
Abstract
Members of the Fusarium solani species complex (FSSC) are causing the majority of the fusariosis in humans. Disseminated fusariosis has a high mortality and is predominantly observed in patients with leukemia. Here, we present the case of a fatal infection by a Fusarium strain with a degenerated phenotype, in a patient with acute lymphatic leukemia. Multiple nasal and skin biopsies as well as blood cultures yielded fungal growth, while in direct and histopathological examination of biopsy material septate hyphae were visible. Initial colonies were white with slimy masses with microconidia reminiscent of Fusarium/Acremonium, but with conidiospore production directly on the hyphae. Multi-locus sequence typing discerned a pionnotal—morphologically degenerated—colony of the recently recognized F. petroliphilum as etiological agent. The culture returned to a typical F. solani species complex morphology only after several weeks of growth in culture. Antifungal susceptibility tests indicate amphotericin B as best drug for this FSSC member rather than any of the azoles or echinocandins.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The types of infection caused by Fusarium spp. in humans range from infections of nail, skin, and eye in immunocompetent hosts to invasive and disseminated infections in mainly immunocompromised patients [1–3]. In particular, the different types of leukemic patients are vulnerable to disseminated infections. For disseminated fusariosis, prolonged neutropenia and T cell immunodeficiency are two of the main risk factors [1]. Disseminated fusariosis is often fatal and presents with characteristic skin lesions with necrotic center and positive blood cultures.
At least six species complexes and several single species with the large genus Fusarium have been responsible for human infections. Such species complexes are often species rich, while many sibling species are morphologically near indistinguishable. The Fusarium solani species complex (FSSC) members are responsible for approximately two-thirds of all cases of fusariosis [4]. Besides causing human infections, members of FSSC are commonly encountered as economically important plant pathogens as well as saprophytes [5].
To recognize the different sibling species within the FSSC, we currently rely on multi-locus sequence typing (MLST) [4–7]. Many of the recognized MLST haplotypes have no binomial names as yet, but some were recently identified, like F. petroliphilum and F. keratoplasticum [8]. Some morphologically recognizable etiological agents like F. falciforme and F. lichenicola lacking the fusiform macroconidia typically associated with Fusarium have now also been placed based on MLST data within the FSSC [9]. A specially curated database is available for the identification of Fusarium strains based on sequence data [6]. However, isolated etiological agents may not be recognized at first to belong to the genus Fusarium as pionnotal—degenerated—appearances that are frequently encountered in human isolates, especially in the FSSC [2, 8].
Guidelines for the treatment of deep and disseminated infections by Fusarium and other hyalohyphomycoses were recently published by a consortium of experts [10]. Fusarium species are known for their refractory behavior toward antifungals [11]. Treatment options seem limited to amphotericin B, voriconazole, and posaconazole, while echinocandins are better avoided [10]. Within species complexes variation exists in susceptibility for specific antifungal compounds [e.g., 12, 13]. Occasionally, human isolates and species that are multiple resistant to all available drugs are encountered [14]. With (MLST) identification of species level rather than (morphological) recognition of species complexes and with antifungal susceptibility test (AFST) data on species level, we may tailor patient treatment better to the encountered etiological agent.
Case Presentation
The 48-year-old female patient who had symptoms of shortness of breath and palpitations was hospitalized with the diagnosis of acute lymphoblastic leukemia to the hematology department of the Uludag University Hospital. The patient developed fever on the second day of remission induction therapy. The initial clinical suspicion was a community-acquired infection, and treatment with cefepime (3× 2 g day−1 i.v.) and clarithromycin (2× 500 mg day−1 for 10 days i.v.) was started. In spite of an initial decrease in fever, the patient’s body temperature increased again on the 17th day of chemotherapy and cefepime was switched to imipenem (4× 500 mg day−1 i.v.). Due to the presence of a central venous line, vancomycine (2× 1 g day−1 i.v.) was also added. Two days later, a black-crusted lesion was observed in the patient’s nasal mucosa. Although the paranasal sinus and chest computerized tomography (CT) did not indicate a fungal infection, fresh microscopic examination (KOH %10) of the nasal biopsy sample revealed septate hyphae, and liposomal amphotericin B at 3 mg/kg/day was added to patient’s regimen on the 19th day of the chemotherapy. On the second day of amphotericin B therapy, the patient had another episode of dyspnea and the chest X-ray indicated bilateral pneumonia. Thus, levofloxacin (1× 500 mg day−1 i.v.) and trimethoprim-sulfamethoxazole (3× 960 mg day−1 i.v.) were added to patient’s regimen on the 20th day of chemotherapy.
