
Abstract
Radial growth was examined in two Scots pine stands that were seeded during the 1920s and 1930s due to reforestation and afforestation activity on the timberline of northern Finnish Lapland. Tree-rings of seeded pines were calibrated against the instrumental records of local weather and large-scale atmospheric patterns and further compared to pines of natural origin on the timberline. The studied stands were shown to contain common growth variability that differed from the variability of natural pines. Deviating growth of seeded pines was attributed to their only moderate growth dependence on mid-summer (July) temperatures and, likewise, their strong dependence on the autumn climate in the previous year, and linked to the different genetic origin of artificial stands due to southern seeds. Stands were thinned for different densities in 1985 and 1986. The growth response to thinning was markedly better than could be expected according to previous studies. We found that the positive growth reaction conceivably benefited from the ameliorated winter conditions, expressed as warmed (March) temperatures and the prolonged positive phase of the Arctic Oscillation (February). The results emphasize the determining influence of climatic fluctuations on reforestation and afforestation near the distributional limits of tree species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Annual radial increments of a tree stem, tree-rings, are commonly used indicators of biotic and abiotic factors affecting tree growth (Mikola 1950; Fritts 1976; Vaganov et al. 2006). The great advantages of tree-rings, and especially ring-widths, are that they are obtainable at low cost and form coherent and annually resolvable time series that provide a long-term perspective on variability of individual-to-regional tree growth. Tree-ring variability shows the influence from age-size related factors, competition and growth resources (Mikola 1950; Fritts et al. 1969; Fritts 1976; Spiecker 2002; Vaganov et al. 2006). Moreover, tree-rings can be directly calibrated against climatic records (Fritts et al. 1971; Fritts 1976; Lindholm 1996).
In northern Finland, the tree-ring growth of Scots pine is known to be limited by summer temperature (Hustich and Elfving 1944; Mikola 1950, 1952, 1962; Sirén 1961; Hari and Sirén 1972; Pohtila 1980; Lindholm 1996; Lindholm et al. 2000; Helama et al. 2002, 2005a; Macias et al. 2004). In addition, the natural variability in climate over North-West Europe that may influence the pines is often described using either the index of the North Atlantic Oscillation (NAO) or Arctic Oscillation (AO) (also called Northern Annular Mode) (Hurrell 1995; Thompson and Wallace 1998, 2000; Hurrell et al. 2001). This variability is characterized by an oscillation of atmospheric masses producing large-scale changes in the mean wind speed and direction over the North Atlantic. The AO is characterized by an alternation of atmospheric mass between the Arctic and middle latitudes, and it accounts for a significantly larger fraction of the variance in Northern Hemisphere surface air temperatures than the NAO (Thompson and Wallace 1998, 2000). The NAO was previously shown to influence the tree-ring growth of the pines by controlling the local climate variability in northern Finland (Macias et al. 2004).
The timberline in Finnish Lapland is often a broad, transitional zone from forest to isolated trees, being largely a polar timberline (Veijola 1998). Scots pine (Pinus sylvestris L.) forms the coniferous timberline in the region (Hustich 1966). These pines represent some of the northernmost occurrences of the species worldwide (Mirov 1967). The two stands studied here were artificially regenerated. Seeding occurred during the earlier half of the 20th century. Initiation of this activity was related to the concern at that time that ongoing cuttings at the timberline would result in the slow death of the northernmost pine forests in Finland (Renvall 1912). Subsequent to these concerns, the natural timberline regeneration has, however, taken place in the region (Hustich 1940, 1948; Sirén 1998; Juntunen et al. 2002) and, considering the harsh climatic conditions, the artificially regenerated pine stands investigated here have performed relatively well. The stands were thinned in the middle of the 1980s. According to Varmola et al. (2004b), the thinning reaction of the pines was markedly stronger and occurred with a considerably shorter lag than could have been expected based on previous experiments in northern Finland (Ruha and Varmola 1997). While the diameter growth response in naturally regenerated stands has not been clearly observed earlier than 10 years after the thinning in northern Finland (Ruha and Varmola 1997), the artificially regenerated pines reacted to increase in growing space in 2–3 years at the coniferous timberline (Varmola et al. 2004b). Moreover, pines responded to thinning systematically according to remaining stand density (Varmola et al. 2004b). Accordingly, one could hypothesise that the climatic conditions may have been especially favourable during the critical periods or that the growth of these pines could be related to climate differently than expected according to literature on natural pines (Hustich and Elfving 1944; Mikola 1950, 1952, 1962; Sirén 1961; Hari and Sirén 1972; Pohtila 1980; Lindholm 1996; Lindholm et al. 2000; Helama et al. 2002, 2005a; Macias et al. 2004).
Primary aim of the study was to test this hypothesis and an assemblage of sub-targets was set to examine the growth variability of the two pine stands in more details by dendroclimatic analyses. Firstly, the growth of seeded pines was compared to growth of natural pines. Further, the dendroclimatic analyses of this study aimed to reveal the potential climatic influences behind the pine growth variability. Previously Varmola et al. (2004b) demonstrated the variations in radial increment within the two stands, with tentative climate correlation to annual temperature sum. Our aim was to perform a systematic growth-climate analysis of seeded and thinned pines using local climate variables and indices of NAO and AO. Moreover, we aimed to unveil the long-term (decadal to centennial) climatic frame in which the growth of these pines had occurred. In so doing, we wanted to show the usefulness of dendroclimatic analyses for forest research, and demonstrate the importance of long-term climatic variations on regeneration and growth especially in the extremely harsh conditions near the distributional limits of trees.
