
Abstract
The objective of this study was to develop linear and nonlinear statistical models to predict enteric methane emission (EME) from cattle (Bos) in the tropics based on dietary and animal characteristic variables. A database from 35 publications, which included 142 mean observations of EME measured on 830 cattle, was constructed to develop EME prediction models. Several extant equations of EME developed for North American and European cattle were also evaluated for suitability of those equations in this dataset. The average feed intake and methane production were 7.7 ± 0.34 kg/day and 7.99 ± 0.39 MJ/day, respectively. The simple linear equation that predicted EME with high precision and accuracy was: methane (MJ/day) = 1.29(±0.906) + 0.878(±0.125) × dry matter intake (DMI, kg/day), [root mean square prediction error (RMSPE) = 31.0 % with 92 % of RMSPE being random error; R 2 = 0.70]. Multiple regression equation that predicted methane production slightly better than simple prediction equations was: methane (MJ/day) = 0.910(±0.746) + 1.472(±0.154) × DMI (kg/day) – 1.388(±0.451) × feeding level as a multiple of maintenace energy intake – 0.669(±0.338) × acid detergent fiber intake (kg/day), [RMSPE = 22.2 %, with 99.6 % of MSPE from random error; R 2 = 0.84]. Among the nonlinear equations developed, Mitscherlich model, i.e., methane (MJ/day) = 71.47(±22.14.6) × (1 - exp{−0.0156(±0.0051) × DMI (kg/day), [RMSPE = 30.3 %, with 97.6 % of RMSPE from random error; R 2 = 0.83] performed better than simple linear and other nonlinear models, but the predictability and goodness of fits of the equation did not improve compared with the multiple regression models. Extant equations overestimated EME, and many extant models had low accuracy and precision. The equations developed in this study will be useful for improved estimates of national methane inventory preparation and for a better understanding of dietary factors influencing EME for tropical cattle feeding systems.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Methane (CH4) production from the fermentation of feeds in the rumen represents a loss of up to 15 % of the gross energy (GE) intake depending upon the type of diets (Holter and Young 1992). Moreover, livestock production systems contribute a substantial share of greenhouse gases (GHG) to the atmosphere with an estimate of 7.1 gigatonnes (in CO2 equivalent) annually representing 14.5 % of total anthropogenic GHG (Gerber et al. 2013), and methane production from enteric fermentation represents about 40 % of total GHG emissions from livestock production systems (Gerber et al. 2013; Patra 2012, 2014a). For these purposes, several models had been developed since many years for predicting methane production and a better understanding of the dietary factors affecting this rumen fermentation process (e.g., Kriss 1930; Ramin and Huhtanen 2013; Patra and Lalhriatpuii 2016). However, all the models of prediction of methane production in cattle (Bos) were developed for North American and European temperate livestock production systems, and thus, the parameter estimates are more related to cattle raised in those regions.
Based on the data of Food and Agricultural Organization (http://faostat3.fao.org, accessed 24 October. 2015), more than 60 % of the global cattle populations are located in the tropical regions of the world, where animal production systems are markedly different from temperate livestock production. Basal feeds for ruminants in tropical environments are predominantly of low quality high fibrous forages (Van Soest 1994). Breeds of livestock species in the tropics also greatly differ from temperate regions, which may also differ for rumen function and digestive efficiency (Hegarty 2004). Several studies indicated that methane production, diversity, and abundances of methanogens in ruminants may vary depending upon diets (Bouchard et al. 2013; Hammond et al. 2013) and breeds (Hernandez-Sanabria et al. 2013; King et al. 2011; Zhou et al. 2010). Despite the considerable differences in feed composition and production systems between tropics and temperate environments, prediction models of methane emissions in cattle from tropical production systems have not been developed from a large data base spanning over wide variety of diets. Moreover, GHG emissions from livestock populations of the developing tropical regions are expected to grow increasingly in the years ahead due to expanding livestock populations (Gerber et al. 2013; Patra 2014a). Development of enteric methane prediction models is, therefore, required to improve estimates of methane outputs in tropical feeding systems.
Methane gas has high global warming potential (28 times greater than CO2), but short lifetime (12 years) in the atmosphere (IPCC 2013). Thus, addressing methane emissions is now considered as a quick and immediate mitigation strategy compared with other gases (IPCC 2013). Several mitigation options and technologies are suggested to decrease enteric methane production in ruminants, of which dietary strategies appear to be the most promising options (Hristov et al. 2013). A better understanding of the dietary composition influencing methane production would be useful for mitigation of methane production in tropical feeding conditions. The modelling approach is an effective tool to assess the effectiveness of different nutritional strategies to reduce methane production from ruminants (Benchaar et al. 2001).
A number of empirical and mechanistic models were developed for predicting methane emission from the studies or database of temperate cattle and sheep (Ovis) production (e.g., Blaxter and Clapperton 1965; Moe and Tyrrell 1979; Kebreab et al. 2008; Ramin and Huhtanen 2013). Statistical models predict methane production from nutrient intake, composition, feeding levels, and digestibility directly, while dynamic mechanistic models estimate methane emission using mathematical descriptions of rumen fermentation biochemistry (e.g., Benchaar et al. 2001; Mills et al. 2001; Kebreab et al. 2006; Ellis et al. 2007) and require complex data that are not available routinely. These models are useful to predict enteric methane emission from cattle without undertaking extensive and costly experiments and to explain the dietary factors exerting methane production. Therefore, the objectives of this study were to develop statistical models for prediction of enteric methane emissions for tropical cattle production systems using commonly measured dietary and animal variables and to compare these models with the extant methane prediction models developed for temperate cattle-rearing systems. The models also identified major dietary factors influencing methane production in tropical cattle production systems, which could be useful for nutritional mitigation of methane production.
2 Materials and methods
2.1 Construction of database
A database was compiled from the studies published in journals and conference proceedings for development of methane prediction models. Criteria for inclusion of studies in the database were that the studies provided in vivo methane production from tropical cattle description of the animals and intakes of dry matter (DM) and other nutrients. Overall, 35 publications (Bairagi and Mohini 2003; Bhar et al. 1999; De et al. 2012; Demarchi et al. 2003; Ghosh et al. 2001; Girdhar et al. 1995; Haque et al. 2001; Hulshof et al. 2012; Jain et al. 2011; Kannan and Garg 2009; Kannan et al. 2010; Kannan et al. 2011; Kennedy and Charmley 2012; Kurihara et al. 1999; Lal et al. 1987; Mohini and Mani 2007; Mohini et al. 2007; Malik and Singal 2008; Mohini et al. 2009; Mohini and Singh 2010; Mupeta et al. 2000; Nascimento et al. 2008; Neto et al. 2009; Oliveira et al. 2007; Pattanaik et al. 2003; Pedreira et al. 2009; Pedreira et al. 2012; Pedreira et al. 2013; Perna et al. 2013; Possenti et al. 2008; Primavesi et al. 2003; Primavesi et al. 2004; Rejil et al. 2008; Srivastava and Garg 2002; Tomkins et al. 2011) fulfilled the criteria for inclusion in this database. The studies in these publications were conducted in India (n = 19), tropical parts of Brazil (n = 12), and Australia (n = 3). One study from Zimbabwe was included in this database. There were many studies available from Australia, but studies conducted on tropical forage diets as stated in the publications were only considered in this database. Methane production was measured using either the sulphur hexafluoride (SF6) technique (n = 109) or respiration chamber (n = 40). Methane production measured by SF6 technique and respiration chamber study was 19.1 ± 6.66 versus 19.1 ± 4.50 g/kg DM intake and 5.81 ± 1.72 versus 5.93 ± 1.48 percent of GE intake, respectively, in this database. There were a total of 149 treatment means obtained from 830 observations from dairy and beef cattle. However, treatments (n = 7) containing feed additives with antimethanogenic properties were removed before statistical analyses.
