.2 (R Core Team 2022). GLMM was fitted using r-package glmmTMB (Brooks et al. 2017) and NLMM using r-package nlme (Pinheiro et al. 2023).
Results
The model selection approach showed that the model including time, site quality, and their interaction was the most parsimonious model (with the lowest BIC value, Table 1) and with the highest support from our data (highest relative Weight value, Table 1). This model indicates that canopy mortality proportion depended on the interaction between site quality and time (χ2 = 131.31, df = 12, p < 0.001; Table S4), showing that differences among site quality levels varied across days after Nov 19, 2021 (Fig. 2A). Canopy mortality proportion was usually the highest in Rc sites, particularly after day 121 (around March 19, 2022, almost the end of austral summer) (Fig. 2A). In contrast, mortality proportion was the lowest (near to zero) in both P15 and Ba site qualities, and this pattern was consistent across the whole studied period (Fig. 2A). Intermediate canopy mortality proportion was usually observed in sites P25, P18, P21, and E35 after day 186, but differences among these site quality levels alternated ranks depending on the time (Fig. 2A). Within these four site qualities, the mortality proportion in P18 was the lowest until August 16, 2022, but increased to higher values—similar to E35—towards the end of the study period (spring; Fig. 2A). In agreement with the mentioned patterns, the total area covered with Rc sites also showed the highest proportion of stands highly affected by canopy mortality, followed by P18, P21, P25, and E35; while in Ba and P15 areas, no stands with high mortality were recorded (Fig. 2B).

The temporal trajectory of canopy mortality proportion (estimated in %) (A) and of the number of forests stands exceeding 80% of pixels with high mortality (B) in different Site Qualities areas. The canopy mortality proportion was estimated from canopy mortality maps based on Sentinel-2 images and a Random forest model developed. Points in (A) represent the mean canopy mortality proportion at forest stand level and bars indicate standard deviation. Solid and dashed lines indicate the Weibull function expected by each Site Qualities derived from non-linear mixed models. The Weibull function could not be fitted to Site Qualities P15 and Ba
Complementary to the pattern detailed above, we found that the maximum accumulated mortality (K), the mortality rate (a) and the day at the inflection point (b) varied significantly among site qualities (all p values < 0.001; Table S5). The fastest mortality rate was recorded in Rc, followed by P21, P25, and E35 (Table 2; Fig. 2A); while the slowest mortality rate was observed in P18 (Table 2; Fig. 2A), which also showed the steepest increases in mortality towards the end of the period analysed (indicated by the highest b parameter; Table 2; Fig. 2A). A similar ranking was observed in the maximum accumulated mortality (parameter K, Table 2; Fig. 2A), but in this case, stands planted in P21 presented a lower accumulated value (Table 2; Fig. 2A).
Discussion
The empirical evidence reported in this study suggest that the Pinus taeda mortality event was triggered by an anomalous increase in maximum temperature and reduced rainfall during the 2021–2022 warm season (i.e. a hotter-drought event) (Fig. 1). This period also coincided with the last portion of three consecutive years of pronounced water stress in the whole study region due to La Niña phase of the ENSO phenomenon (NASA 2023). The registered maximum temperatures, with several consecutive days above 35 °C, are probably much higher than the optimal threshold for carbon gain of P. taeda, in the range of photosynthetic damage (Nedlo et al. 2009; Teskey and Will 1999). In addition, highly productive progenies planted in the area, as a product of breeding programs, are highly vulnerable to xylem cavitation (Bulfe and Fernández 2016; Bulfe et al. 2016), suggesting that a potential trade-off between productivity and resistance to extreme drought conditions has led to the observed phenomenon. Moreover, this study case was temporally coincident with other regional-distributed mortality observations during the same period (Fig. 1), where mortality as primarily triggered by biotic agents was discarded by field-based surveys (SAGyP 2022a). To the best of our knowledge large scale (at regional scale), simultaneous and abrupt increased tree die-off associated with hotter-drought events have not been reported in planted forests (but see evidence of non-simultaneous and/or small-scale events in Carnegie et al. 2022; Pascual et al. 2022).
