
Abstract
Context
Interactions among disturbances, climate, and vegetation influence landscape patterns and ecosystem processes. Climate changes, exotic invasions, beetle outbreaks, altered fire regimes, and human activities may interact to produce landscapes that appear and function beyond historical analogs.
Objectives
We used the mechanistic ecosystem-fire process model FireBGCv2 to model interactions of wildland fire, mountain pine beetle (Dendroctonus ponderosae), and white pine blister rust (Cronartium ribicola) under current and future climates, across three diverse study areas.
Methods
We assessed changes in tree basal area as a measure of landscape response over a 300-year simulation period for the Crown of the Continent in north-central Montana, East Fork of the Bitterroot River in western Montana, and Yellowstone Central Plateau in western Wyoming, USA.
Results
Interacting disturbances reduced overall basal area via increased tree mortality of host species. Wildfire decreased basal area more than beetles or rust, and disturbance interactions modeled under future climate significantly altered landscape basal area as compared with no-disturbance and current climate scenarios. Responses varied among landscapes depending on species composition, sensitivity to fire, and pathogen and beetle suitability and susceptibility.
Conclusions
Understanding disturbance interactions is critical for managing landscapes because forest responses to wildfires, pathogens, and beetle attacks may offset or exacerbate climate influences, with consequences for wildlife, carbon, and biodiversity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Interactions among climate, disturbances, and vegetation determine landscape patterns and influence ecosystem processes. Disturbances disrupt ecosystems, communities, and population structures and change elements of the biological and physical environment (Turner 2010). Disturbance creates and maintains biological diversity, which is generally highest at intermediate post-successional stages. Too little disturbance results in competitive exclusion of some species, while too much disturbance eliminates species lacking ability to rapidly re-colonize (Grime 1973). Climate changes may be altering characteristics of and interactions among disturbance agents, events, and regimes with potential for non-linear feedbacks and novel and unanticipated landscape responses (Turner 2010; Temperli et al. 2013; Buma 2015) and profound shifts in successional pathways, species composition, and landscape carbon (Goetz et al. 2007; Johnstone et al. 2010a, b); Brown and Johnstone 2012.
Disturbance regimes are described by characteristics of frequency, intensity, size, and pattern (Simard 1991; Agee 1993; Keane et al. 2015a). Climate changes can alter the timing, magnitude, frequency, and duration of disturbance events, as well as interactions with vegetation and other disturbances (Fig. 1). Direct and indirect interactions among disturbances can result in highly visible, rapidly occurring, and persistent changes in landscape composition and structure. The importance of these interactions has been shown in studies across the world. In the western US, Miller and Wigand (1994) and Bachelet et al. (2000) describe woodland expansion as the result of reduced fire frequency associated with livestock grazing, while Buma and Wessman (2012) showed that fire, windthrow, and salvage logging dictated landscape composition through individual species responses to interacting disturbances. Other research by Allen (2007) attributed forest dieback in New Mexico to the interactions of fire, grazing, erosion, and severe drought, and Beh et al. (2014) found that novel interactions between sudden oak death and wildfire contributed to the intensity of subsequent ambrosia beetle attacks in California tanoak forests. Hicke et al. (2012) and others have documented changes in fire behavior, extent, and severity result from bark beetle-caused mortality in pine forests, with possible amplification of fire-beetle interactions as climate changes increase potential for wide-spread beetle outbreaks (Bentz et al. 2010). In South America, Matson and Bart (2013) showed that the interaction of fire and grazing dictate shrub encroachment in the Andes. In recent work by Lewis and Lindgren (2002), interactions between Tomentosus spp. root disease and spruce beetle controlled mortality and volume lost in boreal forests of British Columbia, Canada, while the importance of drought, grazing, and fire interactions to the structure and composition of grasslands was documented by Koerner and Collins (2014) in South Africa and Hobbs et al. (2003) in Australia. Together, these findings highlight the importance of considering the interactive effects of multiple disturbances on vegetation and ecosystem processes.