The investigated dietary and animal factors (independent variables) were body weight (BW), intakes of DM, individual nutrients, GE and metabolizable energy (ME), organic matter and GE digestibility, nutrient composition of diets, forage proportion, and feeding level, which were used for regression equation development. Feeding level (FL) as multiple of maintenance was estimated by dividing the ME intake by the maintenance ME requirement for cattle in tropical countries (Kearl 1982). Because digestibility of nutrients changes with level of feeding, digestibility at maintenance level of feeding is the most consistent assessment of digestibility of feeds (Ramin and Huhtanen 2013). Thus, organic matter digestibility (g/kg) at a maintenance level of feeding (OMDm) was used as a predictor of methane production in this study and was determined using the equation as per Ramin and Huhtanen (2013). Digestibility of OM was estimated from the digestibility of DM when a study did not report OM digestibility using the equation derived from the data in this study. Since few variables were not available across all observations in the data set, the number of observations used for development of prediction equations varied between dietary and response variables depending on the regressor variables available. Data reported in differing units of measurements were transformed to the same units. Some records were incomplete or not reported uniformly, which necessitated the calculations (for example, conversion of methane production in L/day or kcal/day to g/day (1 L = 0.714 g methane; 1 L methane = 39.75 kJ energy), energy intake in kcal to MJ (1 kcal = 4.184 kJ), total digestible nutrients (TDN) value to ME (1 kg TDN = 3.6 MJ ME), etc. using standard conversion factors) from the reported data. Whenever possible, missing chemical composition of the diets was calculated from publications included in this dataset with similar ingredients. When a study did not report all possible outcomes and it was not possible to calculate from the reported data, missing variables were considered as missing data.
2.2 Statistical analysis
2.2.1 Linear and binomial model
Statistical analysis procedure used for prediction of methane production from this database was described elsewhere (Patra 2010, 2011). In brief, since studies represented random samples of larger population of studies, methane prediction equations were developed taking into account of the random effect of the study (St-Pierre 2001), using PROC MIXED (SAS 2001) with the following model:
Where:
Y ij = the predicted methane production at the level j of the independent variable X in the study i; B0 = the intercept across all studies (fixed effect); B1 and B2 = the linear and quadratic regressing coefficient of Y on X, respectively, across all studies (fixed effect); X ij = the value j of the variable X in study i; s i = the random study effect on the intercept; b i = the random study effect on regression coefficient of X; and e ij = the unexplained residual error.
The observed methane production data were weighted by the number of animals in each study to take into consideration of unequal variance among studies. The slopes and intercepts by study were included as random effects, and an unstructured variance-covariance matrix or a variance component of variance-covariance structure (when a random covariance component was not significant and a model failed to converge) was performed at the random part of the model (St-Pierre 2001). If random covariance or random slope and squared term of predictors were not significant (P > 0.10), they were removed from the models. All predictors of methane outputs and their quadratic term were further used to develop multiple regression equations employing the backward elimination multiple regression procedure following the algorithm reported by Oldick et al. (1999) and Patra (2010):
where:Y ij = the predicted methane production at the level j of the independent variables X1, X 2… Xn in the study i; B0 = the intercept across all studies (fixed effect); Bl1, Bl2..Bln = the linear and Bq1, Bq2..Bqn = quadratic regression coefficients of Y on X variables, respectively, across all studies (fixed effect); s i = the random effect of study i; b i = the random effect of study i on the regression coefficient of Y on X variables in study i; and e ij = the unexplained residual error. Two-way interactions were added when the coefficients of first order of Xs were significant (P < 0.05). To evaluate collinearities, a variance inflation factor (VIF) of less than 20 for every continuous independent variable tested was assumed. As the main objective of the study was to predict methane emission from dietary variables, high VIF considered here is not a serious problem (Geary 1963). Because model containing two-way interaction effect resulted in VIF of greater than 20, the interaction effect was not included in any final models. The best-fit equations of multiple regression equations that further improved the relationship obtained from simple linear or polynomial regression are presented. All statistical computations were carried out using the PROC MIXED and PROC CORR procedures of the SAS (2001) software system.
2.2.2 Nonlinear models
Since DM intake as sole independent variable predicted methane emission with highest degree of determination in the linear model, the nonlinear models were developed using DM intake as a determinant for prediction of methane outputs, if prediction ability of the equations could be further improved. The non-linear models employed the relationships exhibiting diminishing returns (monomolecular), sigmoidal (Gompertz), and exponential behaviors. The PROC NLMIXED of SAS was used to parameterize the non-linear functions with a little modification of the equation used by Schulin-Zeuthen et al. (2007) and the exponential model in the following forms:
- Monomolecular::
-
Y = a − (a + b) × exp(−c × x)
- Mitscherlich::
-
Y = a × (1 − exp(−c × x))
- Gompertz::
-
Y = b × exp((1 − exp(−c × x) × ln(a + 2b)/b) − 2b)
- Exponential::
-
Y = b × exp(c × x)
- Power::
-
Y = b × x c
Where Y represents the predicted methane production, the parameters a and b represent the upper asymptote and Y intercept of the nonlinear models, respectively, and c determines the shape of the response curve in the nonlinear functions. Study including parameters a, b, and c was considered random in the models (Schinckel and Craig 2002; Schulin-Zeuthen et al. 2007). The lowest value of Bayesian information criteria (BIC, a measure of regression fit) and biological relevance of the parameters estimated was considered to find out the optimum non-linear models.