Material and methods
Seeded and thinned stands
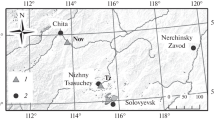
Two experimental stands of Scots pine along the coniferous timberline were studied (Fig. 1). Both belong to the areas that have been reforested and afforested in the protection forests of northernmost Finnish Lapland since the 1910s (Pohtila and Timonen 1980). Stands are located in Pakanajoki (69°33′39′′ N; 29°10′56′′ E; 90 m a.s.l.) and Mierasrova (69°31′33′′ N; 27°13′00′′ E; 190 m a.s.l) and are denoted hereafter as PAK and MIE, respectively. Soil in both stands represents dry forest land with an organic layer of a couple of centimetres only. Both sites were first prescribed burned. Broadcast seeding was carried out on the snow cover in spring 1928 in PAK whereas MIE was band-seeded in 1939. The pre-thinning stand densities were 1669 and 1972 stems per ha in PAK and MIE, respectively, and the thinning occurred in the spring of 1985 and 1986. Prior to that date, both stands were slightly thinned in 1952 (PAK), 1955 (MIE) and 1957 (PAK). The thinning treatments were carried out in PAK and MIE in 40 × 40 m square plots for the densities of 300, 550, 800, 1,050 and 1,300 stems per ha, denoted hereafter as PAK03, PAK05, PAK08, PAK10, PAK13, MIE03, MIE05, MIE08, MIE10 and MIE13, respectively. In addition, one treatment in both stands was left unthinned. The ring-widths within the plots of these treatments are denoted hereafter as PAK99 and MIE99. Each thinning was replicated in four plots in both of the stands. Seeds used in PAK originated 300 km south of the study site (Fig. 1), the origin of MIE seeds was unknown but was likely of southern origin.

Map showing the study region in northern Finland with squares indicating the weather station of Karasjok (K) and the seeded stands in Mierasrova (M) and Pakanajoki (P). An oval indicates the approximate geographical origin of the seeds. The naturally regenerated timberline pines originated from Kultima (1), Pitkäjärvi (2), Kalmankaltio (3), Luttojoki (4) and Kessi (5)
Dendrochronological work
An increment borer was used to extract cores for tree-ring analysis at breast height. The ring-widths were measured from each core to the nearest one hundredth of a millimetre. Ring-widths were cross-dated using dendrochronological approach of Holmes (1983) along with visual comparison of the produced time series. Dendrochronological cross-synchronization of series by their ring-width characteristics, narrow and wide rings, produces absolute dating of each sample to an accuracy of one calendar year (Fritts 1976). An average number of 92 and 80 trees per treatment were sampled in PAK and MIE. In all, collections of ring-widths in PAK and MIE resulted in 27035 and 18848 measured and cross-dated individual ring-width values from 551 and 480 trees, respectively.
Naturally regenerated pines
Control chronology contained Scots pine tree-ring series from five natural-grown sites on the timberline of the same region: Kalmankaltio (68°26′ N; 24°42′ E), Kessi (69°05′ N; 28°50′ E), Kultima (68°25′ N; 22°53′ E), Luttojoki (68°29′ N; 28°05′ E) and Pitkäjärvi (68°18′ N; 24°E). Control chronology was constructed as a sub-sample of particularly young pines (having first ring 1915 or later) that were extracted from much larger tree-ring network of naturally regenerated Scots pines on the timberline. Due to natural timberline conditions, these pines were regenerated sporadically during the periods of favourable climatic conditions and the stands can be characterized by a mixed age structure. This network was previously described and documented by Macias et al. (2004), Varmola et al. (2004a) and Timonen (2005). Similarly to seeded and thinned stands, the soils in control stands represent dry forest land with a thin organic layer. The initial tree-ring material of these sites was collected and cross-dated (Holmes 1983) similarly to the material from experimental stands.
Dendrochonological analysis
Series of tree-ring widths are known to contain a trend that is a non-stationary process partly arising from the geometric constraint of adding an annual volume of biomass to a stem of a tree with increasing radius (Cook 1985, 1990). The growth trend of many species ideally exhibits an abrupt and short lived increase in annual growth from the pith to a juvenile growth maximum, after which exponential decrease occurs as a function of cambial age until the outermost ring (Matalas 1962; Fritts 1976). This is also known to be the case for Scots pine in northern Finland (Mikola 1950; Cook et al. 1995; Helama et al. 2002, 2004b, 2005a, b). In order to remove this non-climatic trend from the initial ring-width series, a single growth curve was used to detrend all the individual ring-width series. Using the formula of modified negative exponential function (Fritts et al. 1969), the relationship between tree age and radial growth can be expressed as
where y represents the expected growth value (in mm) according to cambial age x. This curve was empirically parameterized by Helama et al. (2005b) using a dataset of Scots pine tree-rings that was thought to represent non-climatic tree ageing on the northern timberline in Finland for the past centuries. In so doing, the curve (Eq. 1) takes the form
Initial ring-widths were transformed into dimensionless growth indices as ratios, that is, the observed ring-width values were divided by expected values of the curve (Eq. 2). This procedure follows the regional curve standardization that was described by Briffa et al. (1992). Resulting annual ring-width indices were then averaged according to their calendar year into 12 different chronologies each representing different thinning treatments in PAK (six chronologies) and in MIE (six chronologies).
PAK and MIE growth indices were compared to the control chronology. Ring-width series of this chronology were detrended using the same model (Eq. 2) that was used to remove the growth trend from PAK and MIE ring-width series. Moreover, in order to separate the impacts of climate and thinning reaction on ring-widths, the annual index values of PAK and MIE chronologies were divided by the corresponding indices of the control chronology. These ratios between the thinning treatment chronology and the control chronology were hypothesized to reflect the growth release due to thinning with the minimized climate-driven influence (Haapanen et al. 1979).