Direct and indirect interactions among climate, vegetation, and four co-occurring disturbances of the western US
Exploration of the numerous, complex, and multi-scale interactions among ecological processes, disturbance agents, and climate drivers is difficult using traditional field methods that typically do not extend across large areas and long time periods—although it is precisely this landscape-scale and multi-decadal view that is required to assess if and when disturbance interactions may temporarily or persistently alter ecosystems (Keane et al. 2015b). Alternatively, disturbance interactions and subsequent effects can be modeled using landscape simulation models (Mladenoff 2004; Turner 2005). Here, we use the spatially-explicit ecosystem-fire simulation model FireBGCv2 (Keane et al. 2011; Loehman et al. 2011a; Holsinger et al. 2014; Riggs et al. 2015) to assess how climate changes and interacting disturbances alter landscapes across a range of ecological systems in the US northern Rocky Mountains (“Northern Rockies”). Unlike many process models, FireBGCv2 mechanistically represents interactions of climate drivers, abiotic and biotic disturbance, and vegetation from the scale of individual tree to landscape. The modeled suite of disturbance agents—wildland fire (any fire that occurs in a non-developed or sparsely developed area), mountain pine beetle (Dendroctonus ponderosae), and white pine blister rust (Cronartium ribicola)—co-occur across a broad area of the western United States. Acting independently or synchronously in space and time, wildland fires, mountain pine beetle outbreaks, and white pine blister rust infections can substantially influence forest structure, composition, and function at landscape scales and alter biogeochemical processes such as carbon cycling, water supply, and nutrient cycles (Fettig et al. 2007; Kurz et al. 2008a, b; Edburg et al. 2012).
Background
Wildland fire (fire)
Wildland fire is the most ubiquitous and dominant landscape disturbance throughout forest ecosystems of the Northern Rockies, and is responsive to variability in climate, weather, topography, and vegetation type and distribution (Schoennagel et al. 2004). Many western ecosystems are fire adapted and fire dependent. Beneficial ecological consequences of fire in these systems include nutrient release and redistribution, stimulation of plant growth, increased productivity in soil systems from decomposition of burned material, initiation of vegetation succession and forest regeneration, increased availability of resources for surviving trees, and creation of critical wildlife habitat (Loehman et al. 2014). Fire exclusion since the 1920s has increased surface fuel loads, tree densities, and ladder fuels, especially in low-elevation, dry conifer forests. As a result, fires here may be larger and more intense, and may cause higher rates of tree mortality, than historical fires. In higher elevation forests where fires were historically infrequent, fire exclusion has not significantly impacted fire regimes (Romme and Despain 1989; Schoennagel et al. 2004). Sustained increases in annual and summer temperatures and summer drying projected for the Northern Rockies (Gross et al. 2016) are expected to result in longer fire seasons (i.e., time for more fires to occur and burn for longer periods across larger areas) (Westerling et al. 2006). Earlier onset of snow melt will likely reduce fuel moistures, making a larger portion of the landscape flammable for longer periods of time (Brown et al. 2004), and prolonged droughts may result in widespread, regional fire years (Heyerdahl et al. 2008).
Mountain pine beetle
The principal agent of insect-caused mortality in Northern Rockies pine forests is the mountain pine beetle, D. ponderosae Hopkins. The mountain pine beetle is a native, cambial-feeding bark beetle that attacks ponderosa pine (Pinus ponderosa), western white pine (Pinus monticola), whitebark pine (Pinus albicaulis), limber pine (Pinus flexilus), and lodgepole pine (Pinus contorta) (Gibson et al. 2009; Safranyik et al. 2010). Tree defense against these beetles is complex, including both physical (e.g., resin flow) and chemical (e.g. terpenoid compounds) defenses, but beetles can overwhelm host defenses through sheer numbers with a mass attack strategy (Hood et al. 2015). Mountain pine beetle populations periodically erupt into large-scale outbreaks capable of causing partial or total tree mortality over large areas (Safranyik et al. 2010). Climate affects beetle dynamics directly—temperature governs beetle survival, development, and dispersal (Bentz et al. 2010)—and indirectly via controls on host tree establishment and growth. Coops et al. (2012) modeled areas of vulnerable host trees and areas of potential beetle expansion under future climate, suggesting that timing and location of future outbreaks will depend on complex interactions among climate-driven effects on tree distributions and tree stress, as well as independent effects on mountain pine beetle phenology and outbreak dynamics.