2.3 Model evaluation
Predictive abilities of a range of existing models [Kriss (1930), Axelsson (1949), Mills et al. (2003), IPCC (2006) tier II, Ellis et al. (2007), Yan et al. (2009) and Ramin and Huhtanen (2012, 2013)] that were developed for dairy and beef cattle in temperate breeds and feeding situations were compared using inputs from this databases (Table 1). These equations were selected for comparison because they were commonly evaluated in different studies, and their input variables were available from this compiled database. Equations developed in this study and extant equations were compared using mean square prediction error (MSPE), square root of MSPE (RMSPE) expressed as a percentage of the observed mean (Theil 1966), and coefficient of determination (R 2) (Draper and Smith 1998). The MSPE value was calculated as:
where O i is the observed value for the ith observation, P i is the predicted value for the ith observation, and n is the number of observations. The RMSPE value, which provides an estimate of the overall prediction error, was expressed as a proportion (RMSPE divided by the observed mean) of the observed mean so that comparisons of RMSPE (%) values can be made among equations with different predicted means and so that deviation from observed values can be evaluated. The MSPE value was decomposed into mean bias or error in central tendency (ECT), slope bias or error due to regression (ER), and random or error due to disturbance (ED). These 3 fractions were calculated as follows (Bibby and Toutenburg 1977):
and expressed as a percentage of MSPE. The entities \( \overline{P} \) and Ō are the averaged predicted and observed values, respectively, S P and S O are the standard deviations of the predicted and observed values, respectively, and r is the coefficient of correlation between predicted and observed values. The ECT values indicate how the average of predicted values deviates from the average of observed values. The ER values measure deviation of the least squares regression coefficient (r × SO/SP) from 1 (the value where the model is completely accurate). A large ER value indicates inadequacies in the ability of the model to predict the variable. The ED value represents the variation in observed values unexplained after the mean and the regression biases have been removed.
Concordance correlation coefficient (CCC), also called reproducibility index, was used to evaluate the precision and accuracy of predicted versus observed values for each model (Lin 1989). The CCC estimate represents as a product of two components. The first component is the correlation coefficient (r) that measures a precision (deviation of observations from the best fit line). Second component is the bias correction factor (Cb) that indicates how far the regression line deviates from the line of unity (accuracy). Another estimate (μ) measures location shift relative to the scale (difference of the means relative to the square root of the product of two standard deviations), where a negative value indicates over-prediction and a positive value indicates under-prediction of observed values by the model.
The prediction biases of the equations developed in this study (one simple, two multiple, and one non-linear models) and the extant models (IPCC 2006; Ellis et al. 2007; Ramin and Huhtanen 2012; Patra 2014b), which predicted methane production with greater accuracy and precision, were further evaluated in the form of residual plots. The residuals (observed−predicted) were plotted against predicted values. The independent variable predicted methane outputs was centered around the mean predicted value before the residuals were regressed on the predicted value as described by St-Pierre (2003), and mean centered bias and biases at the minimum and maximum values were determined as described by St-Pierre (2003).
3 Results
3.1 Description of dataset
A description of the dietary and animal characteristics included in this database such as BW, nutrient and energy intake, feed digestibility, and methane production is provided in Table 2. The concentrations of crude protein (CP) and neutral detergent fiber (NDF) ranged from 24 to 235 (mean values of 117) g/kg and from 192 to 821 (mean values of 567) g/kg DM, which signified that quality of diets varied widely in this database. The mean concentrations of CP and NDF in the diets suggested that low-to-medium quality diets were mainly included in this dataset. Though the roughage proportions in the diets ranged from 0 to 1000 g/kg, mean forage proportion was high (753 g/kg), indicating predominantly forage-based diets were included in the study. These dietary situations are typical in the tropical parts of the world. The wide range of digestibilities of DM, NDF, CP, and ether extract (EE) in this study suggested that digestibilities varied considerably depending upon dietary chemical composition. The methane emissions expressed in terms of MJ/day, g/kg DM intake, and % of GE or digestible energy intake also ranged widely in the dataset.
3.2 Correlations between methane production and animal and dietary variables
With exception of EE intake (P = 0.30), methane production expressed as MJ/day was positively (P = 0.002 to <0.001) correlated (r = 0.44 to 0.83) with BW, FL, and intakes of all of the nutrients with highest correlation observed for DM and GE intake (Table 3). However, methane production expressed as g/kg DM intake or % of GE intake negatively correlated (P = 0.02) with EE intake and FL only and tended to positively correlate (P = 0.07 to 0.08) with non-fibrous carbohydrate (NFC) only. Concentration of lignin in diets had a negative (P = 0.04) relationship, and concentrations of CP and acid detergent fiber (ADF) had a tendency of positive and negative relationship, respectively, with daily methane emission expressed as MJ/day; however, the relationships were poor. In contrast, methane outputs expressed as g/kg DM intake or % of GE intake correlated positively with NDF (P < 0.01) and ADF (P = 0.02) concentrations negatively correlated with EE concentration (P < 0.01) and tended (P = 0.07 to 0.09) to correlate with CP concentration; however, correlations between methane emission as g/kg DM intake or % of GE intake and concentrations of lignin and NFC were not significant (P > 0.10). Roughage proportion in the diets correlated (P < 0.001) positively with methane production when expressed as g/kg DM intake or % of GE intake, but not with daily methane outputs.
3.3 Prediction equations for methane production
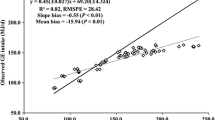
Prediction models for enteric methane emission were developed using BW, intakes of DM, nutrients (NDF, ADF, CP, EE, and NFC) and energy (GE, ME and GE), and dietary composition of nutrients (CP, EE, NDF, ADF, lignin, and NFC) as predictors (Table 4). The BW of the animals predicted methane production with R 2 = 0.43. With the exception of EE and CP, intake of all nutrients significantly predicted methane outputs as a single predictor. The predictions of methane output were high for intake of DM (R 2 = 0.70; Fig. 1), GE (R 2 = 0.69), DE (R 2 = 0.67), and ME (R2 = 0.66), but were low for intakes of NDF (R 2 = 0.47), ADF (R 2 = 0.37), and NFC (R 2 = 0.20). Generally, methane prediction was better at low levels of methane emission (Fig. 1). Among the nutrients evaluated as methane predictors, only fiber (NDF and ADF and lignin) concentrations predicted methane emissions, but the prediction were very low (R 2 = 0.003 to 0.05). Even the multiple regression model using nutrients (NDF and ADF as predictors) had low predictive ability (R 2 = 0.12). Methane emission was not predicted by the concentrations of CP, NFC, and EE in this database. Multiple regression models containing intakes of DM and NDF (R 2 = 0.74), DM and ADF (R 2 = 0.80), DM intake and FL (R 2 = 0.79), as two independent variables in the models improved methane prediction compared with a single predictor, and inclusion of each of these variables had a significant effect on the relationship. Among the multiple regression equations with three independent variables, the model containing DM intake, ADF intake, and FL improved the model fit to a little extent (R 2 = 0.84) compared with the models containing DM intake, FL, and OM digestibility (R 2 = 0.80), and DM intake, FL, and GE digestibility (R 2 = 0.80). Any other independent variables and interaction terms in the multiple regression equations did not further improve prediction of methane emission.

Predicted versus observed methane production (MJ/day), where methane is predicted by Eq. 1 (top left), Eq. 12 (top right), Eq. 17 (bottom left), and Mitscherlich model (bottom right)
Dry matter intake was used to predict methane emission using non-linear models as DM intake had most predictive ability as a single factor of methane prediction. However, exponential growth (R 2 = 0.64) and Gompertz (R 2 = 0.66) models did not improve prediction of methane outputs further compared with the linear model using DM intake (R 2 = 0.70) as a single predictor. Monomolecular (R 2 = 0.72) and power model (R 2 = 0.72) marginally and Mitscherlich model (R 2 = 0.82) greatly improved methane prediction with higher R 2 and lower RMSE values compared with the linear model.
