
Abstract
Macrophage activation is modulated by both environmental cues and endogenous programs. In the present study, we investigated the role of a PAQR family protein, monocyte to macrophage differentiation-associated (MMD), in macrophage activation and unveiled its underlying molecular mechanism. Our results showed that while MMD expression could be detected in all tissues examined, its expression level is significantly up-regulated upon monocyte differentiation. Within cells, EGFP–MMD fusion protein could be co-localized to endoplasmic reticulum, mitochondria, Golgi apparatus, but not lysosomes and cytoplasm. MMD expression is up-regulated in macrophages after LPS stimulation, and this might be modulated by RBP-J, the critical transcription factor of Notch signaling. Overexpression of MMD in macrophages increased the production of TNF-α and NO upon LPS stimulation. We found that MMD overexpression enhanced ERK1/2 and Akt phosphorylation in macrophages after LPS stimulation. Blocking Erk or Akt by pharmacological agent reduced TNF-α or NO production in MMD-overexpressing macrophages, respectively. These results suggested that MMD modulates TNF-α and NO production in macrophages, and this process might involves Erk or Akt.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Macrophages are important immune cells in host. These diverse cell populations act as effector cells via phagocytosis and cytotoxicity in the innate immunity responses, and also participate in the adaptive immune responses by antigen presentation and cytokine production. Moreover, macrophages display notable plasticity and respond to environmental cues in a polarized manner, giving rise to different populations of cells with distinct physiological functions [1]. The polarized macrophages have been summarily classified into two subgroups: M1 and M2 macrophages. M1 polarization, which is triggered by type 1 inflammatory cytokines such as tumor necrosis factor α (TNF-α) or bacterial lipopolysaccharide (LPS), enhances microbicidal or tumoricidal capacity by producing reactive oxygen species (ROS) and nitric oxide (NO), and promotes inflammatory immune responses by the production of pro-inflammatory cytokines such as interleukin (IL)-12 and TNF-α [1]. In contrast, M2 macrophages play a key role in the resolution of inflammation via enhancing production of anti-inflammatory cytokine IL-10 [2, 3].
The endogenous molecular mechanisms for determining macrophage polarization are still poorly understood. The Notch signaling pathway is highly conserved and is involved in multiple cell fate decisions and differentiation processes during hematopoiesis. There are five Notch ligands, including Jagged (Jag) 1 and 2, and delta-like (Dll) 1, 3, 4, and four Notch receptors (Notch1–4) expressed on mammalian cell membrane. Ligand binding triggers proteolytic cleavages of Notch receptors, releasing the intracellular domain of Notch (NICD) to translocate into the nucleus, where NICD associates with the transcription factor RBP-J and recruits other co-activators to activate target gene expression [4]. Several reports have shown that Notch signaling participated in the regulation of macrophage activation and function [5–13]. Our recent study has shown that Notch signaling plays a critical role in the determination of M1 versus M2 polarization of macrophages in anti-tumor immune responses [14]. To further characterize the role of RBP-J-mediated canonical Notch signaling in macrophage activation, we performed a preliminary comparison of gene expression profiles between RBP-J deficient and normal macrophages stimulated with LPS, and the preliminary microarray data suggested that monocyte to macrophage differentiation-associated (MMD) was one of the differentially expressed genes among others (data not shown).
The human MMD (hMMD) gene was first identified in 1995, which is high expressed in mature, in vitro differentiated macrophages but absent in monocytes [15]. While bioinformatical analysis have shown that hMMD gene encodes a putative seven transmembrane (7TM) protein containing 238 amino acid residues, its role in macrophages activation has not been elucidated. In the present study, we report that the monocyte to MMD [15], a member of the progesterone and adipoQ receptor (PAQR) family [16], is involved in macrophage activation.
Materials and methods
Cell culture and transfection
Macrophage cell line RAW264.7 and dendritic cell (DC) line DC2.4 were cultured in RPMI1640 (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS), 2 mM l-glutamine, 50 IU/ml penicillin, and 50 μg/ml streptomycin sulfate. LPS (Sigma, St. Lowis, MI, USA) was used at 1 μg/ml. In some cases, MEK1 inhibitor PD98059 (50 μM) or phosphatidylinositol 3-kinase (PI3K) inhibitor LY294002 (25 μM) (Beyotime Inst. Biotechnol., Shanghai, China) was included in the culture medium.
