
Abstract
Context
North American grassland songbird populations have declined significantly due to habitat loss and fragmentation. Understanding the influence of the surrounding landscape on prairie fragment occupancy is vital for predicting the fate of grassland birds in these heavily altered landscapes.
Objectives
We examined the relative importance of local and landscape variables on grassland bird occupancy of prairie fragments using a focal-patch study. We also investigated the spatial scale at which landscape variables were most influential.
Methods
We surveyed birds on 29 unplowed prairie fragments in western Minnesota and eastern North and South Dakota. We quantified local habitat on the fragment using vegetation surveys and aerial photographs and the landscape surrounding the fragment out to 4 km using aerial photographs. We analyzed occupancy using multi-model approaches applied to multiple logistic regression.
Results
Of 38 species encountered, nine were neither too rare nor too abundant to be analyzed. Predictors of patch occupancy were unique for each bird species, yet general patterns emerged. For eight species, landscape variables were more important than local variables. Mostly, those landscape variables measured configuration (e.g., edge density) and not composition (e.g., percent cover of a particular matrix element). Landscape effects were mostly from variables measured at the greatest extents from the prairie fragment.
Conclusions
Using a focal-patch study design we demonstrated the importance of the surrounding landscape, often out to 4 km from the fragment edge, on prairie occupancy by grassland birds. Effective management of grassland songbirds will require attention to the landscape context of prairie fragments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Globally, grasslands are among the most threatened biomes (Hoekstra et al. 2005). Grasslands across the globe face habitat loss and fragmentation due to habitat conversion (Askins et al. 2007) and habitat degradation due to increasing grazing pressure and fire suppression (Fuhlendorf et al. 2008). In grasslands around the world, habitat loss and degradation has negatively impacted biota including plants (Piqueray et al. 2011; Deák et al. 2016), insects (Moranz et al. 2014; Otto et al. 2016; Poniatowski et al. 2016), herptiles (Gebauer et al. 2013; Rotem et al. 2016), and mammals (Duggan et al. 2011). Grassland birds have been particularly well studied with effects of grassland conversion and degradation documented broadly, such as in Spain (Reino et al. 2012), Brazil (Fontana et al. 2016), and Australia (Baker-Gabb et al. 2016). Overall, grassland bird populations are declining in Europe and North America (Reif 2013).
North America’s grasslands have experienced substantial impacts from agricultural expansion and increasing urbanization. In the Great Plains, almost 70% of historic grassland cover has been lost (Samson et al. 2004), with higher losses in the northern tallgrass prairie, where <1% of the original tallgrass prairie remains (Samson and Knopf 1994). Additionally, the current economic climate continues to drive substantial grassland conversion to agriculture (Wright and Wimberly 2013). Such substantial and on-going habitat loss and fragmentation impacts prairie species, including birds. North American grassland songbirds are experiencing the steepest population declines compared to any other group of birds on the continent (North American Bird Conservation Initiative Canada 2012). From 1968 to 2008, 37% of grassland obligate species declined (Sauer and Link 2011). Even common species like Savannah Sparrows (Passerculus sandwichensis) and Clay-colored Sparrows (Spizella pallida) have declined in at least a part of their range (Igl and Johnson 1997).
In the face of these declines, much research has been done on grassland songbird populations at the habitat patch level including effects of habitat loss and fragmentation (Herkert 1994; Bender et al. 1998; Helzer and Jelinski 1999; Johnson and Igl 2001; Davis and Brittingham 2004; Fletcher 2005) and habitat quality and vegetation structure (O’Leary and Nyberg 2000; Davis 2005; With et al. 2008). Multiple bird species are area- or edge-sensitive (DeLisle and Savidge 1996; Winter et al. 2000; Jensen and Finck 2004; Koper and Manseau 2009) and respond to specific vegetation or structural features (Whitmore 1981; Davis et al. 1999; Winter et al. 2005; Jacobs et al. 2012). These details, while important, tend to ignore that prairie fragments do not exist in isolation, but are embedded in landscapes which vary in terms of the types, amounts, and configuration of matrix (non-habitat) elements. Matrix can provide secondary habitat (Johnson 2000), alter predation or parasitism rates (Borgmann and Rodewald 2004; Patten et al. 2006), and influence dispersal (Haas 1995), all of which can potentially impact grassland birds on prairie remnants. A recent multi-taxa review found that the type of matrix surrounding a patch influenced species richness or abundance in 95% of the studies reviewed (Prevedello and Vieria 2010). Despite this, few studies have analyzed the influence of the surrounding landscape on populations on patches of remnant habitat (Rodewald 2003).
Landscape-level effects may not be identical across all songbirds. Each species has its own specialized suite of resources needed for foraging and reproduction (Ehrlich et al. 1988) that may cause different species to respond differently to landscape elements. Similarly, dispersal, movement, and avoidance behaviors, which can vary between species, could also influence species responses to the surrounding landscape (Eycott et al. 2012), including the spatial scale at which species-landscape interactions manifest. Connecting landscape patterns to life history traits may allow for extrapolation of trends to species with similar resource requirements (such as habitat preferences or functional groups). Identifying such connections between species behaviors and life history traits would permit the development of management techniques that are suitable for more than a single target species making it easier to maximize conservation resources. These patterns could also be used to identify prairies on which particular groups of species are at risk or in need of additional of management attention. A previous study found that it is possible to predict forest bird community responses to landscape changes using species’ life history traits (Hansen and Urban 1992), and it makes sense to try incorporating the same kinds of information for grassland songbirds.
Studying landscape-level effects is best done using a focal-patch approach (Brennan et al. 2002) where each habitat patch and surrounding landscape represents an experimental unit in the analysis and replication occurs at the landscape level rather than the habitat patch level. This focal-patch approach allows effects due to the surrounding landscape to be separated from those due to local characteristics of the habitat patch (Thornton et al. 2011) and minimizes the likelihood of spatial autocorrelation influencing the analysis (Brennan et al. 2002). Despite the obvious advantages of the focal-patch approach, many of the previous landscape studies of grassland songbirds were based on sampling units not focal patches (e.g., Ribic and Sample 2001; Bakker et al. 2002; Kalinowski and Johnson 2010) making it difficult to distinguish habitat and landscape effects in their results.