3.4 Comparison of models
Analyses of RMSPE and CCC of the developed methane prediction equations (Table 5) suggested that equation based on DM intake was the best predictor of methane production among the simple models considering smaller RMSPE (31.0 %, of which 92 % due to random error), greater precision (CCC values = 0.80), and accuracy (Cb = 0.94), followed by GE intake (RMSPE% = 31.9 with 92.8 % error from random sources and CCC and Cb values of 0.79 and 0.93, respectively). The equations based on nutrient composition had greater RMSPE along with lower precision and accuracy than the other models. Among the multiple regression equations, models containing intakes of DM and ADF and FL resulted in the lowest RMSPE values (RMSPE% = 22.2 %) with random error sources of 99.6 % and greater precision (CCC = 0.92) and accuracy (Cb = 0.99). Among the non-linear models, Mitscherlich model marginally improved the prediction of methane in terms of RMSPE (30.3 % with 97.6 % from random error and lower regression bias of 2.03 %), precision (CCC = 0.82), and accuracy (Cb = 0.97) compared with simple linear and non-linear models with a single variable as a predictor. The mean biases (difference between predicted and actual data) were low for all models except for model based on lignin concentration, and concentrations of NDF and ADF. As number of treatments varied among the variables depending upon the available data, precision and accuracy among the models may impose some biases for the comparison of the models.
With the exception of the models of Patra (2014b), methane emissions were over-predicted by all extant models as indicated by higher negative μ values. The extant models generally had greater RMSPE values and larger mean biases. Among the extant models, the mean biases were lower for IPCC (2006), Ellis et al. (2007), and Patra (2014b) models, and the largest mean bias (80 % of RMSPE) was noted for Mills et al. (2003) linear model. The models of IPCC (2006), Patra (2014b), and Ellis et al. (2007) had also better precision (CCC) and accuracy (Cb) among the extant models.
There were no significant mean and linear biases (P > 0.05) for Eq. 17 and Mitscherlich model (Fig. 2). Although the slope biases, but not mean biases, of Eqs. 1 and 13 were significant statistically, they resulted in maximum biases of 2.30 and 2.57 MJ/day over the full range of predicted values, respectively. In contrast, the mean and linear biases (except for Ramin and Huhtanen 2012) of four best extant equations evaluated in this database were significant (P < 0.05) and resulted in a maximum biases of 2.15, 3.02, 2.30, and 2.72 MJ/day over the full range of predicted values for models of IPCC (2006), Ellis et al. (2007), Ramin and Huhtanen (2012), and Patra (2014b), respectively (Fig. 3).

Plot of observed minus predicted methane production (residual) versus predicted methane production from cattle. The independent variable (predicted methane production) was centered around the mean predicted value before the residuals were regressed on the predicted values, where for Eq. 1, Y = 0.079 (±0.21; P = 0.70) + 0.22 (±0.065; P = 0.001) (X − 7.97), R 2 = 0.08, P = 0.001 (top left); for Eq. 13, Y = 0.041 (±0.20; P = 0.84) + 0.24 (±0.059; P = 0.001) (X − 7.80), R 2 = 0.12, P = 0.001 (top right); for Eq. 17, Y = 0.061 (±0.14; P = 0.50) + 0.023 (±0.064; P = 0.96) (X − 7.97), R 2 = 0.002, P = 0.97 (bottom left); for Mitscherlich model, Y = 0.151 (±0.21; P = 0.51) + 0.098 (±0.059; P = 0.11) (X − 7.89), R 2 = 0.02, P = 0.11 (bottom right)

Plot of observed minus predicted methane production (residual) versus predicted methane production from cattle. The independent variable (predicted methane production) was centered around the mean predicted value before the residuals were regressed on the predicted values, where for equation of IPCC (2006), Y = − 0.73 (±0.22; P = 0.001) − 0.10 (±0.050; P = 0.04) (X − 8.77), R 2 = 0.03, P = 0.04 (top left); for linear equation 1 of Ellis et al. (2007), Y = −0.82 (±0.21; P < 0.001) + 0.45 (±0.077; P < 0.001) (X − 8.87), R 2 = 0.20, P < 0.001 (top right); for linear equation 1 of Ramin and Huhtanen (2012), Y = −1.95 (±0.21; P < 0.001) − 0.042 (±0.052; P = 0.42) (X − 10.0), R 2 = 0.005; P = 0.42 (bottom left); for equation of Patra (2014b), Y = 0.65 (±0.21; P = 0.002) + 0.23 (±0.066; P = 0.001) (X − 7.39), R 2 = 0.08, P = 0.001 (bottom right)
4 Discussion
The purpose of this study was to develop statistical models of enteric methane production in cattle of tropical feeding systems and to assess the dietary composition affecting methane production. Majority of the dietary treatment means included in the database were from India (n = 54) and Brazil (n = 55). It is imperative to state that major proportion of cattle populations is centered in these countries, and cattle populations are growing in these regions. A sizeable proportion of cattle population is also located in African countries, and cattle population in Africa is expected to grow faster in coming decades (Gerber et al. 2013). However, the literature on in vivo methane production in cattle in Africa is scanty. Thus, the models developed in this study may be less suited to predict methane production and understand feeding strategies in African countries.
The average methane production from cattle of tropical climate was 1.04 MJ/kg DM intake or 5.84 % of GE intake in this study. The methane emission ranged from 1.12 to 1.49 MJ/kg DM (Ellis et al. 2007; Yan et al. 2009) or 6.37 to 10.1 % of GE intake (Wilkerson et al. 1995; Yan et al. 2009) reported for dairy and beef cattle from temperate situations. It appears that methane production per unit of feed intake is lower for tropical cattle production systems than temperate cattle production systems, which is likely due to the lower quality diets (containing low concentration of CP and high concentration of NDF) fed to the cattle in the tropics compared with the diets offered to cattle in temperate countries (Van Soest 1994). Methane is produced during fermentation of feeds in the rumen production. Digestibility of forages and crop residues in tropical countries is low (Van Soest 1994), and consequently, low quality feeds in tropical countries may result in lower methane production per unit of feed intake. For example, Kurihara et al. (1999) studied two tropical grasses, i.e., mature Angleton grass (Dicanthium aristatum) hay with NDF digestibility of 55 % and immature Rhodes grass (Chloris gayana) hay with NDF digestibility of 69 % for methane production in tropical breed of cattle. Methane production in cattle fed on Angleton grass was lower than in cattle fed on Rhodes grass hay (113 versus 257 g/day and 31.6 versus 36.3 g/kg DM, respectively; Kurihara et al. 1999).
