
Abstract
Context
For depleted wildlife populations, understanding and effectively mitigating the direct and indirect impacts of roads can be a key component of recovery efforts. The ocelot is a federally endangered wild felid at risk of local extinction in south Texas, where vehicle collisions are their largest known source of mortality.
Objectives
To support road mortality mitigation, we evaluated mortality risk across the south Texas road network. We also assessed whether a sympatric carnivore, the bobcat, could serve as a surrogate to inform mortality risk.
Methods
We used several presence-only species distribution models to evaluate a suite of candidate landscape and road attributes and identify those associated with elevated road mortality risk using a 35-year wild felid road mortality dataset. Consensus predictions among models identified high-risk road segments for ocelots and bobcats. We compared predictions between species to determine overlap of high-risk areas.
Results
Areas where core ocelot habitat intersected roadways represented the greatest mortality risk, reflecting ocelot reliance on intact habitat and avoidance of degraded and fragmented landscapes. Bobcat road mortality risk had similar relationships to landscape features, with nearly all areas classified as high-risk for ocelots also identified as high-risk for bobcats.
Conclusions
Ocelots are vulnerable to road mortality in areas where roads intersect their intact scrub habitat. Bobcats, with appropriate data filters in place, are an acceptable surrogate for informing ocelot road mortality, particularly when ocelot mortality data are limited and for time-sensitive conservation decision-making. For ocelots and other fragmentation-intolerant carnivore species, wildlife underpasses and fencing in focused areas where roads and intact habitat intersect may be feasible options to reduce road mortality.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Humans have developed much of the terrestrial biosphere over the past century to support agricultural, industrial, and societal systems (Ellis 2011). The anthropogenic degradation of natural landscapes has negatively impacted wildlife populations worldwide (Crooks et al. 2017; Keinath et al. 2017), and led to loss of global biodiversity and potentially irreversible alteration of ecosystem function (Newbold et al. 2016). Roads are a ubiquitous element of anthropogenic infrastructure and recognition of their impacts has led to the emergent discipline of road ecology to understand the effects of roads on ecosystem structure and function, and the extensive ecological impacts of roads on wildlife (Forman 2003; van der Ree et al. 2015). Roads can be a direct and significant source of mortality through wildlife-vehicle collisions (Lima Santos et al. 2017), and can indirectly restrict animal movement and dispersal through habitat fragmentation and degradation (Bennett et al. 2013; Ceia-Hasse et al. 2017). These direct and indirect impacts create barriers (van Der Ree et al. 2011) which disrupt gene flow and demographic population structure (Proctor et al. 2005; Riley et al. 2006; Balkenhol and Waits 2009), and alter metapopulation dynamics (Ferreras et al. 2001), in some cases leading to local extinctions (Ceia-Hasse et al. 2018).
Mitigation measures to reduce the biological impacts of roads are a key component of wildlife recovery and conservation efforts (Roger et al. 2011). Road mortality mitigation, including wildlife crossing structures, fencing, culvert modification, signage, and speed restrictions can reduce wildlife-vehicle collisions and facilitate movement across roadways (van der Ree et al. 2007; Glista et al. 2009; Meisingset et al. 2014). To optimize the effectiveness of these measures, analyses of wildlife-vehicle collision occurrence can identify relevant landscape and road attributes associated with elevated mortality risk (Gunson and Teixeira 2015; Lin et al. 2019). For example, Ha and Shilling (2018) used road mortality data across four taxonomic groups in northern California to identify environmental and anthropogenic predictors of road mortality risk, highlighting the variability in hotspots of elevated road mortality risk among species groups. Landscape features such as land cover type and configuration (Červinka et al. 2015; Garrote et al. 2018), habitat fragmentation (Grilo et al. 2011), and topography (Seiler 2004) have been shown to influence road mortality risk. Road attributes including traffic volume (Van Langevelde and Jaarsma 2005; Barrientos and Bolonio 2009) and road curvature (Grilo et al. 2009) have also been linked to road mortality vulnerability.
Road mortality is often a larger threat for depleted and endangered populations already at risk of local extirpation. Wide-ranging carnivores are particularly vulnerable to the detrimental effects of roads due to large home range requirements, low densities, low recruitment rates, and sex-biased movement and dispersal (Crooks et al. 2011; Ceia-Hasse et al. 2017). In the U.S., the ocelot (Leopardus pardalis) is a critically endangered felid with populations found only in south Texas, where this species tenuously persists in increasingly road-impacted and fragmented habitats (Janečka et al. 2014). Ocelots in this region primarily inhabit dense Tamaulipan thornscrub (Harveson et al. 2004) and previous research indicates they are intolerant of more open or degraded habitat types and vulnerable to landscape fragmentation (Jackson et al. 2005; Horne et al. 2009; Janečka et al. 2016). Unfortunately, since the 1930s land conversion and road construction have reduced and fragmented thornscrub habitat by 91–95% (Jahrsdoerfer and Leslie 1988; Tremblay et al. 2005), and the lack of connectivity among subpopulations, exacerbated by roads, has led to inbreeding and genetic isolation (Janečka et al. 2011, 2014). For these populations, vehicle collisions are the largest known source of mortality (Haines et al. 2005a). Reduction of road mortality, in addition to restoration of Tamaulipan thornscrub habitat, reestablishment of population connectivity, and translocation of individuals from Mexico to bolster genetic diversity have been identified as the key recovery strategies needed to reduce the extinction risk of south Texas ocelots (Haines et al. 2005b; USFWS 2016).
One major challenge to understanding road mortality risk for endangered populations is data limitation, as data for depleted populations are often sparse (Breiner et al. 2015). Using a surrogate species can increase the information available, but it is critical that potential surrogates are vetted as reliable indicators for the target species (Teixeira et al. 2013). Bobcats are abundant, sympatric carnivores that occupy a comparable dietary niche (Booth-Binczik et al. 2013) and use the same Tamaulipan thornscrub habitats as ocelots, though bobcats are tolerant of a range of canopy cover structures while ocelots more strongly select for closed canopy thornscrub (Harveson et al. 2004; Horne et al. 2009; Leonard 2016). In other regions of the U.S., bobcats have been a model organism for understanding the effects of roads and habitat fragmentation on wide ranging carnivores (Crooks 2002; Litvaitis et al. 2015; Abouelezz et al. 2018). In south Texas, there is potential for bobcats to serve as informative surrogate species for ocelots, however the similarity between bobcat and ocelot road mortality risk has not been evaluated.