Finally, the initial ring-width series were realigned according to their cambial ages and the mean curve for tree ageing was computed as an average of all realigned series in unthinned plots, separately for PAK99 and MIE99. Thus attained trends were further determined using the curvilinear function (Eq. 1) and the modelled growth trends of PAK and MIE were compared to predetermined regional ageing model of pine (Eq. 2).
Dendroclimatological analysis
Prior to climatic comparison, the growth indices were prewhitened using Box and Jenkins (1970) methods of autoregressive and moving average time series modelling (Cook 1985; Guiot 1986; Monserud 1986). The order of the autoregressive-moving average process was determined using Akaike (1974) Information Criteria. Prewhitening transforms autocorrelated series into a series of independent observations. These indices were used here to compute relationships between climate and tree-rings.
Identification of climatic features that affected tree radial growth was achieved using the bootstrapped response functions in a form of principal component regression that was designed to account for the commonly occurring collinearity of the monthly climatic variables. Each bootstrap estimate was obtained by generating 1,000 samples (Biondi 1997; Biondi and Waikul 2004). Ring-width variability was compared to monthly mean temperatures and precipitation sums in the adjacent weather station of Karasjok (69°28′ N; 25°31′ E; 129 m a.s.l) (Fig. 1) that provided high quality records with centennial temporal coverage representing climatic variability similar to study sites. Monthly indices of North Atlantic Oscillation (NAO) and Arctic Oscillation (AO) were included in the bootstrapped correlation analysis (Biondi and Waikul 2004) due to their correlation with local climate (Fig. 2). The NAO indices of Hurrell (1995) are found from the difference of normalized sea level pressure (SLP) between Ponta Delgada, Azores and Stykkisholmur/Reykjavik, Iceland. The AO indices of Thompson and Wallace (2000), identifying AO as the dominant empirical orthogonal function of sea-level pressure variability for the Northern Hemisphere, are found by projecting the AO pattern onto sea-level pressure anomalies.

Correlations (1951–1998) between temperature and precipitation (Karasjok weather station), and North Atlantic Oscillation (NAO; Hurrell 1995) (gray histograms) and Arctic Oscillation (AO; Thompson and Wallace 2000) (white histograms) over winter (DJF, December–January–February), spring (MAM, March–April–May), summer (JJA, June–July–August) and autumn (SON, September–October–November)
In addition to monthly response functions, the growth variability was explained by seasonal tri-monthly temperature and precipitation variables using linear regression. The higher the coefficient of the determination (R 2) in the regression, the more the climatic factors explained the variance in the ring widths.
The calibration period for response functions was the interval between 1945 and 1984. Due to the low number of ring-width series in MIE prior to the 1960s, climate-growth relationships were derived using the chronology of PAK only.
Results
Pre-thinning growth variability was similar in both stands (Fig. 3a, b). Post-thinning release interrupted the slight growth decline that had been in evidence since the middle of the 1970s (Fig. 3a, b). Trees responded to thinning systematically according to remaining stand density. Lag in response was 2–3 years.

Tree radial growth expressed as growth indices in PAK and MIE stands, shown separately for thinning densities of 300, 550, 800, 1,050 and 1,300 stems per ha and for unthinned plots (gray area emphasizes the post-thinning period). Pines in differently thinned plots responded to the thinning in proportional fashion, so that the more intense the thinning was, the more amplified was the growth release afterwards (a, b). Also shown are the ratios between the thinned and control tree-ring indices (c, d). The two lowermost plots (e, f) compared the ageing growth trends in PAK99 and MIE99 (unthinned) and in control dataset (Helama et al. 2005b)
PAK and MIE growth was compared to control chronology as ratios (Fig. 3c, d). The growth in PAK approximated the growth of the control chronology until the middle of the 1970s (Fig. 3c). Thereafter, relative decline occurred (ratios below one), similar to that already seen in the ring-width indices (Fig. 3a). This decline lasted until the end of the records in unthinned plots; only thinning was able to interrupt this decline. In MIE (Fig. 3d), the decline was very similar to that in PAK. However, as the overall growth level was higher in MIE, this decline did not result in such a poor growth as in PAK. The post-thinning radial growth (1989–1998) in PAK05 and PAK08 corresponded to the growth during the same period in the control chronology (Fig. 3c). In the case of the MIE stand, where the overall growth was better already prior to thinning, growth comparable to the control chronology was attained with slighter thinning, in MIE13 (Fig. 3d). In the two stands, the post-thinning tree-ring widths (1989–1998) were 154% (PAK03) and 184% (MIE03) of the width expected due to tree age; without thinning, the tree-rings were markedly narrower than expected by tree age, 69% of the expected growth in PAK99 and 79% in MIE99 (Fig. 3c, d).
The ageing growth trends for PAK and MIE were compared to theoretical pine ageing curve in the region (Eq. 2). The juvenile growth of PAK99 and MIE99 was markedly higher than could be expected according to the curve (Fig. 3e, f). This relationship was reversed as the pines got older, from a cambial age of 30 years, as counted from tree-ring series, upwards (Fig. 3e, f). Since the trees were sampled at breast height, they were actually around 20 years older than this and the corresponding point thus occurred at a cambial age of c. 50 years.