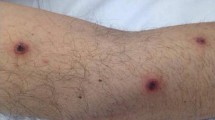
Septate hyaline hyphae were seen in histopathological examination of nasal biopsy, and culture of biopsy material yielded a growth of a mold resembling Fusarium/Acremonium spp. The patient developed 0.5–1-cm-wide maculopapular lesions with necrotic centers and hyperemic margins on the 22nd day of chemotherapy (Fig. 1). Two skin biopsy samples and a blood culture obtained during the febrile period yielded the same growth of mold resembling Fusarium/Acremonium spp. The patient’s condition gradually deteriorated, and on the 28th day of chemotherapy and on the 8th day of liposomal amphotericin B therapy, the patient died. The cultures were sent to the CBS-KNAW Fungal Biodiversity Centre, Netherlands for further identification.

Necrotic maculopapular skin lesions of the patient
Identification of the Fungal Etiological Agent
From nasal biopsy material (two times), skin biopsy materials (two times), and blood culture, mycelial growth was obtained. Direct and histopathological examination of the biopsy material showed hyaline septate hyphae. The colonies grown on Sabouraud-dextrose medium (SABG) were white, with slimy heads full with slightly crooked microconidia reminiscent of Acremonium or Fusarium. However, the colonies did not show typical conidiophores, but spores (Fig. 2a) were produced directly on the hyphae, where also small round scars were visible (Fig. 2b, c). The culture was deposited in the CBS culture collection, Utrecht, The Netherlands, under number CBS 135955.

a Microconidia that is predominantly 0-septate, oval, ellipsoid, and reniform. b Microconidium formed directly on the side of hyphae in the pionnotal mycelium. c Scar tissue on hyphae after microconidia production. d Reverted colonies with long conidiophores typical for members of the FSSC
DNA was isolated with a cetyltrimethyl ammonium bromide (CTAB)-based method [15]. The barcoding internally transcribed spacer (ITS) region and the 28S large ribosomal subunit (LSU) were used for identification of genus level; for identification of species level, partial transcription elongation factor 1 alpha (tef1α) and a subunit of the RNA polymerase (rpb2) sequences were used under conditions as described before [16]. The PCR fragments were sequenced with the ABI Prism_Big DyeTM Terminator v. 3.0 ready reaction cycle sequencing kit (Applied Biosystems, Foster City, CA) and analyzed on an ABI PRISM 3700 Genetic Analyzer (Applied Biosystems). Sequences were compared via BLAST to sequences in GenBank (http://www.ncbi.nlm.nih.gov/genbank/), and to the Fusarium MLST database (http://www.cbs.knaw.nl/Fusarium/) [6]. The obtained sequences (Genbank accession numbers KJ867425 (ITS), KJ867423 (LSU), KJ867424 (tef1α), and KJ867426 (rpb2)) identified the etiological agent as a FSSC type 2 strain and hence an isolate of F. petroliphilum.
Re-examination of older and subcultured colonies showed that the culture slowly reverted to morphology as expected from species belonging to the FSSC with long conidiophores and macroconidia (Fig. 2d). In this particular strain, the pionnotal phenotype proved reversible. Pionnotal Fusarium strains—even ones totally lacking spore production—have regularly been implicated in clinical infections [2, 8].
Antifungal Susceptibility Testing
In vitro AFST was performed for anidulafungin (ANI; Pfizer Central Research, UK), amphotericin B (AMB; Sigma, The Netherlands), fluconazole (FLC; Pfizer Central Research, Sandwich, UK), isavuconazole (ISA; Basilea Pharmaceuticals, Basel, Switzerland), itraconazole (ITC; Janssen-Cilag,Tilburg, The Netherlands), micafungin (MICA; Astellas, Japan), posaconazole (POS; Merck, Whitehouse Station, USA), and voriconazole (VOR; Pfizer Central Research). All antifungal compounds were obtained from their manufacturers as pure powders. The CLSI microdilution method M38-A2 for filamentous fungi [17] was followed using treated 96-wells tissue culture plates from Costar®, RPMI1640 with glutamine without NaHCO3 from Gibco, and MOPS from Sigma. Minimal inhibitory/effective concentrations (MIC/MEC) for CBS 135955 were ANI (>8 mg/l), AMB (1 mg/l), FLC (64 mg/l), ISA (>16 mg/l), ITC (>16 mg/l), MICA (>8 mg/l), POS (>16 mg/l), and VOR (8 mg/l); the results for the other tested F. petroliphilum strains from the CBS collection were similar (Table 1).