Given the need to mitigate ocelot road mortality, understanding spatial patterns of road mortality risk has been identified as an important element of ocelot population recovery efforts (USFWS 2016). Our analysis provides the first evaluation of ocelot road mortality risk across the south Texas road network and the potential relationship between ocelot mortality and landscape characteristics. To consider how landscape and road attributes are associated with ocelot road mortality risk, we used several presence-only species distribution models with a 35-year road mortality dataset (1983–2018) to identify high-risk road segments. We performed a similar analysis with south Texas bobcat road mortalities to ask how well data from this surrogate species can inform ocelot road mortality risk across the landscape. By identifying factors associated with ocelot and bobcat mortality risk, the results of these analyses can inform and support future mitigation planning to reduce ocelot road mortality and promote connectivity of imperiled ocelot populations.
Methods
Study area and focal species
Our study area focused on the road network in the three south Texas counties where ocelot populations are extant: Cameron, Kenedy, and Willacy (Fig. 1). Cameron and Willacy counties lie in the Lower Rio Grande Valley floodplain, and are largely developed for agriculture, ranching, oil and gas extraction, and urban infrastructure. Kenedy County is located to the north, and is primarily comprised of private ranchlands managed for native and exotic game species, with some remaining native habitat suitable for ocelots. U.S. Fish and Wildlife Service (USFWS) estimates that as of 2016, fewer than 100 ocelots remain in these three south Texas counties (USFWS 2016), distributed among small sub-populations that are isolated from each other by habitat loss and fragmentation (Janečka et al. 2011). Bobcats co-occur with ocelots in south Texas, but previous research has shown that, unlike ocelots, their populations are not genetically isolated in this region (Janečka et al. 2016).

Study area encompassing a filtered selection of the road network within the three southeasternmost counties in Texas: Cameron, Kenedy, and Willacy. The road network considered in analyses was limited to paved roads < 2 km in length
Ocelot and Bobcat mortality occurrence Data
Data processing and analyses were done in R 3.6.0 (R Core Team 2019) and ArcGIS 10.6 (ESRI 2018). To obtain road mortality occurrence locations, we collated all wild felid mortality records available to USFWS collected in south Texas from 1 January 1983 through 31 December 2018. USFWS records included mortalities of collared ocelots and bobcats as well as incidental mortality reports from opportunistically encountered unknown cats of both species. All records were reviewed to verify cause of death, sex, age, and location of mortality. Confidence in cause of mortality was categorized as verified, likely, and undetermined. We defined ‘verified’ as mortalities with available veterinary necropsy reports and reliable assessments by USFWS biologists whereas those we classified as ‘likely’ lacked necropsy confirmation but contained sufficient evidence from the condition of the carcass, location, and assessment from USFWS biologists to assign a cause of death. Records lacking the above information were classified as ‘undetermined’.
All verified and likely road mortality reports were revisited to confirm location accuracy. Road mortality locations not recorded with a GPS (e.g., location derived based on a verbal or written description) were assessed for specificity of location description. Road mortality locations where a description was exceedingly vague or subjective were not included in analysis. Bobcats are more widespread in south Texas than ocelots, therefore bobcat road mortalities considered in our analysis were constrained to mortalities located within a 5 km buffer of all ocelot road mortalities to limit bobcat road mortality risk assessment to a relevant range for comparison with ocelot road mortality risk.
To evaluate wild felid road mortality risk across the south Texas road network, we used a presence-only species distribution modeling (SDM) approach. Using SDMs to understand mortality risk across road networks is an established methodology that can elucidate complex road-wildlife interactions across landscapes (Garrote et al. 2018; Ha and Shilling 2018; Kantola et al. 2019; Lin et al. 2019). In presence-only SDM frameworks, occurrence locations are contrasted against background locations to determine probability of presence or habitat suitability across landscapes (Elith et al. 2011; Merow et al. 2013). To compare mortality locations to the surrounding landscape, we generated 500 random locations along the road network in our three focal counties to adequately represent the ranges of landscape and road attributes within our study area and serve as background locations in our modeling approach (Phillips et al. 2009, Online Appendix A). To account for sampling bias and potential spatial autocorrelation, we created a Gaussian kernel density surface using ocelot road mortality locations. This surface was rescaled between 0 and 1 and used as a sampling probability surface to spatially bias background point selection in regard to road mortality occurrence locations and represent a realistic extent for potential ocelot dispersal from currently occupied areas (Kramer-Schadt et al. 2013; Merow et al. 2013; Ha and Shilling 2018).
Characterizing landscape and road attributes
To relate wild felid road mortality risk to landscape and road attributes, we considered four habitat fragmentation metrics, one topographic metric, one hydrographic metric, one road density metric, and one road-habitat interaction metric as candidate predictors (Table 1). Spearman rank correlation among candidate predictors was limited to < 0.60 (Merow et al. 2013). We assigned all landscape and road attributes to 500 m segments of the south Texas road network within our focal counties. The road network considered was limited to paved roads greater than 2 km in length to limit the inclusion of local and private roads in our analysis. The mortality data used in our modeling approach were collected over an extended time period, therefore we were unable to account for all landscape and road attributes that may contribute to ocelot road mortality risk across the south Texas landscape. For example, vehicle-related attributes such as speed and traffic volume could not be included because data were not available for earlier road mortalities, but have been found to be significant in other studies of wildlife and road mortality risk (Visintin et al. 2016).
Habitat fragmentation metrics were calculated for scrub land cover data obtained from the National Land Cover Database (Homer et al. 2015; Jin et al. 2019; Yang et al. 2018). We considered scrub land cover representative of the ocelot’s preferred habitat type based on previous research on ocelot habitat use (Harveson et al. 2004). Using FRAGSTATS (McGarigal et al. 2012), we calculated metrics quantifying habitat fragmentation within a 5 km2 buffer area centered on each 500 m road segment including core area, aggregation, connectance, and the degree of scrub habitat edge contrast with developed land cover types (Table 1). This 5 km2 scale was chosen to represent an ecologically relevant perspective for space use and movement given previously reported home ranges for this species (Laack 1991; Haines et al. 2006; USFWS 2016). Because mortality events accumulated over an extended time-span (1983–2018), we matched mortalities with the closest temporal NLCD layer available to assign landscape characteristics. For ocelot road mortalities that occurred more than three years from the closest available NLCD layer, we used historic imagery available from the Texas Natural Resource Information System (TNRIS) to assess each focal area and visually confirm that land cover change between the year of the mortality and the year for the NLCD layer used was negligible.