White pine blister rust
White pine blister rust, caused by the fungus C. ribicola, was introduced into the US from Europe around 1910 and infects only five-needle pines (Tomback and Achuff 2010). Its complex life cycle requires two hosts with two spore-producing stages on pines and three spore producing stages on Ribes spp. Basidiospores produced on Ribes spp have a narrow window for production and successful infection of pines, requiring warm temperatures (>20 °C) and high humidity (>98 %) (McDonald et al. 1981). The time required for blister rust to kill its host varies by species from 5 to 10 years for western white pine to over 20 years for whitebark pine (Hoff and Hagle 1990). White pine blister rust susceptibility and resistance are highly variable at local and regional scales, but in some areas rust-caused mortality is as high as 90 % (Mahalovich et al. 2006; Schoettle and Sniezko 2007). Keane et al. (2012) note that severe fires that kill rust-resistant pines ensure continued high white pine blister rust infection rates in existing and new cohorts. However, where rust-resistant five-needle pines survive fire they can provide a seed source for repopulating burned areas with high densities of rust-resistant seedlings (Leirfallom et al. 2015).
Disturbance interactions
Few studies have investigated interactive effects of mountain pine beetle, white pine blister rust, and fire disturbances on forests, particular in response to projected climate changes (Keane et al. 2015a). Fire-beetle interactions have been fairly widely investigated, principally with regard to changes in fire behavior, extent, and severity resulting from bark beetle-caused mortality in pine forests (see Hicke et al. 2012 for a summary), but the reciprocal interactions of beetles and fire, particularly through several disturbance cycles, are not well characterized [but see Parker et al. (2006)]. Interactions of mountain pine beetle and white pine blister rust are governed by climatic (e.g., warming temperatures outside host tree physiological tolerance) and biophysical (e.g., topographic setting, stand structure and composition, wildfire patterns) factors that influence the distribution and density of whitebark pines on the landscape (Larson 2011; Hansen et al. 2016). Observationally, the greatest declines in whitebark pine have been found in areas that have experienced blister rust and beetle outbreaks but not fire (Campbell and Antos 2000), possibly because fire, while reducing pines in the short term, can ensure their long-term persistence by eliminating competitors (Keane and Morgan 1994).
Methods
FireBGCv2 simulation model
FireBGCv2 (Fire BioGeoChemical model Version 2) is a spatially explicit, mechanistic fire and forest succession model that links many cross-scale interactions in the simulation of ecosystem processes (see Keane et al. 2011 for complete model documentation). FireBGCv2 simulates key processes such as tree and understory growth, tree regeneration and mortality, litterfall, and aboveground organic matter decomposition using detailed physical and biogeochemical relationships. Wildfire, mountain pine beetle and white pine blister rust activity occur in response to dynamic properties of fuelbeds (fire), tree species and structural traits and landscape composition (beetles and rust), and weather (all disturbance processes). Wildfires ignite when fine fuels are adequate (>0.05 kg m−2) and sufficiently dry (moisture contents <10 %), with probability and origin of ignition determined by input fire return intervals. Fire spread is modeled using slope and wind vectors, fuel characteristics, and fuel moistures, fire behavior characteristics such as flame length, scorch height, and intensity are derived from submodel model that calculates heat transfer to fuels (Albini 1976) and fire-caused tree mortality is modeled as a function of input, individual-tree bark thickness and scorch height. Blister rust infections occur in five-needle pines when daily relative humidity is above 90 % and daily mean temperature is above 10 °C, and tree mortality is simulated using empirical relationships of time since infection and tree diameter. Mountain pine beetle outbreaks are initiated when lethal temperature thresholds (below −40 °C for a single day, or below −20 °C for 2 weeks) are not met for 40 years, and host pine species comprise more than 30 % of the simulation landscape. Beetle-caused tree mortality is simulated at the tree level using empirically-based regression equations that relate probability of mortality to tree diameter. Disturbance processes dynamically and reciprocally interact in the model when and where fires, beetle outbreaks, and/or rust infections alter composition and age structure of host trees and fuelbed properties, such as when beetle killed trees contribute fine fuels to the forest floor, facilitating spreading fires that may kill potential host trees and restrict future outbreak intensity. FireBGCv2 is a cumulative effects model best suited for evaluating long-term ecological dynamics such as those that develop under several disturbance cycles.