Markedly high correlation between the two chronologies, PAK and MIE (Table 1), implied that the climatic growth component was probably very similar in both of them. Climatic impacts on growth were interpreted in detail using the response functions (Fig. 4). Six out of 24 monthly temperature and precipitation variables were found to have a statistically significant influence on pine radial growth during the calibration period (1945–1984), only one of these six being negatively correlated with growth. All three significant temperature variables were positively correlated with growth including the influence of mean temperatures of the previous October, previous March and current July. Precipitation sums that influenced tree growth positively were the previous January and May. November precipitation of previous autumn was negatively related to pine growth. Moreover, the pine growth could be statistically related to NAO indices of the previous November and concurrent June and to AO indices of February. All these monthly indices had positive influence on growth. These responses could be used to assess the thinning response in the context of contemporary climate. Accordingly, it was found that the thinning response may have benefited from the prolonged positive phase of February AO and March temperatures (Fig. 5). Moreover, none of the 9 monthly variables showed markedly negative phase over the post-thinning years (Fig. 5) and it thus seemed likely that the climate was in general favourable to thinning release.

Relationships between the tree-ring indices and monthly meteorological variables of previous (small letters) and concurrent year (capital letters) to growth. Significant relationships are indicated as filled histograms

Local climate fluctuations over the pre- and post-thinng (gray area) periods exemplified with those nine meteorological variables that were shown to have significant influence on growth (Fig. 4). Thin and thick lines indicate the year-to-year and decadal variability in climatic features, respectively, about the long-term mean (horizontal line)
While pre-thinning radial growth in PAK and MIE closely resembled each other, the radial growth was evidently more dissimilar in the control chronology (Table 1; Fig. 6a). To summarise, the climate of the previous autumn had great influence on the growth in the seeded stands, whereas a similar relationship was absent from control chronology (Table 2).

Tree radial growth as tree-ring indices in PAK99 and MIE99 as well as in the control sites since 1915 (a). The 20th century pine growth has been anomalously good in the context of past centuries (the data for long chronology comes from Helama et al. 2005b) (b)
Discussion
Previous observations had shown that the seeded and thinned timberline pine stands had grown surprisingly well (Varmola et al. 2004b). The growth success of timberline Scots pine is depending on climate probably more than pine growth in any other environment (Mikola 1950; Ilvessalo 1970; Lindholm et al. 2000; Helama et al. 2005a). Accordingly, we hypothesised that either the climatic conditions had been especially favourable during the critical growth periods or that the growth of the seeded pines could be related to climate variability differently than the growth of naturally regenerated pines. A set of analyses was performed in order to compare the growth variability of these pines to growth of naturally regenerated pines and to local and large-scale climate variations. Hereafter, dendrochronological and dendroclimatolgocial results are discussed in the context of previously published knowledge of timberline pine growth.
Thermal climate at the coniferous timberline
The harshness of the local climate could be best viewed in the context of the growing season temperatures. According to Norokorpi (1994), the polar timberline of conifers in Finland can be generally expressed as a temperature sum of 600 degree-days and the tree-limit by 550 degree-days. According to the logistic regression model of Mikkola and Virtanen (2006), the timberline can be expected to have a temperature sum of 591 degree-days (R 2 = 0.44) and a mean July temperature of 12.2°C (R 2 = 0.47). Mean temperature sums for PAK and MIE were as low as 667 and 618 degree-days, respectively, with a range of 443–979 and 404–930 degree-days between the years 1961 and 1990 (Varmola et al. 2004b). While the inter-annual variability in thermal climate is ample, the seeded and thinned stands are both located in phytogeographical positions where the climatic conditions have recently remained only slightly milder than the theoretical timberline climate.
Pine growth and local climate
Similarly to our study, the positive influence of mean temperatures of the previous October on pine growth has been found in some of the high latitude studies in Finland, Norway and Sweden (Lindholm 1996; Kirchhefer 1999; Linderholm 2002). According to Lindholm (1996), this response was common to a variety of sites in the northern timberline region in Finland. The effect of mean temperatures in March, which was detected as a positive and significant growth controlling factor, appeared only in some sites or over limited time intervals according to previous studies in Norway and Sweden (e.g. Linderholm 2002; Kirchhefer 2000). According to Kirchhefer (2000), this response was related to heat advection by moist south-western air masses during this particular time of the year, under the threat of winter-desiccation, especially at south-facing slopes.
The prominent positive influence of the mean temperatures in July on Scots pine ring-widths in northern Finland, Norway and Sweden has been described by several authors (Erlandsson 1936; Hustich and Elfving 1944; Mikola 1950; Schove 1950; Sirén 1961; Lindholm 1996; Briffa et al. 1988; Kirchhefer 1999, 2000, 2001; Helama et al. 2004a; Macias et al. 2004). In general, the temporal variability in the growing season temperatures largely determines the growth variability of the trees at high altitudes and latitudes. The dependency of growth on summer temperatures and its connection to the seasonal stage of development was described for Scots pine in northernmost Finland in fine detail by Hari and Sirén (1972). According to Helama (2004a), the relationship between July mean temperatures and pine ring-widths can be proved to be time-independent over the past 300 years, whereas the relationships between June and August temperatures and growth is clearly time-dependent in northernmost Finland. Although July temperatures influenced the growth of seeded pines significantly and positively (Fig. 4), the strength of this influence was weaker than previously shown to be the case elsewhere in the region and in adjacent areas. This observation may, at least partly, explain the low correlation between the control chronology and studied stands (Table 1).