Road density was quantified as kilometers of paved road within a 5 km2 area around each road segment using the Texas Department of Transportation Roadway Inventory layer (TxDOT 2019). To quantify topographic complexity, we calculated topographic position index (TPI) within a 0.25 km2 focal area using a National Elevation Dataset layer at 1-arc second resolution (USGS 2016a, b) and then took the standard deviation of TPI within a 5 km2 area around each road segment. To relate road segments to hydrography, we calculated Euclidean distance from each road segment to the nearest waterway using the U.S. Geological Survey National Hydrography Dataset stream layer (USGS 2016a, b), excluding artificial irrigation and canal waterways because this predictor was chosen to represent potential travel corridors along natural waterways with associated remnant habitat strips rather than water availability (Laack 1991). To quantify the interaction of scrub patches and road segments, we calculated the Euclidean distance of each road segment to the nearest patch of scrub intersecting the roadway within a buffered distance of 25 m on either side of the road segment. All input variables were scaled before analysis using the scale function in R based on recommendations for effective SDM model implementation (Royle et al. 2012).
Risk models
To assess mortality risk across the south Texas road network and ensure the consistency and stability of our results, we used three presence only SDMs: MaxEnt, MaxNet, and MaxLike. MaxEnt is a common presence-only SDM that uses multinomial logistic regression in a machine learning approach to determine the probability distribution of maximum entropy given the discrete study area and occurrence locations to derive a relative occurrence rate, which can approximate road mortality risk (Phillips et al. 2006; Elith et al. 2011; Kantola et al. 2019). MaxNet employs an infinitely-weighted logistic regression where background points are heavily weighted (W = 100) compared to presence points (W = 1) (Phillips et al. 2017). MaxLike directly estimates probability of occurrence using a logit-linear model (Royle et al. 2012). This multi-model framework allowed for comparison of results among the three approaches to evaluate consensus among input variables and resulting predictions and ensure that our conclusions were not an artifact of the individual limitations or behavior of one SDM (Marmion et al. 2009; Merow and Silander 2014). We ran risk models separately for ocelots and bobcats using verified and likely road mortalities with accurate location data from 1983–2018. MaxEnt was run through the dismo package (Hijmans et al. 2017) using only a linear feature class and a regularization multiplier of one to prevent over-fitting (Merow et al. 2013). We duplicated these parameters for MaxNet using the maxnet package (Phillips et al. 2017). MaxLike was run with the maxlike package (Royle et al. 2012) using the “BFGS” method (Merow and Silander 2014).
To calibrate risk models, we evaluated all landscape and road attributes using the three SDM approaches to assess consistency of relationships between predictors and road mortality risk. We removed predictors if their relationships (e.g., signs of the model coefficients) to road mortality risk were not consistent across all three approaches, because a lack of agreement across SDMs suggested poor predictor performance. We also ran preliminary univariate analyses across the three SDM approaches to develop a best consensus of predictor variable inputs for each felid species. In MaxEnt, we assessed percent contribution of a predictor, whether predictor response curves were static when other predictors were held at their mean, and univariate area under the receiver operator characteristic curve (AUC; Ha and Shilling 2018). In MaxNet, we assessed the magnitude of the model coefficients, which is an appropriate cursory assessment of variable importance for glmnet-based models if input variables are scaled. In MaxLike, we assessed significance of predictors at p < 0.05 in univariate analysis.
Once calibrated, we ran 50 iterations of each presence-only SDM using the best consensus model for each species, subsampling with replacement 80% of road mortality locations to train each model and reserving 20% of mortalities to test model performance in a cross-validation approach (Elith et al. 2011; Merow et al. 2013). We reported average coefficients from each SDM, and visualized predictor relationships for MaxEnt, MaxNet, and MaxLike using their packages’ respective predict functions across the full range of each predictor variable while holding all other predictors at their mean. To evaluate model performance, we used the testing data from each iteration to calculate the average area under the receiver operator characteristic curve (AUCtest) for each SDM approach. AUC values range from 0 to 1, where a value of 1 indicates a perfect fit to the data, and 0.5 represents chance performance (Phillips et al. 2006). AUC interpretation is not always straightforward for presence-only SDM frameworks (Fitzpatrick et al. 2013), therefore we also evaluated model performance using the continuous Boyce index (Bcont), which uses a moving window across predicted values for training and background data to create a predicted/expected curve used to calculate Bcont (Hirzel et al. 2006; Breiner et al. 2015). Values for Bcont range from − 1 to 1, where 1 indicates a perfect fit, 0 represents model performance no better than random, and values below 0 indicate that model performance is worse than random (Hirzel et al. 2006).
Risk maps
To develop road mortality risk maps across the south Texas road network for each felid species, we used each of the 50 training model iterations for each SDM approach to assign road mortality risk prediction values across the full south Texas road network. To compare predictions across SDM approaches, we converted model predictions to binary classifications of low and high risk. Therefore, high risk road segments are interpreted as segments with landscape and road attributes consistent with those associated with road mortality occurrences for each species (Garrote et al. 2018; Kantola et al. 2019). Optimal thresholds for binary classification were calculated with the full dataset of road mortality occurrence locations for each species using the maximum sum of sensitivity and specificity (maxSSS), which maximized the proportion of correctly predicted presences and correctly predicted absences (Liu et al. 2005). This threshold selection method is suitable for presence-only data and is robust to a range of known occurrence to background point ratios (Liu et al. 2013, 2016). We evaluated threshold selection performance with two additional metrics, sensitivity and the Symmetric Extremal Dependence Index (SEDI, Ferro and Stephenson 2011). Sensitivity describes the proportion of mortality occurrences accurately assigned a high-risk classification. SEDI is an evaluation metric for thresholded binary SDM predictions that is well-suited to presence-background data with limited prevalence (Wunderlich et al. 2019). SEDI ranges from − 1 to 1, where values above zero indicate predictions that are better than random, zero represents predictions no better than random, and below zero indicates worse than random performance. Road segments that were consistently above the high-risk threshold for all 50 training model iterations and across all three SDM approaches were classified as consensus high-risk. We report the number of ocelot and bobcat mortality occurrences that fall in consensus high-risk road segments for their respective model predictions.
Surrogate evaluation
To evaluate how well our bobcat risk predictions performed as a surrogate for ocelot risk, we assessed the overlap of road segments predicted as high-risk between ocelots and bobcats. We also used our ocelot testing data to calculate average minimum predicted area (MPA) for ocelot and bobcat risk models to compare their respective performance with ocelot testing data. We defined MPA as the minimum number of road segments categorized as high-risk to cover 90% of the occurrence testing data (Engler et al. 2004), where lower MPA values are indicative of a model that is more precise in discerning between occurrence and background locations (Fitzpatrick et al. 2013).