Study areas
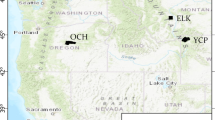
Study areas (Fig. 2) were selected because they represent a range of climate, vegetation, and fire regime types common to the Northern Rockies: (1) crown of the continent (CCE), a 93,000 ha high-elevation mixed conifer ecosystem in north-central Montana with an historical mixed-severity fire regime, (2) East Fork of the Bitterroot River (EFBR), a 128,000 ha, mid-elevation dry mixed-conifer ecosystem in western Montana with an historical elevation-dependent, low- to high-severity fire regime; and (3) Yellowstone Central Plateau (YCP), an 80,000 ha, high-elevation lodgepole pine ecosystem in western Wyoming with an historical low-frequency, high-severity fire regime. All three landscapes contain varying proportions of tree species that are fire-adapted, susceptible to white pine blister rust infection, and are mountain pine beetle hosts. Forest productivity, represented by tree basal area (ba, m2 ha−1, defined as the average cross-sectional area occupied by tree stem at 1.37 m above the ground), is highest for the CCE, followed by EFBR and YCP (Table 1). Model initialization, parameterization, calibration, and validation were performed as part of previous studies (Clark et al. 2016; Loehman et al. 2011a; Holsinger et al. 2014) (Supplementary material 1).

Simulation landscapes: 1 Crown of the Continent (CCE), 2 East Fork of the Bitterroot River (EFBR), and 3 Yellowstone Central Plateau (YCP)
Simulation design and analyses
We used a multifactorial design to evaluate the effects of climate and disturbance on forest productivity and tree mortality. We simulated all combinations of fire, beetles, and rust (“scenarios”) over a 300-year period for two climate factors. Current climate was derived from instrumental weather data from National Climatic Data Center (NCDC 2010-2013) cooperative stations within or near each of the study areas. Daily weather for 62 (CCE), 55 (EFBR), or 99 (YCP) years was used in sequence and repeated for multiple cycles until the 300-year simulation period was complete. Future climate was based on the RCP8.5 greenhouse gas concentration pathway, a baseline scenario that assumes no climate mitigation target and results in a radiative forcing of 8.5 W/m2 at the end of the century (Riahi et al. 2011). Data were produced using the Multivariate Adaptive Constructed Analogs (MACA) 1/24° (~4 km) statistical downscaling method applied to the CNRM-CM5 (National Centre of Meteorological Research France) global change model, part of the Coupled Model Inter-Comparison Project 5 (CMIP5) (Abatzoglou and Brown 2012; Rupp et al. 2013). Ten replicates were produced for each scenario to account for stochastic model processes such as cone crop abundance, tree mortality, and wildfire origination. We evaluated effects of climate and single or interacting disturbances on basal area (m2 ha−1) using analysis of variance followed by Duncan’s multiple comparisons tests of treatment effects. Analyses were performed using the R program (laercio package) (R Core Team 2015), and we report results at the significance level of 0.05. Although FireBGCv2 outputs many other variables basal area was selected because it is a widely used metric in forest ecology and management, is tightly coupled to leaf area and the basic ecophysiological processes of photosynthesis and respiration, and is useful for tracking disturbance-caused mortality (McDowell et al. 2002). Higher basal area values are associated with both more forested area and larger-stemmed (mature) trees. Basal area was produced annually for every forested stand (comprised of 90-m pixels) in the study areas but output at 10-year intervals for computational efficiency. We then averaged across intervals and replicates to produce a single representative basal area value for each scenario. We calculated the percent area (sum of individual stand areas divided by total landscape area) occupied by white pines (white pine blister rust host trees) and all pines (mountain pine beetle host trees) for each simulation and averaged across all replicates per scenarios to assess compositional shifts associated with disturbances.