Various models developed in this study clearly demonstrated that intakes of nutrients particularly DM or GE were the stronger determinant of methane production than nutrient composition. There was a strong relationship between methane production and intake of DM or GE. The prediction of methane production was better at low levels of methane production suggesting that other physiological and microbiological factors such as rumen volume and fermentation characteristics in addition to intakes of nutrients may interplay in methane production in the rumen at high level of methane production (Hegarty 2004). A number of studies also reported that feed intake (DM or energy) was a principal determinant for prediction equations of methane emissions in dairy and beef cattle (e.g., Mills et al. 2003; Ramin and Huhtanen 2013; Yan et al. 2009). In this study, R 2 values of 0.69 to 0.70 in the relationship between methane and DM or GE intake were moderately high. Moderate R 2 values of 0.68 with DM intake or 0.70 with GE intake were reported in UK feeding conditions (Yan et al. 2009). However, the prediction equations using DM intake or ME intake as primary predictors of methane outputs had low R 2 values (0.44 or 0.36) in the study with beef cattle (Ellis et al. 2007). Yan et al. (2000) even reported a R 2 value of 0.85 for the methane prediction equation based on GE intake. These differences among the studies may result from the wider variations of the ingredient and chemical composition of diets and animal characteristics in each database. The ME intake and DE intake are expected to be better determinants of methane outputs than DM intake as the former account for methane production within its derivation (Mills et al. 2003). In the study of Mills et al. (2003), ME intake and DE intake were better predictors of methane production than DM intake and GE intake. However, ME and DE intake predicted methane outputs with less precision compared with DM intake in this study. This may be attributed to the inclusion of calculated ME and DE intake values for many studies included in this dataset instead of direct measured values, thus imposing more errors in ME and DE intake values. This result suggests for better characterization of feeds of tropical regions of the countries instead of using calculated nutritive values of feeds. Thus, extensive research activities are needed to characterize tropical feeds for methane production in the context of food security, mitigation of methane production, and climate change adaptation in tropical developing countries. Ellis et al. (2007) also noted a lower precision and accuracy in the prediction of methane production using ME intake than DM intake in dairy (R 2 = 0.64 versus 0.53) and beef cattle (R 2 = 0.44 versus 0.36) datasets.
It is imperative that the quadratic term of intakes of DM, GE, DE, or ME as a single determinant was not significant (P > 0.05) in predicting methane emission in this study. However, Ramin and Huhtanen (2013) found that the quadratic model containing DM intake as a single predictor improved the goodness of fit compared with the linear model. In contrast, the quadratic term of NDF intake or ADF intake as a single methane predictor in this study, which was negatively related, was significant (P < 0.05). This is expected because particulate passage rate increases to a greater extent with increasing fiber intake compared with other nutrients resulting in lower fermentation of feeds in the rumen and consequently lower methane production. However, many models developed earlier based on fiber intake did not include quadratic term of the fiber fractions (e.g., Mills et al. 2003; Ellis et al. 2007, 2009).
Intake of DM or GE is a key factor in most of the enteric methane emission prediction equation, but is difficult to accurately determine on farms, particularly in grazing conditions. Thus, chemical composition of diets and BW of animals were used for development of prediction equations, as these equations may be useful in situations where intake data may not be available. The BW of animals resulted in a reasonable degree of prediction of methane production. However, concentrations of NDF, ADF, and lignin as a single determinant had lower predictability (R 2 = 0.003 to 0.05) of methane outputs in the database, and goodness of fit of these equations was very low. Even the multiple regression equation based on dietary composition had low predictability (R 2 = 0.12). However, nutrient composition of diets predicted methane outputs with relatively high R 2 values in the studies of Mills et al. (2003) (R 2 = 0.24 to 0.35) and Ellis et al. (2007) (R 2 = 0.01 to 0.35). The low relationship between nutrient composition and methane production in this study is likely due to greater variations of dietary nutrient composition and nutritive values in tropical climates compared with the temperate climates (Van Soest 1994). Tropical feeds have also low predictability of nutritive values from fiber components (Van Soest 1994). Dietary EE was negatively correlated with methane production, and the prediction model for methane production included EE as a predictor when the database included the studies with dietary fat supplementation (e.g., Grainger and Beauchemin 2011; Moate et al. 2011; Patra 2013). However, EE in this study was not associated with methane production, which was presumably due to the presence of low concentrations of EE (29 ± 22 g/kg DM) in the diets. Ellis et al. (2007) reported that the multiple regression equation containing DM intake and EE intake improved the prediction of methane production in the beef database, but not in the dairy database.
Multiple regression equations were presented when they improved the prediction compared with simple regression equations. In simple regression equations, the quadratic term was not significant for DM, but the multiple regression equations contained significant positive quadratic effect of DM and negative quadratic effect of NDF or ADF. This might suggest that methane production is influenced by DM intake as well as fiber intake. The FL was negatively related in the multiple regression equations, suggesting that FL influences methane production mediated probably through changes in passage rate and rumen digestion of feeds (Ramin and Huhtanen 2013). The most improved multiple regression equations developed by Ellis et al. (2007) included ME intake, ADF intake, and lignin intake as determinants with R 2 = 0.85 for the beef dataset, and DM intake, NDF intake, and lignin intake with R 2 = 0.71 for the combined beef and dairy dataset. In the present database, lignin had no significant contribution in the prediction of methane in any multiple regression models.
Methane production in the rumen depends upon dietary factors, rumen functions, and fermentation dynamics and may not follow a linear trend over a wide range of values. Therefore, nonlinear regression models were also evaluated for prediction of methane emission using DM intake. Among the non-linear models, Mitscherlich model improved the relationship between methane production and DM intake compared with the simple linear models. Although Mills et al. (2003) noted that there was a minor difference in RMSPE percentage between the linear and non-linear models for the UK data, the benefits were evident for the American and Northern Ireland data for lactating cows. It is evident that the non-linear models of Ramin and Huhtanen (2012) and Mills et al. (2003) performed slightly better than the linear models of Ramin and Huhtanen (2013) and Mills et al. (2003) when they were challenged with this database. Thus, the non-linear models may be better for predicting methane production in a wide range of intake and dietary variables, which has also been suggested by Mills et al. (2003). Nonetheless, the prediction of methane production should be made with caution when dependent variables are outside the range of this database because few models have intercept values that are not biologically relevant.