Results
Road mortality occurrence data
Over the 35-year data set, recorded mortalities included 46 ocelot road mortalities (41 verified, 5 likely) and 101 bobcat road mortalities (99 verified, 2 likely) across Cameron, Willacy, and Kenedy Counties. Ocelot mortality summary statistics from 1983 to 2018 can be found in Appendix B in the supplementary materials. Forty ocelot mortalities had sufficiently precise location data, whereas 80 bobcat road mortality locations occurred within the 5 km buffer of ocelot road mortalities and had acceptable location data for our presence-only SDM analyses.
Risk models
Based on our model calibration and input variable selection methods across the three SDM approaches, we identified four significant predictors of ocelot road mortality risk, including core area of scrub, distance to scrub patch intersecting roadway, road density, and edge contrast of scrub habitat with developed land cover types (Table 2). Contrast of scrub habitat edge with developed land cover types quantified fragmentation and degradation of scrub habitat. Ocelot road mortality risk increased with greater core area of scrub, while ocelot road mortality risk decreased with greater distance to scrub patch intersecting roadway, road density, and scrub habitat edge contrast with developed land cover types (Fig. 2). Two of the four predictors were also significant for bobcats, as road mortality risk decreased with increasing distance to scrub patch intersecting the roadway and increasing scrub habitat edge contrast with developed land cover types (Fig. 3). Road density and core area of scrub did not perform well in preliminary bobcat model runs and we excluded these covariates from final bobcat models (Table 2).

Relationship of ocelot road mortality risk to input variables (distance to scrub patch intersecting road, edge contrast of scrub habitat with developed land cover types, core area of scrub, and road density) for a MaxEnt, b MaxNet, and c Maxlike SDM approaches. 95% confidence intervals are shown around response curves. Input predictors were back-transformed to the original scale for ease of interpretation

Relationship of bobcat road mortality risk to input variables (distance to scrub patch intersecting road and edge contrast of scrub habitat with developed land cover types) for a MaxEnt, b MaxNet, and c Maxlike SDM approaches. 95% confidence intervals are shown around response curves. Input predictors were back-transformed to the original scale for ease of interpretation
Model coefficients were comparable in sign and relative magnitude across risk models for each species, which is indicative of comparable performance among SDM frameworks (Table 2; Merow and Silander 2014). Evaluation metrics, AUCtest and Bcont, were generally consistent across approaches indicating an acceptable fit of the models to the data for both species (Fig. 4). Average ocelot AUCtest was 0.80 for all three SDM approaches, and Bcont ranged from 0.72 to 0.82. Average bobcat AUCtest was 0.66 for all three SDM approaches, and Bcont ranged from 0.65 to 0.79.

Boxplots of evaluation metrics calculated using reserved testing data for ocelot and bobcat SDMs (MaxEnt, MaxNet, MaxLike). Upper graph shows boxplots for the area under the receiver operator characteristic curve (AUCTest) metric. Lower graph shows boxplots for the continuous Boyce index (BCont) metric
Risk maps
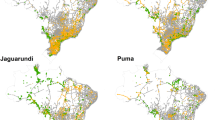
Discretization of the south Texas road network into 500 m segments yielded 2726 road segments that could be used for model predictions. For ocelots, MaxEnt, MaxNet, and MaxLike identified 322, 335, and 319 high-risk road segments respectively, which yielded 313 consensus high-risk road segments across all three models and accounted for 11% of the total road network (Fig. 5a). Average SEDI ranged from 0.68 to 0.71, and average sensitivity ranged from 0.77 to 0.82 across SDM approaches. For bobcats, MaxEnt, MaxNet, and MaxLike identified 551, 729, and 729 high-risk road segments respectively, which yielded 551 consensus high-risk road segments and accounted for 20% of the total road network (Fig. 5b). Average SEDI was 0.42, and average sensitivity ranged from 0.74 to 0.75 across SDM approaches. 25 of 40 ocelot road mortalities fell in consensus high risk road segments as defined by ocelot model predictions, and 47 of 80 bobcat road mortalities fell in consensus high risk road segments, defined as those segments that were consistently above the high-risk threshold for all 50 training model iterations and across all three SDM approaches.

Binary high-low road mortality risk classifications across the south Texas road network derived from consensus agreement of SDM modeling approaches for a ocelots and b bobcats
Surrogate evaluation
When we compared the consensus high-risk road segments between ocelots and bobcats, we found overlap in 307 high-risk road segments. This accounted for 98% of ocelot high-risk road segments. Bobcat road mortality risk predictions identified an additional 244 segments as high-risk that were not classified high-risk for ocelots. Average MPA, or the number of road segments classified as high-risk to correctly predict 90% of ocelot testing data, resulted in 833 road segments and 831 road segments for ocelots and bobcats, respectively. Comparable MPA values indicate that both species’ SDMs perform similarly in terms of predictive accuracy when tested with ocelot road mortality data.
Discussion
Building on previous applications of SDMs to road ecology research (Garrote et al. 2018; Ha and Shilling 2018; Kantola et al. 2019), we successfully applied this method to detect relationships between wild felid road mortality vulnerability and landscape and road attributes, employing a rigorous consensus modeling approach using three presence-only SDMs to account for individual modeling framework limitations and ensure the validity and consistency of results across modeling frameworks. Areas with intact, preferred scrub habitat intersecting roadways were associated with increased ocelot road mortality risk, while factors associated with habitat fragmentation and degradation such as high road density and high contrast of scrub habitat with developed land cover types were associated with lower road mortality risk. Greater road mortality risk in areas with greater amounts of preferred habitat is expected because these areas are more likely to be occupied, a trend that has been found in other studies exploring landscape predictors of road mortality risk for ungulates, small mammals, carnivores, and birds (Grilo et al. 2012; Barthelmess 2014; Ha and Shilling 2018). High-risk road segments were predominantly in areas with protected natural habitat and private undeveloped rangeland, with minimal development and urban infrastructure, underscoring the importance of intact scrub habitat for ocelots and supporting their classification as a fragmentation-intolerant, Tamaulipan thornscrub specialist in this region (Horne et al. 2009; Janečka et al. 2016). The importance of high-quality intact habitat for ocelot abundance in anthropogenically impacted landscapes has also been highlighted in other areas of the ocelot range (Paolino et al. 2018). Our results confirm that ocelots in south Texas are fragmentation intolerant and demonstrate the implications of this relationship for ocelot road mortality risk.