Most grassland songbirds migrate at least short distances every year (Igl and Johnson 1997) requiring them to choose a prairie patch to establish their breeding territory upon their return. This process is hierarchical, as birds are influenced by different factors at progressively smaller scales as they narrow their range of movement from large (migratory movements) to small (establishing nesting or feeding territories; Hutto 1985). As such, the final occupancy patterns may be influenced by aspects of the landscape at broader scales. To capture this effect, we used a larger spatial extent (4 km) than most of the previous landscape studies of grassland songbirds, most of which looked no farther than 2 km into the landscape (e.g., Bergin et al. 2000; Bajema and Lima 2001; Ribic and Sample 2001; Grant et al. 2004; Jacobs et al. 2012).
We report on a study investigating grassland songbird occurrence on remnant prairies embedded in heterogeneous landscapes. We used a focal-patch approach, with the focal-patch defined as the extent of the contiguous native prairie for a given remnant prairie patch, allowing us to distinguish between landscape and local effects on songbird occurrence. We quantified the landscape out to 4 km around the focal patch allowing us to incorporate some of the largest scale landscape data investigated thus far for grassland birds. Finally, we compare the bird species responses to landscape structure across habitat guilds.
Methods
Site selection
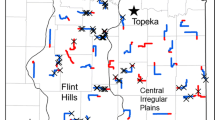
We selected 29 native, unplowed prairie fragments located in western Minnesota and eastern North and South Dakota, owned and/or managed by The Nature Conservancy, Minnesota Department of Natural Resources, U.S. Fish & Wildlife Service, or the University of North Dakota (Median patch size 67.5 ha with a range of 7.1–1180.9 ha; Fig. 1). Prairie fragments are relatively rare in this area with most being very isolated (15 of the 29 fragments did not have any other native, unplowed prairie within 4 km). Fragments were chosen to maximize the range of sampled fragment sizes and the variation in the surrounding landscape (based on cover of woody vegetation, grass, agriculture, and open water), to exclude patches scheduled for burning or grazing management during the 2-year study period, and to ensure a minimum separation of 8 km to reduce issues with spatial autocorrelation.

Field sites in North Dakota, South Dakota, and Minnesota, n = 29. Agency responsible for managing the site is indicated by symbol on the map
Bird counts
We conducted bird counts from mid-May through mid-July, on all 29 sites in 2010 and 2011. Counts occurred when birds were most active and vocal, from dawn until approximately 10:30 to 11:00 am CDT, and on days with weather conditions conducive to hearing and seeing birds (wind speeds less than 32 km/h, minimal precipitation; Bibby et al. 1992). We sampled each site twice during each field season, except when weather conditions and flooding limited access. Seven sites were surveyed twice in 2010, and 26 sites were surveyed twice in 2011.
Each count used a linear transect that allowed sampling of significant portions of each prairie patch while minimizing sampling time (Anderson and Ohmart 1981; Gibbons et al. 1996). Transect length was dictated by prairie fragment size: 400 m in the smallest fragments (7–40 ha), 800 m in the medium fragments (41–161 ha), and 1200 m in the large fragments (>161 ha). These distances were based on the length of transects that would fit within the prairie fragment (on the small and medium fragments) and time available for surveying (on the large fragments). Transects were placed at least 100 m from the edge of the prairie, to avoid edge effects that might influence the bird community (Fletcher 2005). In two cases, prairie fragments were shaped so that a standard-length transect would not fit and still be at least 100 m from the prairie’s edges. For these sites, we used shortened transects (700 and 750 m) that extended as far as the shape of the prairie allowed. Transects were a single straight line, unless the size of the prairie or the location of wetlands prevented it. In those cases we broke the transect into multiple smaller transects (Gates 1981) placed at least 300 m apart to avoid double counting birds (Davis and Brittingham 2004; Koper and Schmiegelow 2006).
Transects were walked at a steady pace and all birds seen or heard within 50 m of the transect were recorded. We used the same observer at all sites to avoid observer bias. Birds flying over the transect were only recorded if they actually landed on the prairie fragment or were observed foraging aerially above it. For each bird sighted we recorded the species and distance along the transect (determined by a hand-held GPS accurate to 3 m, Garmin eTrex H Handheld Navigator).
Local habitat characteristics
We used a Robel pole to quantify vegetation height and structure (Robel et al. 1970) every 100 m along the transect, with the pole placed 1-m off the transect to avoid the disturbance to the vegetation from walking the transect. We also visually estimated the percent cover of grasses, forbs, trees, shrubs, and bare ground within 5 m along each transect. The estimates were made along 100 m segments of the transect, then averaged over the length of the transect. These measurements were performed once during the study (2010) on the same day as the bird surveys, because the relative amount of each cover type was unlikely to change drastically between the two survey years. In addition, we interpreted and digitized (using Arc GIS 9.3 and 10.0: ESRI 2010; 2011) land cover from digital aerial photographs of each prairie fragment to determine the percent cover of grass, woody vegetation, vegetated wetlands, and open water on each prairie fragment. These images were obtained from the National Agriculture Imagery Program (NAIP), via the Minnesota Department of Natural Resources Data Deli (http://deli.dnr.state.mn.us), the North Dakota GIS Hub (http://www.nd.gov/gis), and the South Dakota Department of Environment and Natural Resources (http://www.sdgs.usd.edu). The most recent images available at the time were from 2009 (Minnesota and North Dakota) and 2008 (South Dakota).
Landscape characteristics
We collected landscape-level data by interpreting and digitizing land cover from the digital aerial photographs described in the local habitat characteristics section. We verified the aerial photographs by walking the outer perimeter of each prairie fragment, and driving through each landscape and observing the land cover. For each prairie remnant, we used GPS coordinates to locate the site on the aerial photograph and digitized the remnant boundaries based on the extent of native unplowed prairie using site maps provided by the organization managing the site. Multiple sites were surrounded by grasslands of other types [such as Conservation Reserve Program (CRP) plantings or restored prairie], which were excluded from the remnant since bird species might respond differently to these types of grasslands due to differences in vegetation structure (Ahlering and Merkord 2016).
Once the remnant patch was defined, we created a 4-km buffer around the site, to delineate landscape extent. This distance provided greater spatial extent than previously seen in most avian landscape studies (e.g., Bakker et al. 2002; Renfrew and Ribic 2008; Ribic et al. 2009a) while still being manageable for digitizing landscape features. The digitized landscapes varied from 5418 to 10,448 ha (median = 6435.6 ha, IQR: 6160.3–7578.9 ha) and were separated from their closest neighbor by a minimum of 1 km and a maximum of 79 km, with a median of 13 km (IQR: 4–31 km).