As was expected, we found that canopy mortality and its temporal trajectory were spatially associated with the spatial variation in site quality, which indeed was defined a priori by the forestry company for operational purposes. Canopy mortality characterised by higher rate and maximum accumulation, as well as an earlier initiation, was observed in stands where soil water would probably be scarce and short-lasting (sites locally named “Rc’). It is important to highlight that canopy mortality detection was possible when around 80–90% of the trees presented canopy browning symptoms, which occur at the end of a sequence of processes (McDowell et al. 2008). In particular, progenies of loblolly pine cultivated in the study region present high vulnerability to xylem cavitation, with mean water potential at 50% loss of hydraulic conductivity—P50—around − 1.7 MPa (Bulfe et al. 2016) compared to mean P50 values of -2.6 MPa or -3.1 MPa in US progenies or genotypes (Aspinwall et al. 2011; Rosner et al. 2019). This relatively high vulnerability to xylem cavitation could be the cause of an early and/or a high degree of xylem embolism when soil water deficit and evaporative demand exceeded a critical threshold (Preisler et al. 2021; Kannenberg et al. 2021) inducing to generalised canopy browning. Moreover, even when climatic conditions ameliorated, mortality progressed during winter under normal conditions of temperature and precipitation, indicating that the physiological function could not be recovered. In contrast, we found that the lowest accumulated canopy mortality occurred in those stands located on topographically depressed and waterlogged landscape units (sites P15 and Ba). Under these conditions, the expected higher and long-lasting soil water availability could help the trees to tolerate high atmospheric demand and to minimise physiological damage during the hotter-drought periods, as proposed by the “hydraulic refugia” hypothesis (McDowell et al. 2019; Kannenberg et al. 2021).
Intermediate levels of canopy mortality were observed across stands located where soil water is expected not to be a limiting factor, at least during normal growing seasons (sites P18, P21, P25, and E35). In these sites, the canopy mortality was delayed and occurred at a slower rate than stands located in the most water-limited sites, possibly because the soil water remained available for a longer time. Notably, within intermediate-mortality levels, stands located at P21 site quality presented the lowest accumulated canopy mortality. Field observations there suggest deeper root development compared to trees in sites E35—which presented the highest relative mortality-, associated with the presence of hydromorphic signs in the upper soil layers in the latter situation. Relatively deeper roots, like those in the P21 sites, could potentially allow access to reliable and long-lasting sub-surface soil water sources during drought periods, increasing the chance of tree survival (Mackay et al. 2020). Hence, our results suggest that landscape-induced variations in soil water and intra-specific differences in rooting depth emerge as possible non-exclusive mechanisms (Chitra-Tarak et al. 2018; Nardini et al. 2016, 2021; Paz‐Kagan et al. 2017) explaining variations in canopy mortality trajectory observed in this study. However, more specific studies focused on the rooting systems should be performed to confirm these preliminary observations.
As in other regions worldwide, hotter-droughts events are expected to increase in intensity and frequency in the study region, which is the main forestry region supplying the wood industrial demands of Argentina. In this regard, although no changes in mean precipitation are predicted, high interannual variability in precipitation and an increase of 1.5 and 3.5 °C in mean temperature for 2039 and 2099 (SAyDS 2014), are expected. These projections suggest that hotter-drought events such as those observed recently could increase in frequency (Hammond et al. 2022) over the next decades in this region. This context can be exacerbated if we take into consideration the fact that a sustained expansion of forest plantations is developed into relatively marginal soils in many regions, mainly due to the competition with other land uses (such as protected native forests and agriculture). In the particular case of Corrientes province, afforestation expansion has occurred at the expense of a reduction of wetlands and has indirectly induced overgrazing in the remaining wetland areas (Pereira et al. 2023a). However, recent evidence suggests that afforestation expansion could be considered sustainable if it occurs on highly-degraded wetlands, leaving low-degraded wetlands as priority areas for their conservation (Pereira et al. 2023b). In this regard, a sustainable afforestation planning could involve a strategy of hierarchical nature. This planning could begin with the identification of degraded wetland areas and, based on our results, within these areas select those sites where soil water availability is expected to be higher and long-lasting, but at the same time allow for the development of an extended root system. These areas could present lower hot-drought associated risk at the landscape level.