Comparison between ocelot and bobcat risk models indicates that, with specific data filters in place and when landscape-level data are used, bobcats can be useful surrogates to inform ocelot road mortality risk. This is likely to be of particular importance when ocelot data are limited. Given the modest amount of ocelot data available after 35 years of monitoring (n = 40), timely decision-making to reduce road mortality on shorter temporal scales or for specific mitigation locations will likely need to rely more heavily on bobcat data, keeping in mind the differences in habitat selection for co-occurring ocelots and bobcats (sensu Horne et al 2009).
As the first quantitative comparison of ocelot and bobcat road interactions and road mortality risk in south Texas, our research provides a baseline that can inform future studies aiming to use bobcat data to inform ocelot road mortality risk and mitigation. Bobcat risk models incorporated two of the four road mortality risk predictors for ocelots, and the relationships of these predictors to road mortality risk were consistent between ocelots and bobcats; decreasing distance to scrub habitat intersecting the road and decreasing scrub habitat edge contrast with developed land cover increased road mortality risk for both species. These relationships indicate that bobcats occurring in the same areas as ocelots also exhibit some level of landscape fragmentation intolerance, though likely not to the same degree of ocelot intolerance. For example, both Janečka et al. (2016) and Taylor (2017) found evidence of restricted genetic dispersal for bobcats in extremely fragmented habitats in south Texas, though the genetic impacts of fragmentation were more severe for ocelots. This supports previous descriptions of the bobcat as having intermediate sensitivity to habitat fragmentation (Crooks 2002; Abouelezz et al. 2018). Future applications with bobcats as ocelot surrogates would benefit from additional habitat data, to focus more on intact thornscrub habitats relevant to ocelot populations.
Because the areas of highest ocelot road mortality risk overlap directly with intact thornscrub habitat, our results can be used to inform targeted mitigation efforts including wildlife underpasses, speed limits, signage, and restoration (Fig. 5). Through a productive collaboration with USFWS and the Texas Department of Transportation, a suite of wildlife underpasses were recently constructed on roadways that have been sources of ocelot road mortality in Cameron County. The underpass locations were originally chosen based on expert opinion of USFWS ocelot biologists employing a visual assessment of previous ocelot mortalities and ocelot telemetry data. Our analysis shows that a majority of these underpasses are located within high-risk road segments as defined by our modeling approach, suggesting that these crossings are well-placed to mitigate ocelot-vehicle collision risk. Fencing and speed limits, specifically night speed limits, may also be needed to effectively reduce road mortality risk (Meisingset et al. 2014; Polak et al. 2019).
In addition to mitigation of direct mortality from vehicle collisions, restoration of Tamaulipan thornscrub has also been identified as an important element of ocelot recovery efforts (Haines et al. 2005b). Recent Tamaulipan thornscrub restoration efforts have been focused in areas where ocelot populations are extant to encourage expansion of breeding female ranges through dispersal (USFWS 2016; Alexander et al. 2016). These long-term projects aim to create additional habitat for breeding females and to restore and fortify corridors of Tamaulipan thornscrub between currently isolated populations within south Texas (Lehnen et al. in review). The higher mortality risk associated with scrub habitat intersecting roadways suggests that wildlife crossings should be installed along corridors of preferred habitat, both existing and restored, that intersect roadways to ensure that these areas do not become an additional source of road mortality.
Ocelot movement and resource use data can also provide an important complement to road mortality data to identify drivers of road mortality risk, because mortality data alone may not be able to fully describe the drivers of road mortality risk (Teixeira et al. 2017). Location data and connectivity modeling that quantifies movement across a road network (Loraamm and Downs 2016), as well as connectivity corridors with pinch-points that intersect roads (Chetkiewicz and Boyce 2009; McClure et al. 2017), can be leveraged to inform targeted road mitigation and restoration efforts. Recent work by Lehnen et al. in review applied least cost path and circuit theory methods to evaluate landscape connectivity and identify key habitat linkages between isolated ocelot populations in south Texas. Pairing these types of movement analyses with road mortality risk mapping can support the identification of road segments that could benefit from mitigation action. Additionally, by incorporating bobcat data into these analyses, continued evaluation of how bobcats can serve as an effective surrogate for ocelot research can be extended from road mortality risk to movement and space use analyses.
Our analyses present a robust approach to determine the predictors of road mortality risk for the south Texas ocelot, and there is clear evidence that ocelot road mortality risk is highest in areas of intact Tamaulipan thornscrub habitat. Given the limited extent of this intact habitat, focused road mitigation efforts that target these areas may successfully mitigate the risk from ocelot-vehicle collisions. Our findings support the use of bobcats as a surrogate species for ocelots in the context of road mortality risk if bobcat data are filtered to habitats and spatial extents relevant to ocelots in this region. For ocelots, and other depleted populations, our research also provides a foundation for future efforts to integrate movement, dispersal, and landscape connectivity data to inform our understanding of the risks to ocelots and other medium-sized carnivores in the context of natural and built landscape features.