Among the precipitation sums that significantly influenced tree growth (Fig. 4), the precipitation in May is well described over several sites in the region and also in the vicinity (Lindholm 1996; Kirchhefer 2001; Macias et al. 2004), but also across the country, from northern to southern Finland (Lindholm et al. 2000; Helama et al. 2005a). According to Kirchhefer (2001), this response can be associated with the inducing effect of rainfall on snow-melt and soil warming at that time of the year. Positive influence of the precipitation sum in January was also found by Lindholm (1996) to be apparent in some sites in northernmost Finland, and by Kirchhefer (1999) in northern Norway. As parallel evidence, it is known that very thin snow cover in combination with low temperatures may cause damage to pines, especially in northern Finland (Jalkanen 1993). As a matter of fact, the same processes may interact behind the positive influence of February AO (Fig. 4).
In general, the climate of the previous autumn had great influence on the growth in the seeded stands, whereas similarly strong relationship was absent from control chronology (Table 2). Interestingly, the overriding dependence of growth variability on climate of the previous autumn has not been statistically described in previous studies in the region or adjacent areas, but could potentially be related to the hardening processes of the trees to the dormancy. Along with the relative weak response of seeded pines to mid-summer (July) temperatures, the results may emphasize the potential difference in the genetic origin of the two tree groups, that is, northern and southern seeds of the naturally and artificially regenerated pines, respectively. Genetic differentiation is indeed imprinted in the tree-ring growth of Scots pine: seeded under identical climatic conditions, the pines of southern origin show a later cessation of ring growth and transition to dormancy, and likewise a higher sensitivity to late growing season climate (Savva et al. 2001, 2003, 2006). This pattern could explain the difference in the climatic growth response between the artificially (seeds of southern origin) and naturally (seed of local northern origin) regenerated pines.
North Atlantic Oscillation and Arctic Oscillation
The use of NAO and AO indices as predictors of tree-ring variability was decided due to the fact that these phenomena seem to have significant influence over the local weather (Hurrell 1995; Uvo and Berndtsson 2002) (Fig. 2). Actually already Brier (1948) and Pohtila (1980) pointed to the importance of hemispheric surface pressure pattern and zonal circulation for the growth of northern Finnish pine tree-rings.
Negative growth influence of the precipitation sum of the previous November could be associated with the positive growth response to NAO of the same month (Fig. 4): correlation between November precipitation and November NAO was −0.48; and the correlations between tree-rings and November NAO indices, and tree-rings and November precipitation sums were 0.32 and −0.45, respectively. It seems that November NAO does not explain this relationship better than November precipitation, but the significant correlation between tree-ring and NAO indices is merely a reflection of NAO’s influence on the local precipitation. This seems not to be the case for the positive and significant correlation between June NAO and tree-ring indices (Fig. 4). This is a relationship that has been found in some of timberline sites in Finland, Norway and Sweden (Linderholm 2002; Macias et al. 2004; Helama et al. 2005b). However, neither June mean temperature nor precipitation sum separately influence the growth significantly. It could be even proposed that June NAO may actually describe the response to climate over a broader time-window and represents a transition from the snow-melt to the mid-summer warmth, bearing the features of both of these two important growth factors.
Post-thinning pine growth and climate
As already noted by Varmola et al. (2004b) and exemplified here (Fig. 3), the thinning reaction was markedly better than could have been expected according to the previous experiments in northern Finland (e.g. Ruha and Varmola 1997). The reaction was especially notable considering the harshness of the growing habitats and the greatly limited growth resources. Post-thinning growth was systematically better, the more intense the thinning. In addition, the growth ameliorated within only 2–3 years in PAK and MIE (Fig. 3), whereas the previous studies have documented a delay of 10 years for thinning growth reaction (Ruha and Varmola 1997). Yet, previous study from Finland had shown that weakness of the thinning reaction could be associated to dryness of the site (Hynynen and Arola 1999) by contrast, the PAK and MIE both represent dry forest soil. Moreover, the studied stands are situated at the timberline, whereas the previous studies focused on more southern pines. The better than expected growth response to thinning could be viewed in the context of climatic fluctuations.
It may be possible to associate the growth reaction with the prolonged positive phase of February AO and March temperatures (Fig. 5), which were, based on response functions (Fig. 4), both found to have significant impact on growth. The positive anomalies of both of these variables are probably interrelated, and associated with the significant excursion of the winter-time AO, with prolonged positive phase in the recent past (Thompson and Wallace 1998).
While the site dryness was previously shown to diminish the pine thinning response (Hynynen and Arola 1999), precipitation in May was consistently found influence the pre-thinning growth positively in this study (Fig. 4). Similarly, the growth reduction that occurred during the end of the 1960s in both PAK and MIE stands was evidently coeval to multi-year decrease in May precipitation (see Fig. 5). Interestingly, the notably wet May in 1989 coincided with the starting year of especially enhanced post-thinning growth (Fig. 3). Although transient in character, the early-summer wetness (in 1989) may have contributed to the initiation of the positive thinning response.
Centennial variation in pine growth
Timberline pine growth is characterized by ample year-to-year variations, visible in PAK, MIE and control chronology (Fig. 6). Growth in PAK and MIE was especially well correlated with each other, but also, albeit less well, with the control chronology (Table 1; Fig. 6a). In addition to short-term growth variations, considerable changes in the growth at longer time-scales occurred as well. In general, the 20th century has experienced markedly good radial growth compared to the 19th century (Fig. 6b). A favourable growth period occurred especially during the 1920s and 1930s. This was a positive afforestation and growth phase that was noted and discussed already by contemporaries (Hustich 1940, 1948; Mikola 1952, 1962) in association with the general amelioration of the regional climate during the same period of time (Rogers 1985; Przybylak 2000). This is also a period during which the northern timberline exhibited a clear increase in the regeneration success of pine (Hustich 1940, 1948; Sirén 1998; Juntunen et al. 2002) and during which the stands were seeded.