RAW264.7 cells were transfected with Lipofectamine® LTX reagent or PLUS® reagent (Invitrogen) according to the recommended protocols. Cells were then cultured in the presence of 0.8 mg/ml G418 (Invitrogen) to select stable cell lines. Positive cells were sorted by FACS for the expression of enhanced green fluorescence protein (EGFP), pooled, and were used further analysis. For transient transfection, the transfected cells were cultured in medium for 36 h, and were observed under a fluorescent microscopy (BX51, Olympus, Tokyo, Japan) with a CCD camera (DP70, Olympus) or a confocal microscope (FV1000, Olympus). Organelle staining was performed using different tracking kits (Beyotime Inst. Biotechnol., Shanghai, China) following the manufacturer’s instructions, and nuclei were stained with DAPI. For immunofluorescence staining, cells were fixed with 4% paraformaldehyde and stained with FITC-conjugated anti-Flag (M2, Sigma, St. Lowis, MI, USA). Images were taken using a fluorescence microscope.
For the primary culture of bone marrow-derived macrophages (BMDMs), monocytes were isolated from BM of normal wild type mice, or poly(I)-poly(C)-induced MxCre-RBP-Jf/f (RBP-J−/−) or MxCre-RBP-J+/f (control) mice [14], by using anti-Gr1 (Ly-6G) magnetic beads (Miltenyi Biotec GmbH, Germany). Cells were cultured in the presence of mouse granulocyte macrophage-colony stimulating factor (mGM-CSF, 40 ng/ml) (Pepro Tech Inc.) for 7 days to generate macrophages [14].
Reverse transcription-polymerase chain reaction (RT-PCR) and real-time PCR
Total RNA was extracted from mouse tissues or cells with the TRIzol reagent (Invitrogen) and was reverse transcribed using a PrimeScript 1st Strand cDNA synthesis kit (TaKaRa, Dalian, China) according to the manufacturer’s instructions. The cDNA of MMD was amplified by PCR using primers 5′-TGGATCAATGCGGTTCAGGA-3′ and 5′-GCAATTGGCAGCATGTTCGTAG-3′. The β-actin cDNA was amplified with 5′-CATCCGTAAAGACCTCTATGCCAAC-3′ and 5′-ATGGAGCCACCGATCCACA-3′, as a reference control. The PCR products were analyzed by agarose gel electrophoresis. Real-time PCR was performed using a kit (SYBR Premix EX Taq, TaKaRa) and the ABI PRISM 7500 real-time PCR system following the manufacturer’s protocol, with β-actin as a reference control. Other primers used in real-time PCR included: TNF-α, 5′-CAGGAGGGAGAACAGAAACTCCA-3′ and 5′-CCTGGTTGGCTGCTTGCTT-3′; iNOS, 5′-GCAGAGATTGGAGGCCTTGTG-3′ and 5′-GGGTTGTTGCTGAACTTCCAGTC-3′.
Plasmid construction
The coding region of the mouse MMD cDNA was amplified by RT-PCR from the total RNA of the mouse macrophage cell line RAW264.7 using the forward primer 5′-ATGCGGTTCAGGAATCGATTCCAG-3′ and the reverse primer 5′-TCATAAATGCCGAATAAAGTCTGTGGGAC-3′. To construct the pIRES2–EGFP–MMD plasmid, the MMD cDNA fragment was cloned into pIRES2–EGFP vector (Clontech) and was confirmed by sequencing. The MMD cDNA was also inserted in frame into pEGFP–N1 or pEGFP–C2, to construct pEGFP–N1–MMD and pEGFP–C2–MMD, which express MMD–EGFP and EGFP–MMD, respectively. To fuse Flag tag with MMD at N terminus, MMD cDNA was inserted in frame into pFlag–CMV2, to obtain pFlag–CMV2 MMD. C terminal Flag fusion was achieved by PCR and was expressed in pcDNA3.1(+). The pIRES2–EGFP–NIC plasmid was described previously [17].