References
Abouelezz HG, Donovan TM, Mickey RM, Murdoch JD, Freeman M, Royar K (2018) Landscape composition mediates movement and habitat selection in bobcats (Lynx rufus): implications for conservation planning. Landsc Ecol 33:1301–1318. https://doi.org/10.1007/s10980-018-0654-8
Alexander HD, Moczygemba J, Dick K (2016) Growth and survival of thornscrub forest seedlings in response to restoration strategies aimed at alleviating abiotic and biotic stressors. J Arid Environ 124:180–188. https://doi.org/10.1016/j.jaridenv.2015.06.014
Balkenhol N, Waits LP (2009) Molecular road ecology: exploring the potential of genetics for investigating transportation impacts on wildlife. Mol Ecol 18:4151–4164. https://doi.org/10.1111/j.1365-294X.2009.04322.x
Barrientos R, Bolonio L (2009) The presence of rabbits adjacent to roads increases polecat road mortality. Biodivers Conserv 18:405–418. https://doi.org/10.1007/s10531-008-9499-9
Barthelmess EL (2014) Spatial distribution of road-kills and factors influencing road mortality for mammals in Northern New York State. Biodivers Conserv 23:2491–2514. https://doi.org/10.1007/s10531-014-0734-2
Bennett VJ, Sparks DW, Zollner PA (2013) Modeling the indirect effects of road networks on the foraging activities of bats. Landsc Ecol 28:979–991. https://doi.org/10.1007/s10980-013-9874-0
Booth-Binczik SD, Bradley RD, Thompson CW, Bender LC, Huntley JW, Harvey JA, Laack LL, Mays JL (2013) Food habits of ocelots and potential for competition with bobcats in southern Texas. Southwest Nat 58:403–410
Breiner FT, Guisan A, Bergamini A, Nobis MP (2015) Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol Evol 6:1210–1218. https://doi.org/10.1111/2041-210X.12403
Ceia-Hasse A, Borda-de-Água L, Grilo C, Pereira HM (2017) Global exposure of carnivores to roads. Glob Ecol Biogeogr 26:592–600. https://doi.org/10.1111/geb.12564
Ceia-Hasse A, Navarro LM, Borda-de-Água L, Pereira HM (2018) Population persistence in landscapes fragmented by roads: disentangling isolation, mortality, and the effect of dispersal. Ecol Modell 375:45–53. https://doi.org/10.1016/j.ecolmodel.2018.01.021
Červinka J, Riegert J, Grill S, Šálek M (2015) Large-scale evaluation of carnivore road mortality: the effect of landscape and local scale characteristics. Mammal Res 60:233–243. https://doi.org/10.1007/s13364-015-0226-0
Chetkiewicz C-LB, Boyce MS (2009) Use of resource selection functions to identify conservation corridors. J Appl Ecol 46:1036–1047
Crooks KR (2002) Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv Biol 16:488–502. https://doi.org/10.1046/j.1523-1739.2002.00386.x
Crooks KR, Burdett CL, Theobald DM, King SRB, Di Marco M, Rondinini C, Boitani L (2017) Quantification of habitat fragmentation reveals extinction risk in terrestrial mammals. Proc Natl Acad Sci 114:7635–7640. https://doi.org/10.1073/pnas.1705769114
Crooks KR, Burdett CL, Theobald DM, Rondinini C, Boitani L (2011) Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philos Trans R Soc B Biol Sci 366:2642–2651. https://doi.org/10.1098/rstb.2011.0120
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Ellis EC (2011) Anthropogenic transformation of the terrestrial biosphere. Philos Trans R Soc A 369:1010–1035. https://doi.org/10.1098/rsta.2010.0331
Engler R, Guisan A, Rechsteiner L (2004) An improved approach for predicting the distribution of rare and endangered species from occurrence and pseudo-absence data. J Appl Ecol 41:263–274. https://doi.org/10.1111/j.0021-8901.2004.00881.x
ESRI (2018) ArcGIS version 10.6. Environmental Systems Research Institute, Redlands, California, USA
Ferreras P, Gaona P, Palomares F, Delibes M (2001) Restore habitat or reduce mortality? Implications from a population viability analysis of the Iberian lynx. Anim Conserv 4:265–274. https://doi.org/10.1017/S1367943001001317
Ferro CAT, Stephenson DB (2011) Extremal dependence indices: improved verification measures for deterministic forecasts of rare binary events. Weather Forecast 26:699–713. https://doi.org/10.1175/WAF-D-10-05030.1
Fitzpatrick MC, Gotelli NJ, Ellison AM (2013) MaxEnt versus MaxLike: empirical comparisons with ant species distributions. Ecosphere 4(5):55. https://doi.org/10.1890/ES13-00066.1
Forman RTT (2003) Road ecology: science and solutions. Island Press, Washington
Garrote G, Fernández-López J, López G, Ruiz G, Simón MA (2018) Prediction of Iberian lynx road–mortality in southern Spain: a new approach using the MaxEnt algorithm. Anim Biodivers Conserv 2:217–226
Glista DJ, DeVault TL, DeWoody JA (2009) A review of mitigation measures for reducing wildlife mortality on roadways. Landsc Urban Plan 91:1–7. https://doi.org/10.1016/j.landurbplan.2008.11.001
Grilo C, Ascensão F, Santos-Reis M, Bissonette JA (2011) Do well-connected landscapes promote road-related mortality? Eur J Wildl Res 57:707–716. https://doi.org/10.1007/s10344-010-0478-6
Grilo C, Bissonette JA, Santos-Reis M (2009) Spatial-temporal patterns in Mediterranean carnivore road casualties: Consequences for mitigation. Biol Conserv 142:301–313. https://doi.org/10.1016/j.biocon.2008.10.026
Grilo C, Sousa J, Ascensão F, Matos H, Leitão I, Pinheiro P, Costa M, Bernardo J, Reto D, Lourenço R, Santos-Reis M, Revilla E (2012) Individual spatial responses towards roads: implications for mortality risk. PLoS One 7:1–11. https://doi.org/10.1371/journal.pone.0043811
Gunson K, Teixeira FZ (2015) Road-wildlife mitigation planning can be improved by identifying the patterns and processes associated with wildlife-vehicle collisions. Handb Road Ecol. https://doi.org/10.1002/9781118568170.ch13
Ha H, Shilling F (2018) Modelling potential wildlife-vehicle collisions (WVC) locations using environmental factors and human population density: a case-study from 3 state highways in Central California. Ecol Inform 43:212–221. https://doi.org/10.1016/j.ecoinf.2017.10.005
Haines AM, Grassman LI Jr, Tewes ME, Janečka JE (2006) First ocelot (Leopardus pardalis) monitored with GPS telemetry. Eur J Wildl Res 52:216–218. https://doi.org/10.1007/s10344-006-0043-5
Haines AM, Tewes ME, Laack LL (2005a) Survival and sources of mortality in ocelots. J Wildl Manage 69:255–263
Haines AM, Tewes ME, Laack LL, Grant WE, Young J (2005b) Evaluating recovery strategies for an ocelot (Leopardus pardalis) population in the United States. Biol Conserv 126:512–522. https://doi.org/10.1016/j.biocon.2005.06.032