It is likely to studied stands benefited (especially PAK) from the advantageous climatic phase. Although the growth (and apparently the favourability of the climate) did not reach the level of the 1920s and 1930s during the rest of the century, it is notable that the growth of the natural pines in Lapland during the later part of the 20th century remained at a higher overall level than the 1800s growth (Fig. 6). Even the growth during the 1960s, which was a particularly unfavourable period in the region’s forestry (e.g. Pohtila 1979) was not comparable to the disadvantageous growth periods during the 19th century. The growth deterioration during the first decades of the 19th century was probable among the worst growth intervals over the past eight centuries (Fig. 6b). Actually, the 19th century has been thought to be perhaps the coldest interval in the Northern Hemisphere (Mann et al. 1999) and in the study region (Helama et al. 2002) over the past millennium, belonging to a multi-centennial climatic interval commonly referred to as the ‘Little Ice Age’, during which a cool climate is thought to have prevailed over wide area especially between AD 1570 and 1900 (Matthews and Briffa 2005). Apart from climatic factors, also the nitrogen deposition has been speculated to influence the growth of European forests positively over the recent times but we note that its influence is supposed to be low in northern Europe (Spiecker 1999).
Given that the years of unfavourable growth were also amongst the years of disadvantageous pine regeneration (Sirén 1961, 1993), the negative phases of growth curve (Fig. 6b) determine the historical intervals during which pine seed or seedling performance were likely to be absent or very poor, and vice versa. This may be a rough picture of the realms of the long-term timberline pine growth (∼forestry practices) possibilities in northernmost Lapland and it shows that during the past two centuries, temporally no less than the common life span of naturally grown pines, climate conditions have varied between the extremes, from the especially unfavourable 19th to the generally favourable 20th century. The highly variable past conditions indicate general instability of the regional climate also in the future, with concomitant influences on forestry practices.
Conclusions
Reforestation and afforestation activity that took place during the 1920s and 1930s have resulted in successfully seeded pine stands in Pakanajoki and Mierasrova. Tree-rings of these pines provide insights into the growth variations of relatively young stands in climatologically harsh timberline environment. One more interesting aspect comes from southern origin of seeds. Altogether, the stands serve the forest researchers an interesting and unique experiment.
It was found that while pre-thinning tree-ring variability in seeded stands closely resembled each other, the growth in the naturally regenerated stands in the region was evidently more dissimilar. The growth of naturally regenerated pines has previous shown to correlate strongly with mid-summer temperatures whereas the growth of seeded pines was only moderately related to mid-summer conditions. It is also presumable that the different genetic origin of seeded stands resulted in climate-growth relationships that were expressed as profound dependence on autumn climate in the previous year. Such relationship could potentially be attributed to the hardening processes of the trees to the dormancy via a later cessation of ring growth in pines of southern genetic origin.
It was also found that the pines were seeded and thinned during the periods that were rather favourable for both activities. The warmth of summers especially in 1930s contrasts sharply with the centennial summer cooling of the 1800s that preceded the 20th century climatic amelioration. Post-thinning growth occurred under the climate that likely benefited the reaction to increase in growing space. Prolonged positive phase of the Arctic Oscillation (Northern Annular Mode) in winter overlapped the post-thinning period with conceivable influence on the successful growth release. The results illustrate the might of climatic variability to determine the success of silvicultural practices near the distributional limits of the species.
References
Akaike H (1974) A new look at the statistical model identification. IEEE T Automat Contr AC-19:716–723
Biondi F (1997) Evolutionary and moving response functions in dendroclimatology. Dendrochronologia 15:139–150
Biondi F, Waikul K (2004) DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput Geosci 30:303–311
Box GEP, Jenkins GM (1970) Time series analysis: forecasting and control. Holden-Day, San Francisco
Brier GW (1948) Northern hemisphere surface pressure and climatic fluctuations and vegetation growth in northern Finland. Nature 161:730–731
Briffa KR, Jones PD, Bartholin TS, Eckstein D, Schweingruber FH, Karlén W, Zetterberg P, Eronen M (1992) Fennoscandian summers from AD 500: temperature changes on short and long timescales. Clim Dyn 7:111–119
Briffa KR, Jones PD, Pilcher JR, Hughes MK (1988) Reconstructing summer temperatures in northern Fennoscandia back to AD 1700 using tree-ring data from Scots pine. Arct Alp Res 20:385–394
Cook ER (1985) A time series analysis approach to tree-ring standardization. Dissertation, University of Arizona, Tucson
Cook ER (1990) Conceptual linear aggregate model for tree rings. In: Cook ER, Kairiukstis LA (eds) Methods of dendrochronology: applications in the environmental science. Kluwer Academic Publishers, Dordrecht, pp 98–104