Enzyme-linked immunosorbent assay (ELISA)
The protein level of TNF-α in culture supernatant was assayed using ELISA with an ELISA Kit (NeoBiosciences, Shanghai, China) according to the manufacturer’s instructions.
NO production
The level of NO in culture supernatant was assayed by Griess reaction with a Nitric Oxide Assay Kit (Beyotime Inst. Biotechnol.). The culture supernatants (50 μl) were added to 50 μl Griess reagent I and 50 μl Griess reagent II, and the absorbance at 540 nm was measured with a microplate reader.
Western blot analysis
To detect the expression and phosphorylation of Akt and ERK1/2 in macrophages, western blot analysis was performed routinely, with primary antibodies including: anti-Akt, anti-phospho-Akt (Ser473) (Signalway Antibody, Pearland, TX, USA), anti-p44/42 MAPK, anti-phospho-p44/42 MAPK (Thr202/Tyr204) (Cell Signaling Technology, Boston, MA, USA), or anti-β-actin (Sigma, St. Louis, MO, USA). As secondary antibodies, horseradish peroxidase (HRP)-conjugated anti-rabbit-IgG or anti-mouse-IgG (Boster BioTec) was used.
Statistics
The statistical analysis was performed with the SPSS 11.0 program. Results were expressed as the means ± SD. Comparison between groups was undertaken using the unpaired student’s t test. P < 0.05 was considered statistically significant.
Results
The expression pattern and intracellular localization of MMD
To further investigate the molecular mechanisms of Notch signaling in macrophage activation, we preliminarily compared the gene expression profiles between RBP-J-deficient (RBP-J−/−) and control (RBP-J+/−) BMDMs [14] after LPS stimulation by using microarray analysis. The data suggested that MMD was one of the differentially expressed genes among others (data not shown).
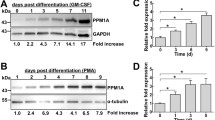
To examine the expression pattern of MMD, we prepared total RNA from different adult tissues, and performed RT-PCR. As shown in Fig. 1a, MMD mRNA could be detected in all examined tissues. However, when we performed similar experiment using RNA from different cell lines and sorted hematopoietic cells [14], we found that MMD was expressed in macrophages, DCs, T-lymphocytes, but not monocytes (Fig. 1b), suggesting that MMD expression might be activated during monocyte differentiation into macrophages and DCs. Indeed, when BM-derived monocytes were induced to differentiate into macrophages, the expression of MMD was up-regulated significantly (Fig. 1c), consistent with previous result [15].

MMD expression is up-regulated upon monocyte differentiation. a Total RNA was prepared from the indicated tissues, reverse transcribed, and cDNA fragment of MMD was amplified by PCR, with β-actin as a reference control. b The cDNA fragment of MMD was amplified by PCR from RNA of the indicated cell lines and sorted cells, with β-actin as a reference control. c BM-derived monocytes were cultured in the presence of GM-CSF to differentiate into macrophages, and mRNA expression of MMD was detected by real-time RT-PCR on the indicated days. Data are means ± SD of three independent experiments
We then examined the intra-cellular localization of MMD. RAW264.7 macrophages were transfected transiently with expression vectors in which MMD was fused with EGFP at either N- or C-terminus, and were observed under a fluorescence microscope. The result showed that the fusion protein had a dotted distribution within cells (supplemental Fig. S1). Similar results were obtained when cells were transfected with MMD fused with Flag tag at either N- or C-terminus (supplemental Fig. S1). We co-localized MMD–EGFP with different organelle trackers. The results showed that MMD–EGFP was co-localized to endoplasmic reticulum (ER), mitochondria, and Golgi apparatus, but not to lysosomes (Fig. 2). These data suggested that MMD localized to ER–Golgi apparatus and mitochondria, but not to lysosomes or cytoplasms.