We defined land use/land cover categories using a land cover classification scheme (Table 1) adapted from a U.S. Geological Survey classification scheme developed for remotely-sensed data (Anderson et al. 1976). We streamlined this scheme to eliminate matrix elements absent in our study area and more finely subdivided grassland categories (i.e., differentiating between native grasslands, marginal grasslands, restored grasslands, and CRP). We interpreted aerial photographs visually and digitized polygons of like land use/land cover.
To assess the spatial scale of landscape effects, we subdivided each of the 29 landscapes using five different buffers (500 m, 1, 2, 3, and 4 km). For each landscape and at each spatial scale we calculated landscape measures including the amount of each landscape element, the configuration of each landscape element, and overall measures of landscape structure.
We calculated percentage cover for each landscape element from the digitized landscape. The remaining landscape variables were calculated from a raster image. We converted each digitized aerial photograph to a raster image using ERDAS Imagine 2011 (Intergraph 2011) and used FRAGSTATS version 3.3 (McGarigal et al. 2002) to calculate landscape indices (Table 2). For each landscape element, we assessed the structure and arrangement of that element within the landscape using various measures (see Table 2). We also assessed indices of the aggregate landscape to determine if the songbirds were responding to the overall combination of the composition and configuration of the elements in the landscape. The aggregate landscape variables were divided into those associated with the composition of the landscape (types of land cover elements present: Habitat Richness, Habitat Diversity) and with the configuration of those different landscape element patches (the arrangement of patches within the landscape: Total Edge Density, Contagion). As the landscape sampled changed size with different sized focal patches, we normalized all landscape variables to avoid any potential bias.
Data analysis
We used multiple logistic regression in R 2.14.2 (R Development Core Team 2015) to relate species presence/absence to landscape and habitat characteristics. We only analyzed species found on 11–20 of the 29 sites (Table 3) to ensure that there was enough variability in the data to allow the model-fitting algorithm to be successful. The logistic regressions were completed within a multi-model framework (Burnham and Anderson 2002). Because of the large number of potential explanatory variables involved, we took a multi-pronged (local variables, matrix elements, and aggregate landscape variables) and multi-stage approach to the multi-model multiple logistic regression analysis (Fig. 2). Due to the fact that we had longer transects on larger prairie fragments there was a potential for the passive sampling effect to bias the presence data (Johnson and Igl 2001; Ribic et al. 2009b). We corrected for this by using transect length as an offset in all the logistic regression models (Faraway 2006). This process was repeated for each bird species analyzed.

For each species, occurrence was analyzed using multiple rounds of multi-model logistic regression analysis to identify the variables with the most statistical support at each level of measurement. For each round of analysis, variables with strong support (ΔAIC < 2) were retained and moved to the next round of analysis
The local variable branch of the analysis consisted of a single multi-model logistic regression analysis to identify the local variables with strong statistical support (defined throughout as those with ΔAICc < 2). The aggregate landscape branch of the analysis focused on those variables associated with overall landscape diversity and structural complexity and was conducted in two steps (Fig. 2). First, for each aggregate landscape variable we identified the spatial scale(s) with the most statistical support which were retained for the next step. Second, that pool of variables was combined in a multi-model multiple logistic regression analysis to determine the final set of aggregate landscape variables with the most statistical support. The third branch of the analysis (Fig. 2) focused on matrix element variables associated with the structure and amount of those individual matrix elements within the landscape. Because of the large number of variables in this branch, we used multiple rounds of analysis to narrow the pool of variables. As with the aggregate landscape variables, the first round was used to identify the important scale(s) for each variable. The most important variables were then identified for each matrix element, then for groupings of similar matrix elements (based on Level 1 classifications described in Table 1). We then used the variables from this round to build final matrix element models consisting of the best supported variables from all matrix element types.
We then incorporated the top variables from the local, aggregate landscape, and matrix element analyses into a single analysis to produce the set of best (ΔAICc < 2) logistic regression models including both landscape and local features. Finally, for each bird species we used the entire set of top models to calculate the deviances associated with each specific variable to determine their relative importance. For each variable within a model, the variable deviance was weighted by that of the model itself. Those weighted deviances were then averaged across all models for each variable to assess the relative importance of that variable.
Results
Of the 38 species encountered, nine—American Goldfinch (Carduelis tristis), Barn Swallow (Hirundo rustica), Grasshopper Sparrow (Ammodramus savannarum), Le Conte’s Sparrow (Ammodramus leconteii), Sedge Wren (Cistothorus platensis), Upland Sandpiper (Bartramia longicauda), Cliff Swallow (Hirundo pyrrhonota), Marsh Wren (Cistothorus palustris), and Western Meadowlark (Sturnells neglecta)—were found on between 11 and 20 field sites (Table 3) allowing for statistical analysis. The multi-model, logistic regression analysis resulted in multiple models for each bird species that explained 28–77% of patch occupancy deviance (Table 4).
Grasshopper Sparrows were more likely to be found on smaller remnant prairies with less brush cover and in landscapes with fewer CRP edges, greater herbaceous riparian patch density, higher matrix richness and diversity, and lower matrix contagion (Fig. 3A). Upland Sandpipers were more likely to be found on remnant prairies with less grass cover and more brush cover, and in landscapes with more CRP, larger patches of rural commercial land cover, more brush at 1 and 3 km and less brush at 2 and 4 km (Fig. 3B). Western Meadowlarks were more likely to be found on remnant prairies with fewer trees and in landscapes with greater matrix diversity, less matrix contagion, more dispersed CRP patches, and larger windbreak patches (Fig. 3C). Cliff Swallows were more likely to be found on larger remnant prairies with less forb cover, and in landscapes with lower matrix richness, lower matrix contagion, fewer brush patch edges, and lower hay patch density (Fig. 3D). Le Conte’s Sparrows were more likely to be found on prairie remnants with sparser vegetation and less open water, and in landscapes with higher matrix contagion, and more CRP edges (Fig. 3E). Marsh Wrens were more likely to be found on prairie remnants with more wetlands and trees, and in landscapes with lower CRP edge density, more wetland cover, fewer wetland patches, and smaller patches of open water (Fig. 3F). Sedge Wrens were more likely to be found on prairie remnants with less wetlands and less open water, and in landscapes with more total edge density, higher hay patch density and more dispersed hay patches, and greater wetland patch density (Fig. 3G). American Goldfinches were more likely to be found on prairie remnants with less forb cover, and in landscapes with higher matrix diversity, more dispersed forested riparian and windbreak patches, fewer hay patches, and more rural residential cover (Fig. 3H). Finally, Barn Swallows were more likely to be found on prairie remnants with less grass cover and denser vegetation, and in landscapes with higher matrix diversity, less brush cover, smaller hay patches, and less windbreak cover arranged with lower patch density and fewer patch edges (Fig. 3I).