Although the estimation of the socioeconomic impact of the mortality event is outside the objectives of our study, a gross estimation can be done to highlight the consequences of this kind of event on the local and regional forestry sector. At provincial level, considering only the areas affected with total canopy mortality (10,543 ha), the dead wood volume would represent around 2,100,000 m3 (i.e. about 200 m3 per ha of stem biomass at the average age of affected trees). This is more than the amount of wood that is allocated annually to the paper industry, the main destination of forestry production in the region (SAGyP, 2022). The high volume of dead wood surpassed the logistics available in the region to cut and transport the logs, so just a small fraction was cut on time. It is important to note that this number summed up to the whole-stands losses due to extended fires in the region that occurred previous and simultaneously with the reported tree mortality event. Fires affected around 31,000 ha of cultivated forests (6% of their area) and 12% of the total surface of Corrientes province, with wetlands as the most affected ecosystems (Saucedo et al. 2023). Moreover, total losses during the studied hot-drought event remain elusive because satellite image classification method cannot accurately estimate areas of partial canopy mortality, so the informed value is somewhat conservative. Besides the losses of huge volumes of wood with no value for the industry, the standing dead trees imply an additional cost, estimated by the forestry companies in U$S 250/300 per ha., to extract the trees and prepare the soils to reforest the area, cost that is added to the normal costs of plantation. As a consequence of the recent extended fires and drought-related mortality of the trees, the local forestry sector foresees that the offer-demand relationships will be largely altered at regional level, with still no clear socio-economic consequences.
Our study remarks the importance of soil characteristics and topography regulating the spatio-temporal forest mortality dynamics mediated by their role on soil water balance. This conclusion reinforces the idea discussed by Trugman et al. (2021) that understanding only the physiology or functional traits of the tree species that explain drought-vulnerability is not enough to accurately predict the drought-mortality in native and planted forests. Hence, from a forestry perspective site selection is a key step to minimise drought-induced mortality, which must be accompanied by academic and forestry sector efforts directed to disentangle whether the available genetic variability and plasticity of currently planted species is enough to adapt to the new climate or if it is necessary to change the species to be planted; and if it is possible to increase—at least to some extent—their adaptability by means of silvicultural management.
References
Allen CD, Macalady AK, Chenchouni H et al (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Aspinwall MJ, King JS, Domec J-C et al (2011) Genetic effects on transpiration, canopy conductance, stomatal sensitivity to vapour pressure deficit, and cavitation resistance in loblolly pine. Ecohydrology 4:168–182. https://doi.org/10.1002/eco.197
Bolker BM, Brooks ME, Clark CJ et al (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135. https://doi.org/10.1016/j.tree.2008.10.008
Brooks ME, Kristensen K, van Benthem KJ et al (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized Linear mixed modeling. R J 9:378. https://doi.org/10.32614/RJ-2017-066
Bulfe N, Faustino L, Pinazo M et al (2016) Bases fisiológicas del crecimiento diferencial en la fase inicial del desarrollo de dos progenies de Pinus taeda bajo condiciones de campo, en Misiones Argentina. Bosque (Valdivia) 37:273–284. https://doi.org/10.4067/S0717-92002016000200006
Bulfe N, Fernández ME (2016) Morpho-physiological response to drought of progenies of Pinus taeda L. contrasting in mean growth rate. New Forest 47:431–451. https://doi.org/10.1007/s11056-016-9524-x
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Carnegie AJ, Kathuria A, Nagel M et al (2022) Current and future risks of drought-induced mortality in Pinus radiata plantations in New South Wales, Australia. Aust for 85:161–177. https://doi.org/10.1080/00049158.2022.2145722
Chitra-Tarak R, Ruiz L, Dattaraja HS et al (2018) The roots of the drought: Hydrology and water uptake strategies mediate forest‐wide demographic response to precipitation. J Ecol 106:1495–1507. https://doi.org/10.1111/1365-2745.12925