The intracellular localization of MMD. RAW264.7 cells were transiently transfected with pEGFP–MMD. Cells were labeled with different organelle trackers, and were observed under a confocal fluorescence microscope. Nuclei were stained blue with DAPI. (Color figure online)
LPS up-regulated MMD expression in macrophages, which was modulated by RBP-J
To investigate whether MMD is involved in macrophage activation, RAW264.7 cells were treated with LPS for 2 h, and the expression of MMD was analyzed by real-time PCR. As shown in Fig. 3a, LPS mildly but significantly up-regulated MMD expression in macrophage. Activation of Notch signaling by the overexpression of NICD also up-regulated MMD mRNA (Fig. 3b). Moreover, primary BMDMs with RBP-J disruption exhibited decreased MMD expression compared with the control macrophages (Fig. 3c). These data suggested that LPS up-regulated MMD gene expression in macrophages, and this process was could be modulated by the RBP-J-mediated Notch signaling.

Notch signaling regulated MMD expression in macrophage. a RAW264.7 cells were stimulated with LPS for 12 h. b RAW–CON and RAW–NICD cells were stimulated with LPS for 12 h. c BMDMs from the RBP-J−/− and the control RBP-J+/− mice were stimulated with LPS for 12 h. MMD expression was analyzed by qRT-PCR. Data are means ± SD of three independent experiments. *P < 0.05
MMD overexpression increased TNF-α and NO production in RAW264.7 cells
LPS stimulates the production of various inflammatory mediators such as TNF-α and NO [1, 18]. To examine whether MMD is involved in these processes, RAW264.7 macrophages were stably transfected with MMD-expression vector (RAW–MMD). No change in cell growth or death was noticed in MMD–overexpressing RAW264.7 cells (data not shown). RAW–MMD and the control RAW–CON cells were stimulated with LPS for 12 h. qRT-PCR analysis for TNF-α and iNOS expression showed that the expression of TNF-α and iNOS mRNA in the RAW–MMD cells was increased (Fig. 4a, b). In agreement with the mRNA expression, the RAW–MMD cells stimulated with LPS produced more TNF-α and NO (Fig. 4c, d). These results indicated that MMD promoted LPS-stimulated production of TNF-α and NO.

MMD overexpression promoted TNF-α and NO production by LPS-stimulated RAW264.7 cells. RAW264.7 cells were stably transfected with MMD-expressing (RAW–MMD) and control (RAW–CON) vector, and stimulated with LPS for 12 h. The mRNA expression of TNF-α (a) and iNOS (b) were analyzed by qRT-PCR. The culture supernatants of the cells were harvested for the determination of TNF-α (c) and NO (d). Data are means ± SD of three independent experiments. *P < 0.05
MMD increased ERK1/2 and Akt phosphorylation in macrophages
It has been shown that LPS modulates TNF-α and NO production by activating multiple signaling pathways such as MAPK and PI3K signaling [18]. So, we stimulated the RAW–MMD and RAW–CON cells with LPS for indicated periods of time, and examined the effects of MMD overexpression on the phosphorylation of ERK1/2 and Akt by western blot analysis. As shown in Fig. 5a, LPS induced a notable ERK1/2 phosphorylation in RAW–CON cells after stimulation with LPS for 30 min, and lasted for a short time but can hardly be detected at 1 h after the stimulation. In contrast, ERK1/2 phosphorylation can be slightly detected in unstimulated RAW–MMD cells. Upon LPS stimulation, the level of phosphorylated ERK1/2 increased quickly, reached its highest levels at 1 h, and sustained up to 2 h. Furthermore, RAW–MMD cells displayed increased level of Akt phosphorylation as compared with the control cells. These results suggested that MMD increases ERK1/2 and Akt phosphorylation in macrophages.

MMD enhanced ERK1/2 and Akt phosphorylation. a Western blot analysis for the level of phospho-ERK1/2 and phospho-Akt in RAW–MMD and control RAW–CON cells, unstimulated or stimulated with LPS for indicated periods of time. Data represented three independent experiments. b, c RAW–MMD cells were treated with PD98059 (50 μM), LY294002 (25 μM) or DMSO (control) and were stimulated with LPS for 12 h. The culture supernatants were harvested for measuring TNF-α (b) and NO (c). Data are means ± SD of three independent experiments. *P < 0.05
Subsequently, RAW–MMD cells were treated with MEK1 inhibitor PD98059 or PI3K inhibitor LY294002, and were stimulated with LPS for 12 h. The culture supernatants were harvested and measured for the production of TNF-α and NO. As shown in Fig. 5b, c, blockade of ERK1/2 phosphorylation with PD98059 significantly decreased TNF-α production, whereas blockade of Akt phosphorylation with LY294002 significantly decreased NO production in LPS-stimulated RAW–MMD cells. These results suggested that MMD increased TNF-α and NO production in LPS-stimulated macrophages, but might through different signaling pathways.