Proportion of deviance explained by the components of the average model constructed from all top models (those with ΔAIC > 2) for each bird species (Panels A–I). The ± above each bar indicates if the term had a positive or negative effect on species presence. Model components are grouped into those associated with the prairie patch (grey bars), those associated with aggregate landscape measures (white bars), and those associated with specific landscape matrix elements (black bars). Variable names are from Table 2. For all landscape variables measurement scale (in km) is indicated by the number at the end of the variable name. For the matrix element variables, the matrix element variable name is from Table 1 and measurement variable name is from Table 2; for example, Pas ED 0.5 refers to edge density of pasture patches within 0.5 km of the prairie patch edge
Presence of eight of the nine species was most strongly influenced by landscape matrix element variables, while Cliff Swallow responded most strongly to local variables (Fig. 4). Among landscape variables, measures of configuration explained substantially more deviance in presence than measures of composition for seven species, while measures of composition and configuration were about equal for Cliff Swallow and measures of composition explained more deviance in presence for Upland Sandpiper (Fig. 5). Five out of nine species had the largest amount of deviance in presence explained by landscape variables measured at the highest spatial scales (3 and 4 km), the Western Meadowlark had an equal amount of deviance explained by large and small spatial scales, and the three remaining species (Cliff Swallow, Sedge Wren, and Marsh Wren) had the most deviance in presence explained by landscape variables measured at the smallest spatial scales (0.5 and 1 km; Fig. 6).

Proportion of deviance explained in the top models partitioned between local, aggregate landscape, and matrix element variables. Species codes as in Table 3. Habitat preference for species indicated: grassland obligates, grassland users, wetland users, tree associated, and human associated

Proportion of deviance explained by landscape variables (both aggregate landscape and matrix elements) in the top models partitioned between measures of landscape composition and configuration (see Table 2 for lists of variables in each group). Species codes as in Table 3. Habitat preference for species indicated: grassland obligates, grassland users, wetland users, tree associated, and human associated

Proportion of deviance explained by landscape variables (both aggregate landscape and matrix elements) in the top models partitioned between the spatial scales investigated. Species codes as in Table 3. Habitat preference for species indicated: grassland obligates, grassland users, wetland users, tree associated, and human associated
There was no clear relationship between the relative importance of local vs landscape variables and habitat guild, although the species with the strongest influences of local variables were all grassland obligates or users. Two grassland obligate species and one of the two grassland users responded most strongly to landscape variables measured at the 4 km scale. The type of variables that had the biggest effect on presence varied by habitat guild. Grassland obligates and grassland users responded primarily to CRP, hay, and brush with most species responding to patch arrangement (e.g., measures of edge, spacing, or patch density) as opposed to amount. The wetland users responded most strongly to patch density of water-based matrix elements.
Discussion
Individual species responses
The presence of Grasshopper Sparrows on a focal patch was strongly influenced by landscape edges. Edge sensitivity has been documented in this species (DeLisle and Savidge 1996; Helzer and Jelinski 1999), including a negative effect of shrubs (Whitmore 1981; Grant et al. 2004; Sutter and Ritchison 2005).
Local patch variables that influence nesting and foraging abilities played the largest role in predicting prairie remnant use by Upland Sandpipers. This species requires bare ground for nesting and feeding (Houston et al. 2011) explaining the negative effect of grass cover on sandpiper presence. Remnant prairie patches are more likely to be used by sandpipers only in landscapes with large enough grassland areas to provide both the bare ground and the dense vegetation needed for birds to feed and nest (Fritcher et al. 2004) which seem to be provided by CRP grasslands.
Western Meadowlarks were strongly influenced by the amount of edge in the landscape surrounding remnant prairies (as indicated by patch size and contagion measures). Western Meadowlarks are both edge (Bock et al. 1999) and area sensitive (Helzer and Jelinski 1999; Johnson and Igl 2001), although area sensitivity may be due to edge sensitivities (Johnson and Igl 2001; Fletcher 2005), and this edge sensitivity likely explains the importance of edges in the landscape for meadowlark occurrence. Locally, the amount of tree cover on the prairie fragment decreased the probability of Meadowlark occurrence, probably due to increased predation risk (Johnson and Temple 1990; Conover et al. 2011).
Unlike most other species in this study, deviance in Cliff Swallow presence was almost entirely explained by local variables. This species prefers open spaces conducive to aerial foraging (Ehrlich et al. 1988; Brown and Brown 1995) which are more likely to occur on larger prairie patches with less forb coverage. Further, insect prey availability seems to be an important determinant of Cliff Swallow colony location and size (Brown et al. 2002) and is likely affected by the size and quality of the prairie patch.
We found Le Conte’s Sparrow were more likely to be present on prairie patches embedded in landscapes with more CRP edges. This species breeds in both native prairie and CRP fields (Igl and Johnson 1995, 1999; Lowther 2005) though is potentially edge avoiding (Johnson and Igl 2001). Interestingly, Le Conte’s Sparrow responded positively to CRP edge density in the surrounding landscape, suggesting that habitat patch edge sensitivity may not extend to the overall landscape structure.
The probability of Marsh Wrens occurring on a prairie patch increased with greater wetland area in the surrounding landscape, as would be expected from a wetland associated species (Cunningham and Johnson 2006; Spautz et al. 2006; Kroodsma and Verner 2014). However, the probability of being present decreased with increasing wetland patch density suggesting that landscapes with highly fragmented wetlands were not as attractive to Marsh Wrens. This is likely due to edge sensitivities (Fairbairn and Dinsmore 2001; Spautz et al. 2006). High edge sensitivity also explains our finding that high CRP edge density close to the prairie patch also decreased the probability of Marsh Wren being present.