Results
Disturbance effects
Basal area was highest for scenarios where no disturbances operated (CCE: 50.81 ± 3.44 m2 ha−1, EFBR: 33.19 ± 1.56 m2 ha−1, YCP: 37.49 ± 4.07 m2 ha−1) and lowest (CCE: 43.19 ± 3.95 m2 ha−1, EFBR: 27.02 ± 2.81 m2 ha−1) or nearly lowest (YCP: 30.15 ± 9.70 m2 ha−1) when all three disturbance processes—fire, white pine blister rust, and mountain pine beetles—co-occurred in scenarios (Table 1; Fig. 3). Fire, with or without other disturbances present, reduced basal area (by about eight to 30 % relative to no-disturbance scenarios) more than rust and/or beetle disturbance scenarios (less than one to about 9 %), regardless of simulated climate or study area. In all cases fire-caused reduction in basal area was significantly different than no-disturbance scenarios (p < 0.05), whereas in scenarios with only beetles and/or rust basal area was not always significantly different than scenarios without disturbance (Table 1). When present, fire also increased the variability in basal area across simulation replicates (Fig. 3) due to the temporal and spatial stochasticity of fire occurrence, a realistic outcome.

Averaged total basal area (m2 ha−1) across disturbance scenarios for three simulation landscapes and two climates. Scenarios are factorial combinations of disturbances: wildland fire (Fire), white pine blister rust (WPBR), and mountain pine beetle (MPB). Tails in boxplots indicate a skewed distribution
When fires were the only operating disturbance the percent of study areas occupied by pines increased relative to no-disturbance scenarios for both current and future climates. For example, without fire on the landscape white pines and all pines were present across ~21 and 27 % of the CCE, whereas when fires were simulated areal extent increased to ~28 and 32 % of the landscape (Table 1). When fire co-occurred with rust or beetles pines still occupied more landscape area than when only rust and/or beetle disturbance was simulated, although the co-occurrence of all three disturbances reduced areal extent of pines.
Climate impacts
Simulated future climate affected landscape basal area directly—even when no disturbances operated—and indirectly, as mediated by single or co-occurring, multiple disturbances. For all study areas and disturbance scenarios basal area was lower under future (RCP8.5) climate than for current climate (Table 1). Future climate most strongly influenced productivity of the YCP landscape, both as a direct effect (46 % reduction in basal area without disturbance) and indirectly, as mediated by disturbance (39–53 % lower) relative to current climate. Direct and indirect effects of future climate on basal area were less pronounced for the CCE (6–12 % lower than current climate) and EFBR (10–18 % lower) landscapes.
Landscape differences
Disturbance-caused changes in basal area were proportionate to the dominance of fire sensitive or host species on each landscape. For example, there are more white pines in the CCE (~21 % as compared with ~2 and ~4 % for EFBR and YCP), but a relatively smaller forest component susceptible to mountain pine beetle (Table 1), maximizing potential for blister rust-associated tree mortality. In contrast, the YCP is dominated by pine species that are both fire sensitive and susceptible to beetle attack and, as observed in the past, can be heavily modified by disturbance interactions (Lynch et al. 2006).