Comparing these results to other published AFST data for the FSSC or particular members of the FSSC, we can observe that the FSSC MICs of VOR are in the range from 1 to 32 mg/l, hence from susceptible to resistant, whereas all tested F. petroliphilum had MIC values of ≥8 mg/ml (Table 2). For ANI, FLC, ISA, ITC, MICA, and POS both F. petroliphilum and the other FSSC members are resistant. Therefore, amphotericin B seems to be the drug with the lowest MIC against F. petroliphilum.
Discussion
In immunocompromised patients, Fusarium spp. either penetrate the skin barrier or cause a pulmonary infection by airborne transmission. In many patients, skin lesions may be the first sign of a disseminated infection and are commonly seen in the early stages of the disease [1, 18]. In 40 % of the cases, Fusarium species can be isolated from blood cultures. In the presence of skin lesions, the identification rate may increase up to 56 % [19]. In the great majority of patients, fungaemia developed within 5 days after skin lesions become evident [20]. In our patient, the causative agent was first isolated from a nasal biopsy, and skin lesions developed later. Similar as reported in the literature, the agent was isolated from blood cultures 4 days after the onset of skin lesions.
Survival rate of patients with fusariosis who are persistent neutropenic remains around 4 % in spite of aggressive therapy [19]. The recently published guidelines for treatment of fusariosis recommend amphotericin B and voriconazole for infections caused by Fusarium [10], while posaconazole—often used as prophylaxis—has also activity against some members of the genus Fusarium [1]. However, our antifungal susceptibility tests of the different F. petroliphilum isolates indicate that for this particular species in the FSSC, the azoles and echinocandins are not effective, leaving only amphotericin B as potentially effective treatment.
Despite the fact that our patient received amphotericin B therapy, she died. Hence, more important than the antifungal drug in effective treatment of disseminated fusariosis in a leukemic patient is reversal of neutropenia and T cell dysfunction. Disseminated infections in other types of immunocompromised patients like SOT patients or HIV patients are extremely rare [1, 2]. In non-leukemic patients, invasive fusariosis remains localized, suggesting that especially neutrophils and T cells have an important role in limiting spread of Fusarium via blood. The prognosis of disseminated fusariosis is poor and is determined largely by the degree of immunosuppression [1].
Fusarium petroliphilum is not a new species causing opportunistic infections in humans, only its status as separate species within the Fusarium solani species complex is recent [8]. We tested five clinical isolates, the oldest being CBS 224.34, first isolated from onychomycosis in 1934. The AFST tests of the 8 antifungal compounds proved very uniform, showing how refractory the species always have been to any antifungal compound available.
In clinical infections, pionnotal Fusarium strains—even ones totally lacking spore production—have regularly been described [2, 8]. In the etiological agent studied here, this degenerate pionnotal phenotype proved reversible. Re-examination of colonies that were subcultured again or kept for several weeks showed that our culture reverted to morphology as expected from species belonging to the FSSC. It seems that for the opportunistic pathogen Fusarium the complex conditions within the human body select for different growth characteristics. Direct production of microconidia on hyphal fragments in tissue could be a reason why Fusarium is one of the few fungi regularly isolated from blood cultures.
References
Nucci M, Anaissie E. Fusarium infections in immunocompromised patients. Clin Microbiol Rev. 2007;20:695–704.
de Hoog GS, Guarro J, Gené J, Figueras MJ. Atlas of clinical fungi. 3rd ed. Utrecht: CBS-KNAW Fungal Biodiversity Centre, CD-ROM; 2011.
Horn DL, Freifeld AG, Schuster MG, Azie NE, Franks B, Kauffman CA. Treatment and outcomes of invasive fusariosis: review of 65 cases from the PATH Alliance®registry. Mycoses 2014. doi:10.1111/myc.12212. [Epub ahead of print].
O’Donnell K, Sutton DA, Fothergill A, et al. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J Clin Microbiol. 2008;46:2477–90.
Zhang N, O’Donnell K, Sutton DA, et al. Members of the Fusarium solani species complex that cause infections in both humans and plants are common in the environment. J Clin Microbiol. 2006;44:2186–90.
O’Donnell K, Sutton DA, Rinaldi MG, et al. Internet-accessible DNA sequence database for identifying Fusaria from human and animal infections. J Clin Microbiol. 2010;48:3708–18.
Debourgogne A, Gueidan C, de Hoog S, Lozniewski A, Machouart M. Comparison of two DNA sequence-based typing schemes for the Fusarium solani species complex and proposal of a new consensus method. J Microbiol Methods. 2012;91:65–72.