In contrast to Marsh Wrens, Sedge Wrens were more likely to be present if the landscape had higher densities of wetland and hay patches. Sedge Wrens rely on wet meadows and seasonal wetlands to provide preferred sedge and grass seeds (Herkert et al. 2001). More wetland patches in the landscape likely increases the availability of wet meadow habitat (Fairbairn and Dinsmore 2001; Riffell et al. 2001; Bakker et al. 2002).
As an edge species, American Goldfinch (Herkert 1994; Horn et al. 2002) rely on shrubs and trees for nesting and cover (Stokes 1950; Middleton 1979; McGraw and Middleton 2009; personal observations). At our study sites, trees and shrubs were mostly found in the windbreaks between fields and, to a lesser extent, in riparian buffers, and landscapes with more evenly spaced windbreaks and forested riparian buffers were more likely to have goldfinches. Monotypic hayfields do not contain enough seeds for foraging (McGraw and Middleton 2009) and the presence of hay fields actually reduces the probability of goldfinch occurrence.
Similar to the American Goldfinch, Barn Swallow presence was driven almost entirely by landscape variables. This species is considered a generalist, using a wide variety of habitats and feeding on many different types of prey (Turner 2006), and preferentially uses edges that concentrate or increase prey availability (Evans et al. 2003). Landscapes with smaller hay patches were more likely to provide such foraging opportunities. The tall vertical structure of windrows can interfere with foraging (Henderson et al. 2007) and so landscapes with more windrows were less likely to have swallows.
Landscape effects
For eight of the nine species studied, the landscape variables explained much more of the deviation in patch occupancy than the local patch characters did. Prior studies of grassland birds have had mixed results with some detecting landscape effects (Söderström and Pärt 2000; Ribic and Sample 2001; Bakker et al. 2002; Hamer et al. 2006; Winter et al. 2006; Renfrew and Ribic 2008), others finding little to no effect of the landscape (Bajema and Lima 2001; Horn et al. 2002; Koper and Schmiegelow 2006; Jacobs et al. 2012), and yet others finding the combination of local and landscape variables having the greatest explanatory power (Fletcher and Koford 2002; Cunningham and Johnson 2006; Quamen 2007). Reviews of landscape studies have found landscape effects in less than 80% of bird-focused studies (Mazerolle and Villard 1999) and that bird studies were least likely to demonstrate landscape effects, even though birds were one of the most frequently studied taxa (Thornton et al. 2011). Some of the ambiguity in the importance of landscape variables is likely due to methodological approaches, particularly how the landscape data were gathered. Studies that simply buffer from the bird sampling points (e.g., Best et al. 2001; Coppedge et al. 2001; Bakker et al. 2002; Fletcher and Koford 2002; Winter et al. 2006) would potentially mix local and landscape data together obscuring the importance of the landscape. Furthermore, many studies tended to measure landscape data on smaller scales (2 km or less) than we did (e.g., Bergin et al. 2000; Bajema and Lima 2001; Ribic and Sample 2001; Grant et al. 2004; Jacobs et al. 2012) and we saw strong bird responses to landscape variables measured 3–4 km beyond the edge of the prairie patch, suggesting studies measuring landscape variables at smaller scales might miss landscape effects on bird occurrence. The design of our study (a focal patch approach that clearly delineated local vs landscape effects and a large extent for the landscape data) allowed us to detect landscape influences on bird occupancy of remnant prairie patches.
Most of the species we analyzed seemed more sensitive to the configuration of elements in the surrounding landscape, rather than the composition of the landscape. One of the striking patterns across the species in this study is the prevalence of responses to habitat edges. Seven of the nine species responded to at least one edge measurement (either edge density or contagion), and three species (Grasshopper Sparrow, LeConte’s Sparrow, Cliff Swallow) responded to more than one edge variable, although the type and magnitude varied significantly between species. Interestingly, only one species responded to total edge density (Sedge Wren), indicating that the specific edge type might be much more important than estimated by previous research (DeLisle and Savidge 1996; Winter et al. 2000; Jensen and Finck 2004; Koper et al. 2009).
The importance of the surrounding landscape, particularly at the larger scales that we observed, match with the hierarchical approach that many bird species use to choose breeding territories (Reed et al. 1999). As birds return in the spring to find new nesting grounds for the year, they first look for specific matrix elements at broad scales. These matrix elements can either be avoided, as Grasshopper Sparrows avoid areas with high CRP edge density, or targeted, as is seen with Upland Sandpipers that choose regions with higher amounts of CRP. Once the migrating birds have selected a region they are going to settle in, focal-patch level characteristics become important, including the relationships with woody vegetation and vegetation structure that have been well documented previously (Whitmore 1981; Davis et al. 1999; Winter et al. 2005; Jacobs et al. 2012).
There was little influence of habitat preference of the bird on relationships between patch occupancy and landscape characters. Grassland obligates and grassland users were slightly more likely to have some influence from local variables, with one grassland user (Cliff Swallow) being the only species to have local variables as important as landscape variables. Given the demonstrated importance of habitat characteristics for these grassland users (Whitmore 1981; Davis et al. 1999; Winter et al. 2005; Jacobs et al. 2012) it is not surprising that these two groups of birds are more likely to have a stronger influence of local variables. It is worth pointing out that even so, for 4 of the 5 species in these two groups, landscape variables were more important than local variables. The only other pattern with guild was that the wetland users responded to landscape variables at shorter distances than birds from all the other guilds, with the exception of the grassland user the Cliff Swallow. Despite these weak patterns across habitat preference groups, it seems that each species responds to the landscape in a unique manner dictated by the specifics of its life history, behavioral responses to landscape elements, and movement capabilities.
Our findings have implications for conservation management for grassland songbirds. While it is true that songbird management can only occur on specific parcels of land (like the focal patch) rather than at the entire landscape scale, understanding the landscape context around the focal patch can help to identify songbird populations located in less-hospitable landscapes, which may be in need of local habitat improvements that would provide population support. As such, future species management plans should include an understanding of the landscape context out to at least 4 km, if not further. Efforts should also be made to include details about matrix element configuration and edge type rather than area only (particularly for those matrix elements that provide sharp contrasts to grassland structure). Plans targeted at species with very specific or limiting habitat requirements should also include information about landscape composition, with specific attention being paid to matrix elements that either complement those requirements or make them harder to be met.