Cook ER, Briffa KR, Meko DM, Graybill DA, Funkhouser G (1995) The ‘segment length curse’ in long tree-ring chronology development for palaeoclimatic studies. Holocene 5:229–237
Erlandsson S (1936) Dendrochronological studies. Stockholms Högskolas Geokronological Institute, Report 23:1–116
Fritts HC (1976) Tree rings and climate. Academic Press, London
Fritts HC, Mosimann JE, Bottorff CP (1969) A revised computer program for standardizing tree-ring series. Tree-Ring Bull 29:15–20
Fritts HC, Blasing TJ, Hayden BP, Kutzbach JE (1971) Multivariate techniques for specifying tree-growth and climate relationships and for reconstructing anomalies in paleoclimate. J Appl Meteorol 10:845–864
Guiot J (1986) ARMA techniques for modelling tree-ring response to climate and for reconstructing variations of paleoclimates. Ecol Model 33:149–171
Haapanen T, Hari P, Kellomäki S (1979) Effect of fertilization and thinning on radial growth of Scots pine. Silva Fenn 13(2):184–189
Hari P, Sirén G (1972) Influence of some ecological factors and the seasonal stage of development upon the annual ring width and radial growth index. Royal College of Forestry, Department of Reforestation, Stockholm, Research Notes 40:1–22
Helama S, Lindholm M, Timonen M, Meriläinen J, Eronen M (2002) The supra-long Scots pine tree-ring record for Finnish Lapland: part 2, interannual to centennial variability in summer temperatures for 7500 years. Holocene 12:681–687
Helama S, Holopainen J, Timonen M, Ogurtsov MG, Lindholm M, Meriläinen J, Eronen M (2004a) Comparison of living-tree and subfossil ring-widths with summer temperatures from 18th, 19th and 20th centuries in northern Finland. Dendrochronologia 21:147–154
Helama S, Lindholm M, Timonen M, Eronen M (2004b) Detection of climate signal in dendrochronological data analysis: a comparison of tree-ring standardization methods. Theor Appl Climatol 79:239–254
Helama S, Lindholm M, Meriläinen J, Timonen M, Eronen M (2005a) Multicentennial ring-width chronologies of Scots pine along north-south gradient across Finland. Tree-Ring Res 61:21–32
Helama S, Timonen M, Lindholm M, Meriläinen J, Eronen M (2005b) Extracting long-period climate fluctuations from tree-ring chronologies over timescales of centuries to millennia. Int J Climatol 25:1767–1779
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–75
Hurrell JW (1995) Decadal trends in the North Atlantic Oscillation and relationships to regional temperature and precipitation. Science 269:676–679
Hurrell JW, Kushnir Y, Visbeck M (2001) The North Atlantic Oscillation. Science 291:603–604
Hustich I (1940) Tallstudier sommaren 1939 i Enare och Ustjoki. Acta Soc Fauna Flora Fenn 62(6):1–75
Hustich I (1948) The Scotch pine in northernmost Finland and its dependence on the climate in the last decades. Acta Bot Fenn 42:1–75
Hustich I (1966) On the forest-tundra and northern tree-lines, a preliminary synthesis. Rep Kevo Subarctic Res Stat 3:1–47
Hustich I, Elfving G (1944) Die Radialzuwachsvaariationen der Waldgrenzkiefer. Soc Sci Fenn Comm Biologicae 9(8):1–18
Hynynen J, Arola M (1999) Ensiharvennusajankohdan vaikutus hoidetun männikön kehitykseen ja harvennuksen kannattavuuteen. Metsätieteen aikakauskirja 1999(1):5–23
Ilvessalo Y (1970) Natural development and yield capacity of forest stands on mineral soils in northern Lapland. Acta For Fenn 108:1–43
Jalkanen R (1993) Defoliation of pines caused by injury to roots resulting from low temperatures. The Finnish Forest Research Institute, Research Papers 451:77–88
Juntunen V, Neuvonen S, Norokorpi Y, Tasanen T (2002) Potential for timberline advance in northern Finland, as revealed by monitoring during 1983–99. Arctic 55:348–361
Kirchhefer AJ (1999) Pine growth and climate AD 1800–1992 along a transect across the Scandes at 69°N. In: Kirchhefer AJ (ed) Dendroclimatology on Scots pine (Pinus sylvestris L.) in northern Norway. Dissertation, Department of Biology, University of Tromsø
Kirchhefer AJ (2000) The influence of slope aspect on radial increment of Pinus sylvestris L. in northern Norway and its implications for climate reconstructions. Dendrochronologia 18:27–40
Kirchhefer AJ (2001) Reconstruction of summer temperature from tree ring of Scots pine, Pinus sylvestris L., in coastal northern Norway. Holocene 11:41–52
Linderholm HW (2002) 20th century Scots pine growth variations in the central Scandinavian Mountains related to climate change. Arct Antarct Alp Res 34:378–387
Lindholm M (1996) Reconstruction of past climate from ring-width chronologies of Scots pine (Pinus sylvestris L.) at the northern forest limit in Fennoscandia. Dissertation, University of Joensuu
Lindholm M, Lehtonen H, Kolström T, Meriläinen J, Eronen M, Timonen M (2000) Climatic signals extracted from ring-width chronologies of Scots pine from the northern, middle and southern parts of the boreal forest belt in Finland. Silva Fenn 34:317–329
Macias M, Timonen M, Kirchhefer A, Lindholm M, Eronen M, Gutierrez E (2004) Growth variability of Scots pine along a west-east gradient across northern Fennoscandia: a dendroclimatic approach. Arct Antarct Alp Res 36:565–574
Mann ME, Bradley RS, Hughes MK (1999) Northern hemisphere temperatures during the past millennium: inferences, uncertainties, and limitations. Geophys Res Lett 26:759–762
Matalas NC (1962) Statistical properties of tree ring data. Bull Int Assoc Sci Hydrol 7:39–47
Matthews JA, Briffa KR (2005) The ‘little ice age’: re-evaluation of an evolving concept. Geogr Ann 87A:17–36