Harveson PM, Tewes ME, Anderson GL, Laack LL (2004) Habitat use by ocelots in south Texas: implications for restoration. Wildl Soc Bull 32:948–954
Hirzel AH, Le Lay G, Helfer V, Randin C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Modell 199:142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017
Homer CG, Dewitz JA, Yang L, Jin S, Danielson P, Xian G, Coulston J, Herold ND, Wickham JD, Megown K (2015) Completion of the 2011 National Land Cover Database (NLCD) for the conterminous United States-Representing a decade of land cover change information. Photogramm Eng Remote Sens 81:345–354 https://doi.org/10.14358/PERS.81.5.345
Horne JS, Haines AM, Tewes ME, Laack LL (2009) Habitat partitioning by sympatric ocelots and bobcats: implications for recovery of ocelots in southern Texas. Southwest Nat 54:119–126. https://doi.org/10.1894/PS-49.1
Jackson VL, Laack LL, Zimmerman EG (2005) Landscape metrics associated with habitat use by ocelots in south Texas. J Wildl Manage 69:733–738
Jahrsdoerfer SE, Leslie DMJ (1988) Tamaulipan brushland of the Lower Rio Grande Valley of south Texas: description, human impacts, and management options. Biol Rep IV USFWS 88:63
Janečka JE, Tewes ME, Davis IA, Haines AM, Caso A, Blankenship TL, Honeycutt RL (2016) Genetic differences in the response to landscape fragmentation by a habitat generalist, the bobcat, and a habitat specialist, the ocelot. Conserv Genet 17(5):1093–1108. https://doi.org/10.1007/s10592-016-0846-1
Janečka JE, Tewes ME, Laack LL, Caso A, Grassman LI, Haines AM, Shindle DB, Davis BW, Murphy WJ, Honeycutt RL (2011) Reduced genetic diversity and isolation of remnant ocelot populations occupying a severely fragmented landscape in southern Texas. Anim Conserv 14:608–619. https://doi.org/10.1111/j.1469-1795.2011.00475.x
Janečka JE, Tewes ME, Laack L, Caso A, Grassman LI, Honeycutt RL (2014) Loss of genetic diversity among ocelots in the United States during the 20th century linked to human induced population reductions. PLoS One 9:1–10. https://doi.org/10.1371/journal.pone.0089384
Jin S, Homer C, Yang L, Danielson P, Dewitz J, Li C, Zhu Z, Xian G, Howard D (2019) Overall methodology design for the United States national land cover database (NLCD) 2016 products. Remote Sens. https://doi.org/10.3390/rs11242971
Kantola T, Tracy JL, Baum KA, Quinn MA, Coulson RN (2019) Spatial risk assessment of eastern monarch butterfly road mortality during autumn migration within the southern corridor. Biol Conserv 231:150–160. https://doi.org/10.1016/j.biocon.2019.01.008
Keinath DA, Doak DF, Hodges KE, Prugh LR, Fagan W, Sekercioglu CH, Buchart SHM, Kauffman M (2017) A global analysis of traits predicting species sensitivity to habitat fragmentation. Glob Ecol Biogeogr 26:115–127. https://doi.org/10.1111/geb.12509
Kramer-Schadt S, Niedballa J, Pilgrim JD, Schröder B, Lindenborn J, Reinfelder V, Stillfried M, Heckmann I, Scharf AK, Augeri DM, Cheyne SM, Hearn AJ, Ross J, Macdonald DW, Mathai J, Eaton J, Marshall AJ, Semiadi G, Rustam R, Bernard H, Alfred R, Samejima H, Duckworth JW, Breitenmoser-Wuersten C, Belant JL, Hofer H, Wilting A (2013) The importance of correcting for sampling bias in MaxEnt species distribution models. Divers Distrib 19:1366–1379. https://doi.org/10.1111/ddi.12096
Laack L (1991) Ecology of the ocelot (Felis pardalis) in south Texas. Texas A & I University, Kingsville
Leonard JP (2016) Home range characteristics, activity patterns, and resource selection of sympatric ocelots (Leopardus pardalis) and bobcats (Lynx rufus) and major histocompatibility complex variation in ocelots. Texas A&M, Kingsville
Lima Santos RA, Ascensão F, Ribeiro ML, Bager A, Santos-Reis M, Aguiar LMS (2017) Assessing the consistency of hotspot and hot-moment patterns of wildlife road mortality over time. Perspect Ecol Conserv 15:56–60. https://doi.org/10.1016/j.pecon.2017.03.003
Lin YP, Anthony J, Lin WC, Lien WY, Petway JR, Lin TE (2019) Spatiotemporal identification of roadkill probability and systematic conservation planning. Landsc Ecol 34:717–735. https://doi.org/10.1007/s10980-019-00807-w
Litvaitis JA, Reed GC, Carroll RP, Litvaitis MK, Tash J, Mahard T, Broman DJA, Callahan C, Ellingwood M (2015) Bobcats (Lynx rufus) as a model organism to investigate the effects of roads on wide-ranging carnivores. Environ Manage 55:1366–1376. https://doi.org/10.1007/s00267-015-0468-2
Liu C, Berry PM, Dawson TP, Pearson RG (2005) Selecting thresholds of occurrence in the prediction of species distributions. Ecography (Cop) 28:385–393. https://doi.org/10.1111/j.0906-7590.2005.03957.x
Liu C, Newell G, White M (2016) On the selection of thresholds for predicting species occurrence with presence-only data. Ecol Evol 6:337–348. https://doi.org/10.1002/ece3.1878
Liu C, White M, Newell G (2013) Selecting thresholds for the prediction of species occurrence with presence-only data. J Biogeogr 40:778–789. https://doi.org/10.1111/jbi.12058
Loraamm RW, Downs JA (2016) A wildlife movement approach to optimally locate wildlife crossing structures. Int J Geogr Inf Sci 30:74–88. https://doi.org/10.1080/13658816.2015.1083995
Marmion M, Parviainen M, Luoto M, Heikkinen RK, Thuiller W (2009) Evaluation of consensus methods in predictive species distribution modelling. Divers Distrib 15:59–69. https://doi.org/10.1111/j.1472-4642.2008.00491.x
McClure ML, Dickson BG, Nicholson KL (2017) Modeling connectivity to identify current and future anthropogenic barriers to movement of large carnivores: a case study in the American Southwest. Ecol Evol 7:3762–3772. https://doi.org/10.1002/ece3.2939
McGarigal K, Cushman SA, Ene E (2012) FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps
Meisingset EL, Loe LE, Brekkum Ø, Mysterud A (2014) Targeting mitigation efforts: the role of speed limit and road edge clearance for deer-vehicle collisions. J Wildl Manage 78:679–688. https://doi.org/10.1002/jwmg.712
Merow C, Silander JAJ (2014) A comparison of Maxlike and Maxent for modelling species distributions. Methods Ecol Evol 5:215–225. https://doi.org/10.1111/2041-210X.12152
Merow C, Smith MJ, Silander JA (2013) A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography (Cop) 36:1058–1069. https://doi.org/10.1111/j.1600-0587.2013.07872.x