References
Ahlering MA, Merkord CL (2016) Cattle grazing and grassland birds in the northern tallgrass prairie. J Wildl Manag 80(4):643–654
Anderson BW, Ohmart RD (1981) Comparisons of avian census results using variable distance transect and variable circular plot techniques. Stud Avian Biol 6:186–192
Anderson JR, Hardy EE, Roach JT, Witmer RE (1976) A land use and land cover classification system for use with remote sensor data. United States Government Printing Office, Washington, DC
Askins RA, Cháívez-Ramírez F, Dale BC, Haas CA, Herkert JR, Knopf FL, Vickery PD (2007) Conservation of grassland birds in North America: understanding ecological processes in different regions. Ornithol Monogr 64:1–46
Bajema RA, Lima SL (2001) Landscape-level analysis of Henslow’s Sparrow (Ammodramus henslowii) abundance in reclaimed coal mine grasslands. Am Midl Nat 145(2):288–298
Baker-Gabb D, Antos M, Brown G (2016) Recent decline of the critically endangered Plains-wanderer (Pedionomus torquatus), and the application of a simple method for assessing its cause: major changes in grassland structure. Ecol Manag Restor 17(3):235–242
Bakker KK, Naugle DE, Higgins KF (2002) Incorporating landscape attributes into models for migratory grassland bird conservation. Conserv Biol 16(6):1638–1646
Bender DJ, Contreras TA, Fahrig L (1998) Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79(2):517–533
Bergin TM, Best LB, Freemark KE, Koehler KJ (2000) Effects of landscape structure on nest predation in roadsides of a midwestern agroecosystem: a multiscale analysis. Landscape Ecol 15(2):131–143
Best LB, Bergin TM, Freemark KE (2001) Influence of landscape composition on bird use of rowcrop fields. J Wildl Manag 65(3):442–449
Bibby CJ, Burgess ND, Hill DA (1992) Bird census techniques. Academic Press, San Diego, CA
Bock CE, Bock JH, Bennett BC (1999) Songbird abundance in grasslands at a suburban interface on the Colorado high plains. Stud Avian Biol 19:131–136
Borgmann KL, Rodewald AD (2004) Nest predation in an urbanizing landscape: the role of exotic shrubs. Ecol Appl 14(6):1757–1765
Brennan JM, Bender DJ, Contreras TA, Fahrig L (2002) Focal patch landscape studies for wildlife management: optimizing sampling effort across scales. In: Liu J, Taylor WW (eds) Integrating landscape ecology into natural resource management. Cambridge University Press, Cambridge, pp 68–91
Brown CR, Brown MB (1995) Cliff Swallow (Petrochelidon pyrrhonota). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/149
Brown CR, Sas CM, Brown MB (2002) Colony choice in Cliff Swallows: effects of heterogeneity in foraging habitat. Auk 119(2):446–460
Burnham KP, Anderson DR (2002) Model selection and inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Conover RR, Wes Burger L, Linder ET (2011) Grassland bird nest ecology and survival in upland habitat buffers near wooded edges. Wildl Soc Bull 35(4):353–361
Coppedge BR, Engle DM, Masters RE, Gregory MS (2001) Avian response to landscape change in fragmented southern great plains grasslands. Ecol Appl 11(1):47–59
Cunningham MA, Johnson DH (2006) Proximate and landscape factors influence grassland bird distributions. Ecol Appl 16(3):1062–1075
Davis SK (2005) Nest-site selection patterns and the influence of vegetation on nest survival of mixed-grass prairie passerines. Condor 107(3):605–616
Davis SK, Brittingham M (2004) Area sensitivity in grassland passerines: effects of patch size, patch shape, and vegetation structure on bird abundance and occurrence in southern Saskatchewan. Auk 121(4):1130–1145
Davis SK, Duncan DC, Skeel M (1999) Distribution and habitat associations of three endemic grassland songbirds in southern Saskatchewan. Wilson Bull 111(3):389–396
Deák B, Valkö O, Török P, Thmérész B (2016) Factors threatening grassland specialist plants—a multi-proxy study on the vegetation of isolated grasslands. Biol Conser 204:255–262
DeLisle JM, Savidge JA (1996) Reproductive success of Grasshopper Sparrows in relation to edge. Prairie Nat 28(3):107–113
Duggan JM, Schooley RL, Heske EJ (2011) Modeling occupancy dynamics of a rare species, Franklin’s ground squirrel, with limited data: are simple connectivity metrics adequate? Landscape Ecol 26(10):1477–1490
Ehrlich PR, Dobkin DS, Wheye D (1988) The Birder’s Handbook: a field guide to the natural history of North American birds. Simon and Schuster Inc., New York
Evans KL, Bradbury RB, Wilson JD (2003) Selection of hedgerows by Swallows Hirundo rustica foraging on farmland: the influence of local habitat and weather. Bird Study 50(1):8–14
Eycott AE, Stewart GB, Buyung-Ali LM, Bowler DE, Watts K, Pullin AS (2012) A meta-analysis on the impact of different matrix structures on species movement rates. Landscape Ecol 27(9):1263–1278
Fairbairn SE, Dinsmore JJ (2001) Local and landscape-level influences on wetland bird communities of the prairie pothole region of Iowa, USA. Wetlands 21(1):41–47
Faraway JJ (2006) Extending the linear model with R: generalized linear, mixed effects and nonparametric regression models. Chapman & Hall/CRC, Boca Raton
Fletcher RJ (2005) Multiple edge effects and their implications in fragmented landscapes. J Anim Ecol 74(2):342–352
Fletcher RJ, Koford RR (2002) Habitat and landscape associations of breeding birds in native and restored grasslands. J Wildl Manag 66(4):1011–1022
Fontana CS, Dotta G, Marques CK, Repenning M, Agne CA, dos Santos RJ (2016) Conservation of grassland birds in South Brazil: a land management perspective. Nat Conserv 14(2):83–87
Fritcher SC, Rumble MA, Flake LD (2004) Grassland bird densities in seral stages of mixed-grass prairie. J Range Manag 57(4):351–357
Fuhlendorf SD, Engle DM, Kerby J, Hamilton R (2008) Pyric herbivory: rewilding landscapes through the recoupling of fire and grazing. Conserv Biol 23(3):588–598