Several extant regression equations were used to validate the predictability of methane production in this database. The newly developed models generally performed better than the extant models as these equations had lower RMSPE values compared with the extant models. Among the extant models, the equations of IPCC (2006) and Patra (2014b) had better goodness of fit in this database. This is because IPCC (2006) considered both tropical and temperate feeding situations; Patra (2014b) included most of the data from tropical countries of buffalo production system. The lower accuracy and precision of the extant models developed from cattle of North America and European situations compared with new equations developed in this study may be attributed to the animal type, geographical and dietary differences (King et al. 2011; Wright and Klieve 2011). Among extant equations, few simple DM intake models were quite good compared with multiple regression models developed in this study (e.g., Ellis et al. 2007; Ramin and Huhtanen 2013) when challenged in this database. This indicates that the parameters in those models derived using temperate cattle are unable to reflect the associations in tropical cattle. The diets in this study were of low to medium quality whereas the diets for cattle in North America and European countries were of medium to high quality. Besides, the breeds of the tropical countries are mainly of zebu types, which have low nutrient requirements due to low body weight and productivity. Preparation of inventory of methane emissions as per methodology of IPCC (2006) tier II requires country-specific methane conversion factors (Ym; methane production as a proportion of GE intake) for each categories of livestock, but many tropical countries, especially in Africa and Asia, do not have country-specific Ym because of lack of information of the methane energy loss as a percentage of GE intake for those countries. The enteric methane production based on the activity data of Patra (2012) and IPCC (2006) tier II methodologies, methane production from Indian cattle was 7736 Gg/year in 2007. However, methane production from Indian cattle using the same activity data and model 1 and model 3 was 5665 and 5703 Gg/year, respectively, in 2007, which was considerably (26.5 %) lower compared with the estimates based on the IPCC (2006) methane conversion factor. The models reported in this study should be considered for more accurately preparing the enteric methane emission inventories for cattle in the tropical countries.
5 Conclusions
Linear models developed based on DM intake or GE intake as a single predictor improved the prediction of methane production. The multiple regression equation based on intakes of NDF, ADF, and NFC improved the goodness of model fit and had better precision and accuracy than the linear models. Among the nonlinear models, Mitscherlich model performed better than simple linear models. The extant models developed for cattle in temperate production systems over-predicted methane emissions, when they were challenged in this database, and most of these extant equations except IPCC (2006) had low precision and accuracy for the prediction of methane outputs from cattle of tropical feeding situations. A better estimate of enteric methane emissions by livestock species using IPCC (2006) tier II and III methodologies requires different activity data such as animal numbers in different categories and ages of each animal species, milk production, growth rate, dietary composition, feed availability, energy/DM requirements, and country-specific Ym and methane emission (i.e., per animal annually) factors. The IPCC (2006) developed methodologies to estimate enteric methane emissions with the use of Ym. However, Ym does not directly represent variations in methane emissions determined by the ruminal fermentation of distinct carbohydrates and feeding levels. Thus, the usefulness of Ym based models in predicting enteric methane emissions and evaluating dietary methane mitigation options is limited (Moraes et al. 2014). Furthermore, the low predictive ability of the Ym approach may introduce considerable inaccuracy in preparation of enteric methane emission inventory (Ellis et al. 2010; Patra 2014b). The equations developed in this study will be valuable to estimate country-specific Ym and methane emission factors using feed intake and diet characteristics, which will be useful for more accurately preparing enteric methane emission inventory data from cattle in tropical regions of the countries instead of using default enteric methane emissions factors of IPCC (2006) when other activity data are available. Moreover, this study specifies a better understanding of dietary factors influencing methane production in cattle for tropical production systems. Nonetheless, these newly developed models should be evaluated on an external database for testing the goodness of fit of the prediction equations of methane production from cattle in the tropics.
References
Axelsson J (1949) The amount of produced methane energy in the European metabolic experiments with adult cattle. Kungl Lantbrukshogskolans Ann [Ann R Agric Coll Sweden] 16:404–419
Bairagi B, Mohini M (2003) Methane production and nutrient utilization at low levels of oil in diets of steers. Indian J Anim Prod Manage 19:84–89
Benchaar C, Pomar C, Chiquette J (2001) Evaluation of dietary strategies to reduce methane production in ruminants: a modelling approach. Can J Anim Sci 81:563–574
Bhar R, Lal M, Khan MY (1999) Nutrient utilization in crossbred dairy cows under economical feeding regime. Indian J Anim Nutr 16:358–361
Bibby J, Toutenburg H (1977) Prediction and improved estimation in linear models. Wiley, London
Blaxter KL, Clapperton JL (1965) Prediction of the amount of methane produced by ruminants. Br J Nutr 19:511–522
Bouchard K, Wittenberg KM, Legesse G, Krause DO, Khafipour E, Buckley KE, Ominski KH (2013) Comparison of feed intake, body weight gain, enteric methane emission and relative abundance of rumen microbes in steers fed sainfoin and lucerne silages under western Canadian conditions. Grass Forage Sci. doi:10.1111/gfs.12105
De D, Mohini M, Singh GP (2012) Influence of monensin enriched UMMB feeding on in vivo methane emission in crossbred calves fed on wheat straw and concentrate based diet. Indian J Anim Sci 82:640–644
Demarchi JJAA, Manella MQ, Lourenço AJ, Alleoni GF, Frighetto RTS, Primavesi O, Lima MA (2003) Preliminary results on methane emission by Nelore cattle in brazil grazing Brachiaria brizantha cv. Marandu. In: Proceedings of the 3rd international methane and nitrous oxide mitigation conference, Beijing, China, 17–21 November 17–21 2003
Draper NR, Smith H (1998) Applied regression analysis, 3rd edn. Wiley, New York
Ellis JL, Kebreab E, Odongo NE, McBride BW, Okine EK, France J (2007) Prediction of methane production from dairy and beef cattle. J Dairy Sci 90:3456–3467
Ellis JL, Kebreab E, Odongo NE, Beauchemin K, Mcginn S, Nkrumah JD, Moore SS, Christopherson R, Murdoch GK, McBride BW, Okine EK, France J (2009) Modeling methane production from beef cattle using linear and nonlinear approaches. J Anim Sci 87:1334–1345