Mikkola K, Virtanen T (2006) Ilmaston ja maastonmuotojen vaikutus männyn metsänrajaan. Working Papers of the Finnish Forest Research Institute 25:14–24
Mikola P (1950) Puiden kasvun vaihteluista ja niiden merkityksestä kasvututkimuksessa. Summary in English: on the varitions in tree growth and their significnce to growth studies. Comm Inst For Fenn 38(5):1–131
Mikola P (1952) Havumetsien viimeaikaisesta kehitytksestä metsänrajaseudulla. Summary in English: on the recent development of coniferous forests in the timber-line region of northern Finland. Comm Inst For Fenn 40(2):1–32
Mikola P (1962) Temperature and tree growth near the northern timber line. In: Kozlowski TT (ed) Tree growth. Ronald Press, New York, pp 265–274
Mirov NT (1967) The genus Pinus. Ronald Press, New York
Monserud RA (1986) Time-series analyses of tree-ring chronologies. For Sci 32:349–372
Norokorpi Y (1994) Havumetsänrajan sijainnin määräytyminen Suomessa. Metsäntutkimuslaitoksen tiedonantoja 539:7–15
Pohtila E (1979) 1960-luvun vastoinkäymiset. Silva Fenn 13(1A):1–50
Pohtila E (1980) Climatic fluctuations and forestry in Lapland. Holarct Ecol 3:91–98
Pohtila E Timonen M (1980) Suojametsäalueen viljelytaimikot ja niiden varhaiskehitys. Summary in English: scots pine plantations and their early development in the protection forests of Finnish Lapland. Folia For 453:1–18
Przybylak R (2000) Temporal and spatial variation of surface air temperature over the period of instrumental observations in the Arctic. Int J Climatol 20:587–614
Renvall A (1912) Die periodischen Erscheinungen der Reproduktion der Kiefer an der polaren Waldgrenze. Acta For Fenn 1:1–155
Rogers JC (1985) Atmospheric circulation changes associated with the warming over the northern North Atlantic in the 1920s. J Clim Appl Meteorol 24:1303–1310
Ruha T, Varmola M (1997) Precommercial thinning in naturally regenerated Scots pine stands in northern Finland. Silva Fenn 31:401–415
Savva JV, Vaganov EA (2006) Genetic and environmental effects assessment in Scots pine provenances planted in central Siberia. Mitigat Adapt Strat Global Change 11:269–290
Savva YV, Kuz’mina NA, Vaganov EA (2001) Sensitivity of the structural characteristics of tree rings to climatic changes in different climatypes of Scotch pine. Russ J Ecol 32:400–407
Savva YV, Schweingruber FH, Vaganov EA, Milyutin LI (2003) Influence of climate changes on tree-ring characteristics of Scots pine provenences in southern Siberia (forest-steppe). IAWA J 24:371–383
Schove DJ (1950) Tree rings and summer temperatures, A.D. 1501–1930. Scot Geogr Mag 66:37–42
Sirén G (1961) Skogsgränstallen som indikator för klimatfluktuationerna i norra Fennoskandien under historisk tid. Summary in English. Comm Inst For Fenn 54:1–66
Sirén G (1993) Pine seed-year frequency in the subarctic of Finland: a pilot study. World Resour Rev 5:95–103
Sirén G (1998) Results and conclusions of pine advance in subarctic Finland in the 20th century. The Finnish Forest Research Institute, Research Papers 677:7–16
Spiecker H (1999) Overview of recent growth trends in European forests. Water Air Soil Poll 116:33–46
Spiecker H (2002) Tree rings and forest management in Europe. Dendrochronologia 20:191–202
Thompson DWJ, Wallace JM (1998) The arctic oscillation signature in the wintertime geopotential height and temperature fields. Geophys Res Lett 25:1297–1300
Thompson DWJ, Wallace JM (2000) Annular modes in the extratropical circulation. Part I: month-to-month variability. J Clim 13:1000–1016
Timonen M (2005) Muuttuvan ilmaston ennakointi metsän uudistamisessa. In: Hyppönen M, Hallikainen V, Jalkanen R (eds) Metsätaloutta kairoilla—Metsänuudistaminen Pohjois-Suomessa. Kustannusosakeyhtiö Metsälehti
Uvo CB, Berndtsson R (2002) North Atlantic Oscillation; a climatic indicator to predict hyrdopower availability in Scandinavia. Nord Hydrol 33:415–424
Vaganov EA, Hughes MK, Shashkin AV (2006) Growth dynamics of conifer tree rings. Images of past and future environments. Ecol Stud 183:1–354
Varmola M, Hyppönen M, Mäkitalo K, Mikkola K, Timonen M (2004a) Forest management and regeneration success in protection forests near the timberline in Finnish Lapland. Scand J For Res 19:424–441
Varmola M, Salminen H, Timonen M (2004b) Thinning response and growth trends of seeded Scots pine stands at the arctic timberline. Silva Fenn 38(1):71–83
Veijola P (1998) The northern timberline and timberline forests in Fennoscandia. The Finnish Forest Research Institute, Research Papers 672:1–242
Acknowledgements
Funding provided by the foundation of Marjatta ja Eino Kollin Säätiö (to SH) enabled the dendrochronological analyses to be carried out, whereas the post-doctoral scholarship from the foundation of Koneen Säätiö (to SH) made the final preparation of the manuscript possible. These grants are greatly acknowledged. Metsähallitus—Northern Lapland District for Wilderness Management is acknowledged for their support for remeasurements. For. Techn. Tauno Luosujärvi was responsible for most of the field works and tree-ring measurements together with For. Techn. Hannu Herva, their work is acknowledged. Figure 1 was aided by Online Map Creation. Comments from three anonymous referees greatly improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Helama, S., Salminen, H., Timonen, M. et al. Dendroclimatological analysis of seeded and thinned Scots pine (Pinus sylvestris L.) stands at the coniferous timberline. New Forests 35, 267–284 (2008). https://doi.org/10.1007/s11056-007-9076-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-007-9076-1