Contreras FI, Contreras SA, Méndez CR et al (2018) El Paisaje De Lomadas arenosas desde El Pleistoceno superior a la actualidad. Boletín Geográfico 40:30–50
Černý K, Pešková V, Soukup F et al (2016) Gemmamyces bud blight of Picea pungens: a sudden disease outbreak in Central Europe. Plant Pathol 65:1267–1278. https://doi.org/10.1111/ppa.12513
Gatica G, Gyenge J, Fernández ME (2023) Detección De Niveles De mortalidad en plantaciones forestales del NE argentino usando una combinación de índices espectrales satelitales. In: Peri et al (eds) Actas Del VIII Congreso Forestal Latinoamericano Y V Congreso Forestal Argentino. CONFLAT, Mendoza, pp 27–30
Hammond WM, Williams AP, Abatzoglou JT et al (2022) Global field observations of tree die-off reveal hotter-drought fingerprint for Earth’s forests. Nat Commun 13:1761. https://doi.org/10.1038/s41467-022-29289-2
Holuša J, Lubojacký J, Čurn V et al (2018a) Combined effects of drought stress and Armillaria infection on tree mortality in Norway spruce plantations. For Ecol Manag 427:434–445. https://doi.org/10.1016/j.foreco.2018.01.031
Ji Y, Zhou G, Li Z et al (2020) Triggers of widespread dieback and mortality of poplar (Populus spp.) plantations across northern China. J Arid Environ 174:104076. https://doi.org/10.1016/j.jaridenv.2019.104076
Kannenberg SA, Driscoll AW, Malesky D, Anderegg WRL (2021) Rapid and surprising dieback of Utah juniper in the southwestern USA due to acute drought stress. For Ecol Manag 480:118639. https://doi.org/10.1016/j.foreco.2020.118639
Mackay DS, Savoy PR, Grossiord C et al (2020) Conifers depend on established roots during drought: results from a coupled model of carbon allocation and hydraulics. New Phytol 225:679–692. https://doi.org/10.1111/nph.16043
McDowell N, Grossiord C, Adams HD et al (2019) Mechanisms of a coniferous woodland persistence under drought and heat. Environ Res Lett 14:045014. https://doi.org/10.1088/1748-9326/ab0921
Meyer H, Reudenbach C, Wöllauer S, et al (2019) Importance of spatial predictor variable selection in machine learning applications – Moving from data reproduction to spatial prediction. Ecol Model 411:108815. https://doi.org/10.1016/j.ecolmodel.2019.108815
Nardini A, Casolo V, Dal Borgo A et al (2016) Rooting depth, water relations and non-structural carbohydrate dynamics in three woody angiosperms differentially affected by an extreme summer drought: rooting depth and plant hydraulics. Plant Cell Environ 39:618–627. https://doi.org/10.1111/pce.12646
Nardini A, Petruzzellis F, Marusig D et al (2021) Water ‘on the rocks’: a summer drink for thirsty trees? New Phytol 229:199–212. https://doi.org/10.1111/nph.16859
NASA (2023) El Niño-Southern Oscillation (ENSO) Index. Jet Propultion Laboratory, https://sealevel.jpl.nasa.gov/overlay-elnino/. Accessed 10 April 2023
Nedlo JE, Martin TA, Vose JM, Teskey RO (2009) Growing season temperatures limit growth of loblolly pine (Pinus taeda L.) seedlings across a wide geographic transect. Trees 23:751–759. https://doi.org/10.1007/s00468-009-0317-0
Nosetto MD, Jobbagy EG, Paruelo JM (2005) Land-use change and water losses: the case of grassland afforestation across a soil textural gradient in central Argentina. Global Change Biol 11:1101–1117. https://doi.org/10.1111/j.1365-2486.2005.00975.x
Pascual A, Tupinambá-Simões F, Guerra-Hernández J, Bravo F (2022) High-resolution planet satellite imagery and multi-temporal surveys to predict risk of tree mortality in tropical eucalypt forestry. J Environ Manage 310:114804. https://doi.org/10.1016/j.jenvman.2022.114804
Paz-Kagan T, Brodrick PG, Vaughn NR et al (2017) What mediates tree mortality during drought in the southern Sierra Nevada? Ecol Appl 27:2443–2457. https://doi.org/10.1002/eap.1620
Pereira LF, Kurtz DB, Fernandes-Filho EI (2023b) Forestry feedbacks in Malezales’ degradation: insights from past to plan future policy-driven forestry expansion over wetlands in Argentina. J Nat Conserv 76:126504. https://doi.org/10.1016/j.jnc.2023.126504
Pereira LF, Mendes Oliveira RR, Kurtz DB et al (2023a) It is possible to conciliate policy-driven forestry expansion and malezales wetlands conservation in Argentina? Environ Sci Policy 142:153–163. https://doi.org/10.1016/j.envsci.2023.02.011
Pinheiro JC, Bates D (2000) Mixed-effects models in S and S-Plus. Springer Verlag, New York, Inc.