Ellis JL, Bannink A, France J, Kebreab E, Dijkstra J (2010) Evaluation of enteric methane prediction equations for dairy cows used in whole farm models. Glob Chang Biol 16:3246–3256
Geary RC (1963) Some results about relations between stochastic variables: a discussion document. Rev Int Stat Inst 31:163–181
Gerber PJ, Steinfeld H, Henderson B, Mottet A, Opio C, Dijkman J, Falcucci A, Tempio G (2013) Tackling climate change through livestock – a global assessment of emissions and mitigation opportunities. Food and Agriculture Organization of the United Nations (FAO), Rome
Ghosh A, Murarilal, Haque N, Khan MY (2001) Determination of energy requirement for maintenance of adult nonlactating crossbred cows. Indian J Anim Sci 71:950–954
Girdhar N, Khan MY, Lal M (1995) Energy balance studies in Holstein Friesian and HF x Hariana calves fed on wheat straw based ration. Indian J Anim Nutr 12:13–18
Grainger C, Beauchemin KA (2011) Can enteric methane emissions from ruminants be lowered without lowering their production? Anim Feed Sci Technol 166–167:308–320
Hammond KJ, Burke JL, Koolarrd JP, Muetzel S, Pinares-Patino CS, Waghorn GC (2013) Effects of feed intake on enteric methane emissions from sheep fed fresh white clover (Trifolium repens) and perennial ryegrass (Lolium perenne) forages. Anim Feed Sci Technol 179:121–132
Haque N, Saraswat ML, Sahoo A (2001) Methane production and energy balance in crossbred male calves fed on rations containing different ratios of green sorghum and wheat straw. Indian J Anim Sci 71:797–799
Hegarty RS (2004) Genotype differences and their impact on digestive tract function of ruminants: a review. Aust J Exp Agric 44:458–467
Hernandez-Sanabria E, Goonewardene LA, Wang Z, Zhou M, Moore SS, Guan LL (2013) Influence of sire breed on the interplay among rumen microbial populations inhabiting the rumen liquid of the progeny in beef cattle. PLoS ONE 8(3), e58461. doi:10.1371/journal.pone.0058461
Holter JB, Young AJ (1992) Methane production in dry and lactating Holstein cows. J Dairy Sci 75:2165–2175
Hristov AN, Oh J, Lee C, Meinen R, Montes F, Ott T, Firkins J, Rotz A, Dell C, Adesogan A, Yang W, Tricarico J, Kebreab E, Waghorn G, Dijkstra J, Oosting S (2013) Mitigation of greenhouse gas emissions in livestock production—a review of technical options for non-CO2 emissions, vol 177, FAO animal production and health. FAO, Rome
Hulshof RBA, Berndt A, Gerrits WJJ, Dijkstra J, van Zijderveld SM, Newbold JR, Perdok HB (2012) Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane-based diets. J Anim Sci 90:2317–2323
IPCC (2006) 2006 IPCC guidelines for national greenhouse gas inventories. IGES, Kanagawa, http://www.ipcc-nggip.iges.or.jp
IPCC (2013) Climate change 2013: the physical science basis. In: Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge and New York
Jain P, Mohini M, Singhal KK, Tyagi AK (2011) Effect of herbal mixture supplementation on methane emission and milk production in cattle. Indian J Anim Nutr 28:377–384
Kannan A, Garg MR (2009) Effect of ration balancing on methane emission reduction in lactating animals under field conditions. Indian J Dairy Sci 62:292–296
Kannan A, Garg MR, Kumar BVM (2011) Effect of ration balancing on milk production, microbial protein synthesis and methane emission in crossbred cows under field conditions in Chittoor district of Andhra Pradesh. Indian J Anim Nutr 28:117–123
Kannan A, Garg MR, Singh P (2010) Effect of ration balancing on methane emission and milk production in lactating animals under field conditions in Rae Bareli district of Uttar Pradesh. Indian J Anim Nutr 27:103–108
Kearl LC (1982) Nutrient requirements of ruminants in developing countries. International Feedstuffs Institute, Utah Agriculture Experiment Station, Utah State University, Logan
Kebreab E, Clark K, Wagner-Riddle C, France J (2006) Methane and nitrous oxide emissions from Canadian animal agriculture: a review. Can J Anim Sci 86:135–158
Kebreab E, Johnson KA, Archibeque SL, Pape D, Wirth T (2008) Model for estimating enteric methane emissions from United States dairy and feedlot cattle. J Anim Sci 86:2738–2748
Kennedy APM, Charmley E (2012) Methane yields from Brahman cattle fed tropical grasses and legumes. Anim Prod Sci 52:225–239
King EE, Smith RP, St-Pierre B, Wright AD (2011) Differences in the rumen methanogen populations of lactating Jersey and Holstein dairy cows under the same diet regimen. Appl Environ Microbiol 77:5682–5687
Kriss M (1930) Quantitative relations of the dry matter of the food consumed, the heat production, the gaseous outgo, and the insensible loss in body weight of cattle. J Agric Res 40:283–295
Kurihara M, Magner T, Hunter RA, McCrabb GJ (1999) Methane production and energy partition of cattle in the tropics. Br J Nutr 81:227–234
Lal M, Khan MY, Kishan J, Katiyar RC, Joshi DC (1987) Comparative nutrient utilization by Holstein Fresian, crossbred cattle and buffaloes fed on wheat straw based ration. Indian J Anim Nutr 4:177–180
Lin LIK (1989) A concordance correlation coefficient to evaluate reproducibility. Biometrics 45:255–268
Malik PK, Singal KK (2008) Influence of lucerne fodder supplementation on enteric methane emission in crossbred calves. Indian J Anim Sci 78:293–297
Mills JAN, Dijkstra J, Bannink A, Cammell SB, Kebreab E, France J (2001) A mechanistic model of whole-tract digestion and methanogenesis in the lactating dairy cow: model development, evaluation, and application. J Anim Sci 79:1584–1597
Mills JAN, Kebreab E, Yates CW, Crompton LA, Cammell SB, Dhanoa MS, Agnew RE, France J (2003) Alternative approaches to predicting methane emissions from dairy cows. J Anim Sci 81:3143–3150
Moate PJ, Williams SRO, Grainger C, Hannah MC, Ponnampalam EN, Eckard RJ (2011) Influence of cold-pressed canola, brewers grains and hominy meal as dietary supplements suitable for reducing enteric methane emissions from lactating dairy cows. Anim Feed Sci Technol 166–167:254–264
Moe PW, Tyrrell HF (1979) Methane production in dairy cows. J Dairy Sci 62:1583–1586
Mohini M, Mani V (2007) Effect of various ratios of wheat straw and concentrate mixture on digestibility and methane production in cattle. In: Bakshi MPS, Wadha M (eds) Proceedings of tropical animal nutrition conference, Karnal, India, 4–7 October 2007
Mohini M, Mani V, Singh GP (2007) Effect of different ratios of green and dry roughages on milk production and methane emission in cattle. Indian J Anim Sci 76:79–82
Mohini M, Singh GP (2010) Effect of supplementation of urea molassess mineral block on the milk yield and methane production in lactating cattle on different plane of nutrition. Indian J Anim Nutr 27:96–102
Mohini M, Singhal KK, Sirohi SK, Mohanta RK (2009) Methane emission from Sahiwal cows on dietary supplementation of fumeric acid as a feed additive. Indian J Anim Nutr 26:51–55
Moraes LE, Strathe AB, Fadel JG, Casper DP, Kebreab E (2014) Prediction of enteric methane emissions from cattle. Glob Chang Biol 20:2140–2148
Mupeta B, Livingston R, Westberg H, Hamma M (2000) Effect of graded levels of ram press sunflower cake on yield and composition of milk and methane production in crossbred (Bos taurus x Bos indicus) cows given ad libitum mixed maize straw, groundnut tops, sunflower heads basal feed. In: Proceedings of the second international methane mitigation conference, Novosibirsk, Russia, 8–23 June 2000.