Discussion
In an attempt to identify gene downstream to Notch signaling in the regulation of macrophage activation, we isolated MMD that might be involved in macrophage differentiation [15]. Although the mRNA expression of MMD is up-regulated in Notch activating macrophages and down-regulated in RBP-J deficient macrophages, Notch signaling appears not modulate the transcription of MMD directly, because bioinformatical analysis did not reveal RBP-J- nor Hes-binding site in the promoter of MMD. Notch signaling might indirectly influence the expression of MMD through other pathways [4], but further experiments are necessary to validate this speculation.
The exact biological function of MMD is not clear so far. The hMMD gene was first identified in 1995, which is high expressed in mature, in vitro differentiated macrophages but absent in monocytes [15]. Bioinformatical analysis shows that hMMD gene encodes a putative 7TM protein containing 238 amino acid residues [15]. In 2004, a macrophage/microglia activation factor (MAF) was identified, which is actually a MMD orthologue in rat and is expressed in lesion-associated microglial cells after brain trauma [19, 20]. Later, the MMD protein and other 10 7TM protein members sharing a common conserved 7TM motif were classified as a new protein family, the PAQR family, which is structurally and topologically different from the G protein-coupled receptor (GPCR) family [16]. According to the phylogenetic analysis, PAQR10 and 11, also known as MMD2 and MMD respectively, have exceptionally evolutionary rooting with substantially greater sequence similarity with the bacterial hemolysin III type protein. The MMD protein sequence is high conserved in different species, sharing 99% amino acid sequence identity between the orthologue of human and mouse [15]. The deep evolutionary rooting and high conservation of MMD protein suggest that MMD plays an important role, likely via effecting on some primary signaling pathways just as PAQR3, another PAQR member. PAQR3, also known as Raf kinase trapping to Golgi (RKTG), localizes at the Golgi apparatus and blocks the Ras/Raf/MEK/ERK signaling cascade via sequestrating Raf-1 to the Golgi apparatus and thus inhibits the proliferation and tumorigenicity of tumor cells [21–23]. Taking into consideration that LPS modulates TNF-α and NO production by activating multiple signaling pathways such as MAPK and PI3K signaling [24, 25], MMD might increase the production of TNF-α and NO in LPS-stimulated macrophages through ERK1/2 and Akt phosphorylation.
It has been generally known that the Ras/Raf/MEK/ERK signaling cascade takes place on the plasma membrane (PM). However, the PM is not the only platform for Ras/Raf/MEK/ERK signaling induction. Apart from the PM, Ras/Raf/MEK/ERK signaling induction also takes place in endomembranes, such as endosomes, the ER, and the Golgi apparatus [26–28]. The various biological consequences of ERK activation depend on not only the magnitude, but also the kinetics and subcellular compartment of ERK activity. For example, ERK activation stimulated by growth factor is transient from the PM but sustained from endosomes [29, 30]. During the negative versus positive selection of thymocytes, ERK activation from the PM induces programmed cell death, whereas ERK activation from the Golgi induces proliferation [31]. Our data showed that MMD overexpression in macrophages can not only increase the phosphorylation level of ERK, but also introduce a slower but more sustained ERK activation mediated by LPS compared with a rapid and transient signaling as control. This activation characteristic as well as the localization of MMD protein in endomembranes suggests that MMD presumably take part in the compartmentalization of ERK signaling. Moreover, our data showed that MMD also increased the phosphorylation level of Akt. It has been reported that in addition to Raf-1, Ras effectors also include PI3K, which leads to Akt activation [32]. So, we presume that MMD may spatially regulate activation of Ras, the common up-stream kinase of ERK and Akt.