Gates CE (1981) Optimizing sampling frequency and numbers of transects and stations. Stud Avian Biol 6:399–404
Gebauer K, Dickinson KJM, Whigham PA, Seddon PJ (2013) Matrix matters: differences of grand skink metapopulation parameters in native tussock grasslands and exotic pasture grasslands. PLoS ONE 8(10):e76076
Gibbons DW, Hill DA, Sutherland WJ (1996) Birds. In: Sutherland WJ (ed) Ecological census techniques: a handbook. Cambridge University Press, Cambridge, pp 227–259
Grant TA, Madden E, Berkey GB (2004) Tree and shrub invasion in northern mixed-grass prairie: implications for breeding grassland birds. Wildl Soc Bull 32(3):807–818
Haas CA (1995) Dispersal and use of corridors by birds in wooded patches on an agricultural landscape. Conserv Biol 9(4):845–854
Hamer TL, Flather CH, Noon BR (2006) Factors associated with grassland bird species richness: the relative roles of grassland area, landscape structure, and prey. Landscape Ecol 21(4):569–583
Hansen AJ, Urban DL (1992) Avian response to landscape pattern: the role of species’ life histories. Landscape Ecol 7(3):163–180
Helzer CJ, Jelinski DE (1999) The relative importance of patch area and perimeter-area ratio to grassland breeding birds. Ecol Appl 9(4):1448–1458
Henderson I, Holt C, Vickery J (2007) National and regional patterns of habitat association with foraging barn swallows Hirundo rustica in the UK. Bird Study 54(3):371–377
Herkert JR (1994) The effects of habitat fragmentation on Midwestern grassland bird communities. Ecol Appl 4(3):461–471
Herkert JR, Kroodsma DE, Gibbs JP (2001) Sedge Wren (Cistothorus platensis). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. http://bna.birds.cornell.edu/bna/species/582
Hoekstra JM, Boucher TM, Ricketts TH, Roberts C (2005) Confronting a biome crisis: global disparities of habitat loss and protection. Ecol Lett 8(1):23–29
Horn DJ, Koford RR, Braland ML (2002) Effects of field size and landscape composition on grassland birds in south-central Iowa. J Iowa Acad Sci 109:1–7
Houston CS, Jackson CR, Bowen DE (2011) Upland Sandpiper (Bartramia longicauda). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. http://bna.birds.cornell.edu/bna/species/580
Hutto RL (1985) Habitat selection by nonbreeding, migratory land birds. In: Cody ML (ed) Habitat selection in birds. Academic Press, Orlando, pp 455–476
Igl LD, Johnson DH (1995) Dramatic increase of Le Conte’s Sparrow in Conservation Reserve Program fields in the northern Great Plains. Prairie Nat 27(2):89–94
Igl LD, Johnson DH (1997) Changes in breeding bird populations in North Dakota: 1967 to 1992-93. Auk 114(1):74–92
Igl LD, Johnson DH (1999) Le Conte’s Sparrows breeding in conservation reserve program fields: precipitation and patterns of population change. Stud Avian Biol 19:178–186
Intergraph (2011) ERDAS Imagine Version 11.0, Madison, AL
Jacobs RB, Thompson FR, Koford RR, la Sorte FA, Woodward HD, Fitzgerald JA (2012) Habitat and landscape effects on abundance of Missouri’s grassland birds. J Wildl Manag 76(2):372–381
Jensen WE, Finck EJ (2004) Edge effects on nesting Dickcissels (Spiza americana) in relation to edge type of remnant tallgrass prairie in Kansas. Am Midl Nat 151(1):192–199
Johnson DH (2000) Grassland bird use of Conservation Reserve Program fields in the Great Plains. In: Heard LP, Allen AW, Best LB, Brady SJ, Burger W, Esser AJ, Hackett E, Johnson DH, Pederson RL, Reynolds RE, Rewa C, Ryan MR, Molleur RT, Buck P (eds) A comprehensive review of farm bill contributions to wildlife conservation, 1985-2000. USDA/NRCS/WHMI, Jamestown, pp 19–33
Johnson DH, Igl LD (2001) Area requirements of grassland birds: a regional perspective. Auk 118(1):24–34
Johnson RG, Temple SA (1990) Nest predation and brood parasitism of tallgrass prairie birds. J Wildl Manag 54(1):106–111
Kalinowski RS, Johnson MD (2010) Influence of suburban habitat on a wintering bird community in coastal northern California. Condor 112(2):274–282
Koper N, Manseau M (2009) Generalized estimating equations and generalized linear mixed-effects models for modelling resource selection. J Appl Ecol 46(3):590–599
Koper N, Schmiegelow FKA (2006) A multi-scale analysis of avian response to habitat amount and fragmentation in the Canadian dry mixed-grass prairie. Landscape Ecol 21(7):1045–1059
Koper N, Walker DJ, Champagne J (2009) Nonlinear effects of distance to habitat edge on Sprague’s pipits in southern Alberta, Canada. Landscape Ecol 24(10):1287–1297
Kroodsma DE, Verner J (2014) Marsh wren (Cistotorus palustris). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. http://bna.birds.cornell.edu/bna/species/308
Lowther PE (2005) Le Conte’s sparrow (Ammodramus leconteii). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. http://bna.birds.cornell.edu/bna/species/224
Mazerolle MJ, Villard M-A (1999) Patch characteristics and landscape context as predictors of species presence and abundance: a review. Ecoscience 6(1):117–124
McGarigal K, Cushman SA, Ene E (2002) FRAGSTATS V3: spatial pattern analysis program for categorical maps
McGraw KJ, Middleton AL (2009) American Goldfinch (Spinus tristis). In: Poole A (ed) The birds of North America online. Cornell Lab of Ornithology, Ithaca. http://bna.birds.cornell.edu/bna/species/080
Middleton ALA (1979) Influence of age and habitat on reproduction by the American Goldfinch. Ecology 60(2):418–432
Moranz RA, Fuhlendorf SD, Engle DM (2014) Making sense of a prairie butterfly paradox: the effects of grazing, time since fire, and sampling period on regal fritillary abundance. Biol Conserv 173:32–41
North American Bird Conservation Initiative Canada (2012) The state of Canada’s birds, 2012. Environment Canada, Ottawa
O’Leary CH, Nyberg DW (2000) Treelines between fields reduce the density of grassland birds. Nat Areas J 20(3):243–249