Pinheiro JC, Bates D, R Core Team (2023) Nlme: Linear and nonlinear mixed effects models. R Package Version 3:1–162
Preisler Y, Tatarinov F, Grünzweig JM, Yakir D (2021) Seeking the point of no return in the sequence of events leading to mortality of mature trees. Plant Cell Environ 44:1315–1328. https://doi.org/10.1111/pce.13942
McDowell N, Pockman WT, Allen CD et al (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178:719–739. https://doi.org/10.1111/j.1469-8137.2008.02436.x
R Core Team (2022) R: A language and environment for statistical computing
Rosner S, Johnson DM, Voggeneder K, Domec J-C (2019) The conifer-curve: fast prediction of hydraulic conductivity loss and vulnerability to cavitation. Ann for Sci 76:1–15. https://doi.org/10.1007/s13595-019-0868-1
SAGyP (2022a) Análisis climático y productivo en relación a la mortalidad de Pinus taeda en la provincia de corrientes. Secretaría De Agricultura, Ganadería Y Pesca, Ministerio De Economía De Argentina. INTA-CONICET y Secretaría de Desarrollo Foresto-industria de la provincia de Corrientes, Corrientes, Argentina
SAGyP (2022b) Mapa de Plantaciones Forestales de Actualización Permanente. Dirección Nacional de Desarrollo Foresto Industrial, Secretaría de Agricultura Ganadería y Pesca de la Nación, Ministerio de Economía. https://visor-geoforestal.sigforestal.repl.co/. Accessed 21 March 2023
Saucedo G, Perucca AR, Kurtz DB (2023) Las Causas De Los incendios de principios del año 2022 en la provincia de corrientes. Ecología Austral 33:273–284
SAyDS (2014) Tercera Comunicación Nacional Sobre Cambio Climático. Cambio Climático en Argentina; Tendencias Y Proyecciones. Secretaría de Ambiente y Desarrollo Sustentable de la Nación, Buenos Aires, Argentina
Schwantes AM, Parolari AJ, Swenson JJ et al (2018) Accounting for landscape heterogeneity improves spatial predictions of tree vulnerability to drought. New Phytol 220:132–146. https://doi.org/10.1111/nph.15274
Senf C, Buras A, Zang CS et al (2020) Excess forest mortality is consistently linked to drought across Europe. Nat Commun 11:6200. https://doi.org/10.1038/s41467-020-19924-1
Tai X, Mackay DS, Anderegg WRL et al (2017) Plant hydraulics improves and topography mediates prediction of aspen mortality in southwestern USA. New Phytol 213:113–127. https://doi.org/10.1111/nph.14098
Teskey RO, Will RE (1999) Acclimation of loblolly pine (Pinus taeda) seedlings to high temperatures. Tree Physiol 19:519–525. https://doi.org/10.1093/treephys/19.8.519
Trugman AT, Anderegg LDL, Anderegg WRL et al (2021) Why is Tree Drought Mortality so hard to Predict? Trends Ecol Evol 36:520–532. https://doi.org/10.1016/j.tree.2021.02.001
Zuur AF, Ieno EN, Walker NJ et al (2009) Mixed effects models and extensions in ecology with R. Springer Science + Business Media, LLC 2009, New York, NY
Acknowledgements
The authors thank the technical and professional personnel of the private forestry company who collaborated in the field work and in the provision of forest management information. This work was supported by the Agencia Nacional de Promoción Científica y Tecnológica de Argentina, MinCyT under Grant PICT 2018–02510 Préstamo BID and Grant PICT-2021-I-INVI-00709 Préstamo BID. This research was developed in the framework of the International Associated Lab Forestia INRAE-INTA (France, Argentina). This manuscript arises from the interaction between researchers from CONICET and INTA (Argentina), in conjunction with policymakers and professionals from the forestry industry of the Mesopotamian region.
Author information
Authors and Affiliations
Contributions
G.G., J.G., and M.E.F. developed the methodology for mortality estimation and wrote the first version of the main manuscript text. G.G. processed and analyzed data. N.B. and N.P. provided the field data. F.D.T. provided forestry management information. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
No conflicts of interest are declared.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Gatica, G., Gyenge, J.E., Bulfe, N.M.L. et al. Unprecedented mortality induced by extreme hot-drought in commercial planted Pinus spp stands is locally modulated by soil characteristics. New Forests 55, 1333–1347 (2024). https://doi.org/10.1007/s11056-024-10037-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-024-10037-x