References
Martinez FO, Sica A, Mantovani A, Locati M (2008) Macrophage activation and polarization. Front Biosci 13:453–461
Gordon S (2003) Alternative activation of macrophages. Nat Rev Immunol 3(1):23–35
Varin A, Gordon S (2009) Alternative activation of macrophages: immune function and cellular biology. Immunobiology 214(7):630–641
Kopan R, Ilagan MX (2009) The canonical Notch signaling pathway: unfolding the activation mechanism. Cell 137(2):216–233
Ohishi K, Varnum-Finney B, Serda RE, Anasetti C, Bernstein ID (2001) The Notch ligand, Delta-1, inhibits the differentiation of monocytes into macrophages but permits their differentiation into dendritic cells. Blood 98(5):1402–1407
Hu X, Chung AY, Wu I, Foldi J, Chen J, Ji JD, Tateya T, Kang YJ, Han J, Gessler M, Kageyama R, Ivashkiv LB (2008) Integrated regulation of toll-like receptor responses by Notch and interferon-gamma pathways. Immunity 29(5):691–703
Narayana Y, Balaji KN (2008) NOTCH1 up-regulation and signaling involved in Mycobacterium bovis BCG-induced SOCS3 expression in macrophages. J Biol Chem 283(18):12501–12511
Outtz HH, Wu JK, Wang X, Kitajewski J (2010) Notch1 deficiency results in decreased inflammation during wound healing and regulates vascular endothelial growth factor receptor-1 and inflammatory cytokine expression in macrophages. J Immunol 185(7):4363–4373
Monsalve E, Pérez MA, Rubio A, Ruiz-Hidalgo MJ, Baladrón V, García-Ramírez JJ, Gómez JC, Laborda J, Díaz-Guerra MJ (2006) Notch-1 up-regulation and signaling following macrophage activation modulates gene expression patterns known to affect antigen-presenting capacity and cytotoxic activity. J Immunol 176(9):5362–5373
Fung E, Tang SM, Canner JP, Morishige K, Arboleda-Velasquez JF, Cardoso AA, Carlesso N, Aster JC, Aikawa M (2007) Delta-like 4 induces notch signaling in macrophages: implications for inflammation. Circulation 115(23):2948–2956
Palaga T, Buranaruk C, Rengpipat S, Fauq AH, Golde TE, Kaufmann SH, Osborne BA (2008) Notch signaling is activated by TLR stimulation and regulates macrophage functions. Eur J Immunol 38(1):174–183
Monsalve E, Ruiz-García A, Baladrón V, Ruiz-Hidalgo MJ, Sánchez-Solana B, Rivero S, García-Ramírez JJ, Rubio A, Laborda J, Díaz-Guerra MJ (2009) Notch1 upregulates LPS-induced macrophage activation by increasing NF-kappaB activity. Eur J Immunol 39(9):2556–2570
Zhang W, Xu W, Xiong S (2010) Blockade of Notch1 signaling alleviates murine lupus via blunting macrophage activation and M2b polarization. J Immunol 184(11):6465–6478
Wang YC, He F, Feng F, Liu XW, Dong GY, Qin HY, Hu XB, Zheng MH, Liang L, Feng L, Liang YM, Han H (2010) Notch signaling determines the M1 versus M2 polarization of macrophages in antitumor immune responses. Cancer Res 70(12):4840–4849
Rehli M, Krause SW, Schwarzfischer L, Kreutz M, Andreesen R (1995) Molecular cloning of a novel macrophage maturation-associated transcript encoding a protein with several potential transmembrane domains. Biochem Biophys Res Commun 217(2):661–667
Tang YT, Hu T, Arterburn M, Boyle B, Bright JM, Emtage PC, Funk WD (2005) PAQR proteins: a novel membrane receptor family defined by an ancient 7-transmembrane pass motif. J Mol Evol 61(3):372–380
Yin DD, Fan FY, Hu XB, Hou LH, Zhang XP, Liu L, Liang YM, Han H (2009) Notch signaling inhibits the growth of the human chronic myeloid leukemia cell line K562. Leuk Res 33(1):109–114
Kawai T, Akira S (2006) TLR signaling. Cell Death Differ 13(5):816–825
Brauer AU, Nitsch R, Savaskan NE (2004) Identification of macrophage/microglia activation factor (MAF) associated with late endosomes/lysosomes in microglial cells. FEBS Lett 563(1–3):41–48