Short DP, O’Donnell K, Thrane U, et al. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet Biol. 2013;53:59–70.
Summerbell RC, Schroers HJ. Analysis of phylogenetic relationship of Cylindrocarpon lichenicola and Acremonium falciforme to the Fusarium solani species complex and a review of similarities in the spectrum of opportunistic infections caused by these fungi. J Clin Microbiol. 2002;40:2866–75.
Tortorano AM, Richardson M, Roilides E, et al. ESCMID & ECMM joint guidelines on diagnosis and management of hyalohyphomycosis: Fusarium spp, Scedosporium spp, and others. Clin Microbiol Infect. 2014;20 Suppl s3:27–46.
Guarro J. Fusariosis, a complex infection caused by a high diversity of fungal species refractory to treatment. Eur J Clin Microbiol Infect Dis. 2013;32:1491–500.
Azor M, Gené J, Cano J, Guarro J. Universal in vitro antifungal resistance of genetic clades of the Fusarium solani species complex. Antimicrob Agents Chemother. 2007;51:1500–3.
Alastruey-Izquierdo A, Cuenca-Estrella M, Monzón A, et al. Antifungal susceptibility profile of clinical Fusarium spp. isolates identified by molecular methods. J Antimicrob Chemother. 2008;61:805–9.
Kebabcı N, van Diepeningen AD, Ener B, et al. Fatal breakthrough infection with Fusarium andiyazi: new multi-resistant aetiological agent cross-reacting with Aspergillus galactomannan enzyme immunoassay. Mycoses. 2013;57:249–55.
Möller EM, Bahnweg G, Sandermann H, Geiger HH. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucl Acids Res. 1992;20:6115–6.
Davari M, Wei SH, Babay-Ahari et al. Geographic differences in trichothecene chemotypes of Fusarium graminearum in the Northwest and North of Iran. World Mycotoxin J. 2013;6:137–50.
Clinical and Laboratory Standards Institute. Reference method for broth dilution antifungal susceptibility testing of filamentous fungi. Approved standard. 2nd ed. Document M38-A2. Wayne, PA: Clinical and Laboratory Standards Institute; 2008.
Jossi M, Ambrosioni J, Macedo-Vinas M, Garbino J. Invasive fusariosis with prolonged fungemia in a patient with acute lymphobastic leukemia: case report and review of the literature. Int J Infect Dis. 2010;14:e354–6.
Nucci M, Anaissie EJ, Queinoz-Telles F, et al. Outcome predictors of 84 patients with hematologic malignancies and Fusarium infection. Cancer. 2003;98:315–9.
Grigis A, Farina C, Symoens F, Nolard N, Goglio A. Nosocomial pseudo-outbreak of Fusarium verticillioides associated with sterile plastic containers. Infect Control Hosp Epidemiol. 2000;21:50–2.
Hsiue HC, Ruan SY, Kuo YL, Huang YT, Hsueh PR. Invasive infections caused by non-Aspergillus moulds identified by sequencing analysis at a tertiary care hospital in Taiwan, 2000–2008. Clin Microbiol Infect. 2010;16:1204–6.
Azor M, Gené J, Cano J, Guarro J. Universal in vitro antifungal resistance of genetic clades of the Fusarium solani species complex. Antimicrob Agents Chemother. 2007;51:1500–3.
Nedret Koç A, Erdem F, Patıroĝlu T. Case report. Acremonium falciforme fungemia in a patient with acute leukaemia. Mycoses. 2002;45:202–3.
James EA, Orchard K, McWhinney PH, et al. Disseminated infection due to Cylindrocarpon lichenicola in a patient with acute myeloid leukaemia. J Infect. 1997;34:65–7.
Rodriguez-Villalobos H, Georgala A, Beguin H, et al. Disseminated infection due to Cylindrocarpon (Fusarium) lichenicola in a neutropenic patient with acute leukaemia: report of a case and review of the literature. Eur J Clin Microbiol Infect Dis. 2003;22:62–5.
Conflict of interest
J.F.M. received grants form Astellas, Merck and Basilea. He has been a consultant to Basilea and Merck and received speaker fees from Merck and Gilead.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ersal, T., Al-Hatmi, A.S.M., Dalyan Cilo, B. et al. Fatal Disseminated Infection with Fusarium petroliphilum . Mycopathologia 179, 119–124 (2015). https://doi.org/10.1007/s11046-014-9813-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11046-014-9813-x