Newbold T, Hudson LN, Arnell AP, Contu S, De Palma A, Ferrier S, Hill SLL, Hoskins AJ, Lysenko I, Phillips HRP, Burton VJ, Chng CWT, Emerson S, Gao D, Hale GP, Hutton J, Jung M, Sanchez-Ortiz K, Simmons BI, Whitmee S, Zhang H, Scharlemann JPW, Purvis A (2016) Has land use pushed terrestrial biodiversity beyond the planetary boundary? A global assessment. Science 353:291–288. https://doi.org/10.1126/science.aaf2201
Paolino RM, Royle JA, Versiani NF, Rodrigues TF, Pasqualotto N, Krepschi VG, Chiarello AG (2018) Importance of riparian forest corridors for the ocelot in agricultural landscapes. J Mammal 99:874–884. https://doi.org/10.1093/jmammal/gyy075
Phillips SJ, Anderson RP, Dudík M, Schapire RE, Blair ME (2017) Opening the black box: an open-source release of Maxent. Ecography (Cop) 40:887–893. https://doi.org/10.1111/ecog.03049
Phillips SJ, Anderson RP, Shapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Modell 190:231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Phillips SJ, Dudík M, Elith J, Graham CH, Lehmann A, Leathwick J, Ferrier S (2009) Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol Appl 19:181–197. https://doi.org/10.1890/07-2153.1
Polak T, Nicholson E, Grilo C, Bennett JR, Possingham HP (2019) Optimal planning to mitigate the impacts of roads on multiple species. J Appl Ecol 56:201–213. https://doi.org/10.1111/1365-2664.13258
Proctor MF, McLellan BN, Strobeck C, Barclay RM (2005) Genetic analysis reveals demographic fragmentation of grizzly bears yielding vulnerably small populations. Proc R Soc B 272:2409–2416. https://doi.org/10.1098/rspb.2005.3246
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.Rproject.org/
Riley SPD, Pollinger JP, Sauvajot RM, York EC, Bromley C, Fuller TK, Wayne RK (2006) A southern California freeway is a physical and social barrier to gene flow in carnivores. Mol Ecol 15:1733–1741. https://doi.org/10.1111/j.1365-294X.2006.02907.x
Roger E, Laffan SW, Ramp D (2011) Road impacts a tipping point for wildlife populations in threatened landscapes. Popul Ecol 53:215–227. https://doi.org/10.1007/s10144-010-0209-6
Royle JA, Chandler RB, Yackulic C, Nichols JD (2012) Likelihood analysis of species occurrence probability from presence-only data for modelling species distributions. Methods Ecol Evol 3:545–554. https://doi.org/10.1111/j.2041-210X.2011.00182.x
Seiler A (2004) Trends and spatial patterns in ungulate-vehicle collisions in Sweden. Wildl Biol 10:301–313. https://doi.org/10.2981/wlb.2004.036
Taylor DR (2017) Genetic analysis of bobcats (Lynx rufus) and ocelots (Leopardus pardalis) in a fragmented landscape using noninvasive sampling. Texas A&M, Kingsville
Teixeira FZ, Coelho IP, Esperandio IB, Oliveira NR, Porto F, Dornelles SS, Delazeri NR, Tavares M, Martins MB, Kindel A (2013) Are road-kill hotspots coincident among different vertebrate groups? Oecol Austral 17:36–47
Teixeira FZ, Kindel A, Hartz SM, Mitchell S, Fahrig L (2017) When road-kill hotspots do not indicate the best sites for road-kill mitigation. J Appl Ecol 54:1544–1551. https://doi.org/10.1111/1365-2664.12870
Tremblay TA, White WA, Raney JA, Trembiay TA, White WA, Raney JAYA (2005) Native woodland loss during the mid 1900s in Cameron County, Texas. Southwest Nat 50:479–482
USFWS (2016) Recovery Plan for the Ocelot (Leopardus pardalis). U.S. Fish and Wildlife Service, Southwest Region, Albuquerque, New Mexico
van Der Ree R, Jaeger JAG, van Der Grift EA, Clevenger AP (2011) Effects of roads and traffic on wildlife populations and landscape function: road ecology is moving toward larger scales. Ecol Soc 16(1):48. https://doi.org/10.2307/26268822
van der Ree R, Smith DJ, Grilo C (2015) Handbook of road ecology. Wiley, Hoboken
van der Ree R, van der Grift E, Gulle N, Holland K, Mata C, Suarez F (2007) Overcoming the barrier effect of roads- how effective are mitigation strategies? An Int Rev use Eff underpasses overpasses Des to increase permeability roads wildlife pp 423–431
Van Langevelde F, Jaarsma CF (2005) Using traffic flow theory to model traffic mortality in mammals. Landsc Ecol 19:895–907. https://doi.org/10.1007/s10980-005-0464-7
Visintin C, van der Ree R, McCarthy MA (2016) A simple framework for a complex problem? Predicting wildlife–vehicle collisions. Ecol Evol 6:6409–6421. https://doi.org/10.1002/ece3.2306
Wunderlich RF, Lin YP, Anthony J, Petway JR (2019) Two alternative evaluation metrics to replace the true skill statistic in the assessment of species distribution models. Nat Conserv 35:97–116. https://doi.org/10.3897/natureconservation.35.33918
Yang L, Jin S, Danielson P, Homer C, Gass L, Bender SM, Case A, Costello C, Dewitz J, Fry J, Funk M, Granneman B, Liknes GC, Rigge M, Xian G (2018) A new generation of the United States National Land Cover Database (NLCD): Requirements, research priorities, design, and implementation strategies. ISPRS J Photogramm Remote Sens 146:108–123. https://doi.org/10.1016/j.isprsjprs.2018.09.006
Hijmans RJ, Phillips S, Leathwick J, Elith J (2017). dismo: Species Distribution Modeling. R package version 1.1–4. https://CRAN.R-project.org/package=dismo
US Geological Survey (2016a) National Hydrography Dataset (ver. USGS National Hydrography Dataset Best Resolution) https://www.usgs.gov/core-science-systems/ngp/national-hydrography/access-national-hydrography-products
US Geological Survey (2016b) USGS National Elevation Dataset (NED) 1 arc-second Downloadable Data Collection from The National Map 3D Elevation Program (3DEP) - National Geospatial Data Asset (NGDA) National Elevation Dataset (NED): U.S. Geological Survey.
Acknowledgements
Thank you to researchers in addition to U.S. Fish and Wildlife biologists who have collected road mortality data since 1983. Thanks to two anonymous reviewers for providing constructive comments that improved this manuscript. We also appreciate reviews by M. Sternberg (USFWS), M. Jennings, A. Nara and L. McClenaghan (SDSU). This material is based upon work supported by the National Science Foundation Graduate Research Fellowship under Grant No. (1321850).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Schmidt, G.M., Lewison, R.L. & Swarts, H.M. Identifying landscape predictors of ocelot road mortality. Landscape Ecol 35, 1651–1666 (2020). https://doi.org/10.1007/s10980-020-01042-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-020-01042-4