Otto CRV, Roth CL, Carlson BL, Smart MD (2016) Land-use change reduces habitat suitability for supporting managed honey bee colonies in the Northern Great Plains. Proc Natl Acad Sci USA 113(37):10430–10435
Patten MA, Shochat E, Reinking DL, Wolfe DH, Sherrod SK (2006) Habitat edge, land management, and rates of brood parasitism in tallgrass prairie. Ecol Appl 16(2):687–695
Piqueray J, Bisteau E, Cristofoli S, Palm R, Poschold P, Mahy G (2011) Plant species extinction debt in a temperate biodiversity hotspot: community, species and functional traits approaches. Biol Conserv 144(5):1619–1629
Poniatowski D, Löffler F, Stuhldreher G, Borchard F, Krämer B, Fartmann T (2016) Functional connectivity as an indicator for patch occupancy in grassland specialists. Ecol Ind 67:735–742
Prevedello JA, Vieria MV (2010) Does the type of matrix matter? A quantitative review of the evidence. Biodivers Conserv 19(5):1205–1223
Quamen FR (2007) A landscape approach to grassland bird conservation in the Prairie Pothole Region of the Northern Great Plains. Ph.D. thesis, University of Montana, pp 150
R Development Core Team (2015) R: a language and environment for statistical computing, Vienna
Reed JM, Boulinier T, Danchin E, Oring LW (1999) Informed dispersal: prospecting by birds for breeding sites. Curr Ornithol 15:189–259
Reif J (2013) Long-term trends in bird populations: a review of patterns and potential drivers in North America and Europe. Acta Ornithol 48(1):1–16
Reino L, Beja P, Araújo MB, Dray S, Segurado P (2012) Does local habitat fragmentation affect large-scale distributions? The case of a specialist grassland bird. Divers Distrib 19(4):423–432
Renfrew RB, Ribic CA (2008) Multi-scale models of grassland passerine abundance in a fragmented system in Wisconsin. Landscape Ecol 23(2):181–193
Ribic CA, Guzy MJ, Sample DW (2009a) Grassland bird use of remnant prairie and Conservation Reserve Program fields in an agricultural landscape in Wisconsin. Am Midl Nat 161(1):110–121
Ribic CA, Korford RR, Herkert JR, Johnson DH, Niemuth ND, Naugle DE, Bakker KK, Sample DW, Renfrew RB (2009b) Area sensitivity in North American grassland birds: patterns and processes. Auk 126(2):233–244
Ribic CA, Sample DW (2001) Associations of grassland birds with landscape factors in southern Wisconsin. Am Midl Nat 146(1):105–121
Riffell SK, Keas BE, Burton TM (2001) Area and habitat relationships of birds in Great Lakes coastal wet meadows. Wetlands 21(4):492–507
Robel RJ, Briggs JN, Dayton AD, Hulbert LC (1970) Relationships between visual obstruction measurements and weight of grassland vegetation. J Range Manag 23(4):295–297
Rodewald AD (2003) The importance of land uses within the landscape matrix. Wildl Soc Bull 31(2):586–592
Rotem G, Gavish Y, Shacham B, Giladi I, Bouskila A, Ziv Y (2016) Combined effects of climatic gradient and domestic livestock grazing on reptile community structure in a heterogeneous agroecosystem. Oecologia 180(1):231–242
Samson FB, Knopf FL (1994) Prairie conservation in North America. Bioscience 44(6):418–421
Samson FB, Knopf FL, Ostlie WR (2004) Great plains ecosystems: past, present, and future. Wildl Soc Bull 32(1):6–15
Sauer JR, Link WA (2011) Analysis of the North American breeding bird survey using hierarchical models. Auk 128(1):87–98
Söderström B, Pärt T (2000) Influence of landscape scale on farmland birds breeding in semi-natural pastures. Conserv Biol 14(2):522–533
Spautz H, Nur N, Stralberg D, Chan Y (2006) Multiple-scale habitat relationships of tidal-marsh breeding birds in the San Francisco Bay estuary. Stud Avian Biol 32:247–269
Stokes AW (1950) Breeding behavior of the goldfinch. Wilson Bulletin 62(3):107–127
Sutter B, Ritchison G (2005) Effects of grazing on vegetation structure, prey availability, and reproductive success of grasshopper sparrows. J Field Ornithol 76(4):345–351
Thornton DH, Branch LC, Sunquist ME (2011) The influence of landscape, patch, and within-patch factors on species presence and abundance: a review of focal patch studies. Landscape Ecol 26(1):7–18
Turner AK (2006) The barn swallow. Poyser, London
Whitmore RC (1981) Structural characteristics of Grasshopper Sparrow habitat. J Wildl Manag 45(3):811–814
Winter M, Johnson DH, Faaborg J (2000) Evidence for edge effects on multiple levels in tallgrass prairie. Condor 102(2):256–266
Winter M, Johnson DH, Shaffer JA, Donovan TM, Svedarsky WD (2006) Patch size and landscape effects on density and nest success of grassland birds. J Wildl Manag 70(1):158–172
Winter M, Shaffer JA, Johnson DH, Donovan TM, Svedarsky WD, Jones PW, Euliss BR (2005) Habitat and nesting of Le Conte’s Sparrows in the northern tallgrass prairie. J Field Ornithol 76(1):61–71
With KA, King AW, Jensen WE (2008) Remaining large grasslands may not be sufficient to prevent grassland bird declines. Biol Conserv 141(12):3152–3167
Wright CK, Wimberly MC (2013) Recent land use change in the Western Corn Belt threatens grasslands and wetlands. Proc Natl Acad Sci USA 110(10):4134–4139
Acknowledgements
This manuscript was greatly improved by the input of Douglas H. Johnson and two anonymous reviewers. We thank the Nature Conservancy, Minnesota Department of Natural Resources, U.S. Fish and Wildlife Service, and the University of North Dakota for access to the prairie fragments under their management. This work was supported by funding from the Minnesota Ornithologists’ Union, North Dakota View, the University of North Dakota Intercollegiate Academics Fund, and the Esther Wadsworth Wheeler Award.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Shahan, J.L., Goodwin, B.J. & Rundquist, B.C. Grassland songbird occurrence on remnant prairie patches is primarily determined by landscape characteristics. Landscape Ecol 32, 971–988 (2017). https://doi.org/10.1007/s10980-017-0500-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-017-0500-4