Lunemann A, Ullrich O, Diestel A, Jöns T, Ninnemann O, Kovac A, Pohl EE, Hass R, Nitsch R, Hendrix S, Jöns T, Ninnemann O, Kovac A, Pohl EE, Hass R, Nitsch R, Hendrix S (2006) Macrophage/microglia activation factor expression is restricted to lesion-associated microglial cells after brain trauma. Glia 53(4):412–419
Feng L, Xie X, Ding Q, Luo X, He J, Fan F, Liu W, Wang Z, Chen Y (2007) Spatial regulation of Raf kinase signaling by RKTG. Proc Natl Acad Sci USA 104(36):14348–14353
Fan F, Feng L, He J, Wang X, Jiang X, Zhang Y, Wang Z, Chen Y (2008) RKTG sequesters B-Raf to the Golgi apparatus and inhibits the proliferation and tumorigenicity of human malignant melanoma cells. Carcinogenesis 29(6):1157–1163
Xie X, Zhang Y, Jiang Y, Liu W, Ma H, Wang Z, Chen Y (2008) Suppressive function of RKTG on chemical carcinogen-induced skin carcinogenesis in mouse. Carcinogenesis 29(8):1632–1638
Oh JH, Lee TJ, Park JW, Kwon TK (2008) Withaferin A inhibits iNOS expression and nitric oxide production by Akt inactivation and down-regulating LPS-induced activity of NF-kappaB in RAW 264.7 cells. Eur J Pharmacol 599(1–3):11–17
Lahti A, Lähde M, Kankaanranta H, Moilanen E (2000) Inhibition of extracellular signal-regulated kinase suppresses endotoxin-induced nitric oxide synthesis in mouse macrophages and in human colon epithelial cells. J Pharmacol Exp Ther 294(3):1188–1194
Chiu VK, Bivona T, Hach A, Sajous JB, Silletti J, Wiener H, Johnson RL 2nd, Cox AD, Philips MR (2002) Ras signaling on the endoplasmic reticulum and the Golgi. Nat Cell Biol 4(5):343–350
Perez de Castro I, Bivona TG, Philips MR, Pellicer A (2004) Ras activation in Jurkat T cells following low-grade stimulation of the T-cell receptor is specific to N-Ras and occurs only on the Golgi apparatus. Mol Cell Biol 24(8):3485–3496
Roy S, Wyse B, Hancock JF (2002) H-Ras signaling and K-Ras signaling are differentially dependent on endocytosis. Mol Cell Biol 22(14):5128–5140
Oksvold MP, Skarpen E, Wierod L, Paulsen RE, Huitfeldt HS (2001) Re-localization of activated EGF receptor and its signal transducers to multivesicular compartments downstream of early endosomes in response to EGF. Eur J Cell Biol 80(4):285–294
Taub N, Teis D, Ebner HL, Hess MW, Huber LA (2007) Late endosomal traffic of the epidermal growth factor receptor ensures spatial and temporal fidelity of mitogen-activated protein kinase signaling. Mol Biol Cell 18(12):4698–4710
Daniels MA, Teixeiro E, Gill J, Hausmann B, Roubaty D, Holmberg K, Werlen G, Hollander GA, Gascoigne NR, Palmer E (2006) Thymic selection threshold defined by compartmentalization of Ras/MAPK signalling. Nature 444(7120):724–729
Gupta S, Ramjaun AR, Haiko P, Wang Y, Warne PH, Nicke B, Nye E, Stamp G, Alitalo K, Downward J (2007) Binding of ras to phosphoinositide 3-kinase p110alpha is required for ras driven tumorigenesis in mice. Cell 129(5):957–968
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (30830067, 91029731) and the Ministry of Science and Technology of China (2009CB521706, 2011ZXJ09101-02C).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Qiang Liu, Jin Zheng, Dan-Dan Yin, and Jie Xiang have contributed equally to this study.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Liu, Q., Zheng, J., Yin, DD. et al. Monocyte to macrophage differentiation-associated (MMD) positively regulates ERK and Akt activation and TNF-α and NO production in macrophages. Mol Biol Rep 39, 5643–5650 (2012). https://doi.org/10.1007/s11033-011-1370-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-011-1370-5