Nascimento CFM, Demarchi JJAA, Berndt A, Rodrigues PHM (2008) Methane emissions by nellore beef cattle consuming Brachiaria brizantha with different stages of maturation. Aust J Exp Agric 48:lxiv–lxv
Neto GB, Berndt A, Nogueira JR, Demarchi JJAA, Nogueira JC (2009) Monensin and protein supplements on methane production and rumen protozoa in bovine fed low quality forage. South Afr J Anim Sci39 (Suppl 1):280–283
Oldick BS, Firkins JL, St-Pierre NR (1999) Estimation of microbial nitrogen flow to the duodenum of cattle based on dry matter intake and diet composition. J Dairy Sci 82:1497–1511
Oliveira SG, Berchielli TT, Pedreira MS, Primavesi O, Frighetto R, Lima MA (2007) Effect of tannin levels in sorghum silage and concentrate supplementation on apparent digestibility and methane emission in beef cattle. Anim Feed Sci Technol 135:236–248
Patra AK (2010) Aspects of nitrogen metabolism in sheep-fed mixed diets containing tree and shrub foliages. Br J Nutr 103:1319–1330
Patra AK (2011) Responses of feeding prebiotics on nutrient digestibility, faecal microbiota composition and short-chain fatty acid concentrations in dogs. Animal 5:1743–1750
Patra AK (2012) Estimation of methane and nitrous oxide emissions from Indian livestock. J Environ Monit 14:2673–2684
Patra AK (2013) The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: a meta-analysis. Livest Sci 155:244–254
Patra AK (2014a) Trends and projected estimates of GHG emissions from Indian livestock in comparisons with GHG emissions from world and developing countries. Asian-Australas J Anim Sci 27:592–599
Patra AK (2014b) Prediction of enteric methane emission from buffaloes using statistical models. Agric Ecosyst Environ 195:139–148
Patra AK, Lalhriatpuii M (2016) Development of statistical models for prediction of enteric methane emission from goats using nutrient composition and intake variables. Agric Ecosyst Environ 215:89-99
Pattanaik AK, Sastry VRB, Katiyar, RC, Lal M (2003) Influence of grain processing and dietary protein degradability on nitrogen metabolism, energy balance and methane production in young calves. Asian-Australas J Anim Sci 16:1443–1450
Pedreira MS, Berchielli TT, Primavesi O, Oliveira SG, Frighetto R, Lima MA (2012) Influence of different supplements and sugarcane (Saccharum officinarum L.) cultivars on intake, digestible variables and methane production of dairy heifers under tropical conditions. Trop Anim Health Prod 44:1773–1778
Pedreira MS, Oliveira SG, Primavesi O, Lima MA, Frighetto R, Berchielli TT (2013) Methane emissions and estimates of ruminal fermentation parameters in beef cattle fed different dietary concentrate levels. R Bras Zootec 42:592–598
Pedreira MS, Primavesi O, Lima MA, Frighetto R, Oliveira SG, Berchielli TT (2009) Ruminal methane emission by dairy cattle in southeast Brazil. Sci Agric (Piracicaba, Braz) 66:742–750
Perna Jr F, Marino CT, Pinedo LA, Cassiano ECO, Martins MF, Solórzano LAR, Meyer PM, Frighetto RTS, Berndt A, Rodrigues PHM (2013) Effect of feed additives on methane production as determined by the tracer technique SF6 in bovine. In: Proceedings of the 4th international conference on sustainable animal agriculture for developing countries (saadc2013), Lanzhou university, Lanzhou, China, 27–31 7uly 2013
Possenti RA, Franzolin R, Schammas EA, Demarchi JJAA, Frighetto RTS, Lima MA (2008) Efeitos de dietas contendo Leucaena leucocephala e Saccharomyces cerevisiae sobre a fermentação ruminal e a emissão de gás metano em bovines R Bras Zootec 37:1509–1516
Primavesi O, Frighetto RTS, Pedreira MS, Lima M A, Berchielli TT, Barbosa PF (2004) Metano entérico de bovinos leiteiros em condições tropicais brasileiras. Pesquisa Agropecuária Brasileira 39:277–283
Primavesi O, Frighetto R, Pedreira MS, Lima MA, Berchielli TT, Rodrigues AA (2003) Low-fiber sugarcane to improve meat production with less methane emission in tropical dry season. In: Proceedings of the 3rd international methane and nitrous oxide mitigation conference Beijing, China, 17–21 November 2003
Ramin M, Huhtanen P (2012) Development of non-linear models for predicting enteric methane production. Acta Agric Scand Sect A Anim Sci 62:254–258
Ramin M, Huhtanen P (2013) Development of equations for predicting methane emissions from ruminants. J Dairy Sci 96:2476–2493
Rejil MC, Mohini M, Singhal M (2008) Methane emission as affected by dietary supplementation of raw and roasted fenugreek seeds in cattle. Indian J Anim Nutr 25:37–42
SAS (2001) SAS User’s guide. Statistics. SAS Inst. Inc., Cary
Schinckel AP, Craig BA (2002) Evaluation of alternative nonlinear mixed effects models of swine growth. Prof Anim Sci 18:219–226
Schulin-Zeuthen M, Kebreab E, Gerrits WJ, Lopez S, Fan MZ, Dias RS, France J (2007) Meta-analysis of phosphorus balance data from growing pigs. J Anim Sci 85:1953–1961
Srivastava AK, Garg MR (2002) Use of sulphur hexafluoride tracer technique for measurement of methane emission from ruminants. Indian J Dairy Sci 55:36–39
St-Pierre NR (2001) Invited review: Integrating quantitative findings from multiple studies using mixed model methodology. J Dairy Sci 84:741–755
St-Pierre NR (2003) Reassessment of biases in predicted nitrogen flows to the duodenum by NRC 2001. J Dairy Sci 86:344–350
Theil H (1966) Applied economic forecasting. North-Holland Publishing Company, Amsterdam
Tomkins NW, McGinn SM, Turner DA, Charmley E (2011) Comparison of open-circuit respiration chambers with a micrometeorological method for determining methane emissions from beef cattle grazing a tropical pasture. Anim Feed Sci Technol 166–167:240– 247
Van Soest PJ (1994) Nutritional ecology of the ruminant. Cornell University Press, USA
Wilkerson VA, Casper DP, Mertens DR (1995) The prediction of methane production of Holstein cows by several equations. J Dairy Sci 78:2402–2414
Wright ADG, Klieve AV (2011) Does the complexity of the rumen microbial ecology preclude methane mitigation? Anim Feed Sci Technol 166–167:248–253
Yan T, Agnew RE, Gordon FJ, Porter MG (2000) Prediction of methane energy output in dairy and beef cattle offered grass silage-based diets. Livest Prod Sci 64:253–263
Yan T, Porter MG, Mayne CS (2009) Prediction of methane emission from beef cattle using data measured in indirect open-circuit respiration calorimeters. Animal 3:1455–1462
Zhou M, Hernandez-Sanabria E, Guan LL (2010) Characterization of variation in rumen methanogenic communities under different dietary and host feed efficiency conditions, as determined by PCR-denaturing gradient gel electrophoresis analysis. Appl Environ Microbiol 76:3776–3786
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
Author declares that there is no conflict of interest.
Rights and permissions
About this article

Cite this article
Patra, A.K. Prediction of enteric methane emission from cattle using linear and non-linear statistical models in tropical production systems. Mitig Adapt Strateg Glob Change 22, 629–650 (2017). https://doi.org/10.1007/s11027-015-9691-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11027-015-9691-7