Discussion and conclusions
Disturbances and interactions alter landscapes
Disturbance interactions caused detectable, direct, and persistent changes to landscape basal area. Without doubt fire is the most pervasive and influential disturbance on the simulated landscapes; however, the interactions of fire with other disturbances reduced basal area more than when fire operated alone. Disturbance interactions were not purely additive, indicating the presence of non-linear behaviors and feedbacks; and, while forest disturbances may act independently or synchronously in space and time, their interactions are multidirectional. For example, in YCP, where most trees can be killed by both fire and beetles, stand-replacing fires likely reduced beetle mortality by limiting the abundance and distribution of host trees of sufficient threshold diameter, as has been noted elsewhere (Kulakowski et al. 2012). As simulated, the YCP landscape was more vulnerable to disturbances—especially fire and beetles—than CCE or EFBR, although all three landscapes would likely appear and function much differently should fire, beetle outbreaks, and rust infections co-occur over multiple disturbance cycles.
Although changes in basal area provide a measure of disturbance-related mortality and altered site productivity, the reduction in basal area that occurred with disturbance may reflect a return to pre-suppression era landscape patterns rather than a true shift away from ecological reference conditions (Loehman et al. 2011a). Western dry forests are currently in a state of “fire deficit” (Marlon et al. 2012), and fire exclusion from forested biomes has increased surface and canopy fuel loads and canopy cover, shifted the composition of mature forests toward late seral, shade-tolerant, non-fire adapted species, and hindered regeneration of fire-adapted species such as Ponderosa pine (Agee 1998; Taylor and Skinner 2003; Peterson et al. 2005). Simulations with fire increased the proportion of all pines on the simulation landscapes by removing late seral competitors, allowing for fire adapted species to re-establish in burned areas. Given expected climate change-driven increases in fire frequency and area burned in the coming decades (McKenzie et al. 2004; Flannigan et al. 2006), restoration of fire may reduce susceptibility (enhance resilience) of fire-prone and fire-adapted landscapes (Fule 2008) and maintain forested communities.
Climate changes cause direct impacts and act synergistically with disturbances
Climate changes decreased forest productivity and forested area, and further changes in basal area with co-occurring disturbances suggest that the synergy of climate changes and disturbance processes may amplify overall forest losses. Direct climate impacts (about eight to 46 % relative decline in basal area across landscapes), indicate that the study areas may experience a loss of area of climate suitability for some tree species in the future (Hansen and Phillips 2015). Projected climate changes potentiated disturbance activity and associated tree mortality: warmer temperatures increased annual burned area, individual fire size, and fire frequency (Clark et al. 2016; Loehman et al. 2011b; Holsinger et al. 2014) and relaxed low-temperature constraints on beetle and blister rust activity (Keane et al. 2015a), although the persistence of forest cover across several disturbance cycles indicates that landscapes are resilient—although not resistant—to climate-disturbance synergies. Importantly, fire maintained or increased the relative proportion of pines on the study landscape even in future climate, because species adaptations enabled survival and recruitment in a free-fire environment, in contrast to late-successional species.
Landscapes exhibit differential responses to climate changes and disturbance interactions
Variability in landscape response (sensitivity) to climate factors and disturbances reflects compositional differences and physiognomic characteristics (Keane et al. 2015a, b). Species composition and configuration influence fire behavior and fire effects, and host availability for and susceptibility to mountain pine beetle attacks and blister rust infections. Landscapes with a greater proportion of vulnerable species and/or greater connectivity (few barriers to spread of wildfire, insects, or pathogens) (McKenzie and Kennedy 2011) are less resistant to climate changes and disturbance interactions, and may demonstrate loss of ecological resilience; i.e., inability to recover from disturbances (Holling 1973). Our results suggest that YCP, and to a lesser degree EFBR and CCE, may be reorganized by climate-disturbance interactions. Evaluating landscape-specific disturbance sensitivity and response is critical for land management, as neglecting interaction effects can lead to an underestimation of climate-disturbance synergies (Seidl and Rammer 2016).
Limitations of simulation models
Modeling necessitates simplification of real-world processes, and the FireBGCv2 model simplifies or excludes some biophysical processes that affect disturbances. Tree defenses against mountain pine beetles were not included—for example, recent work by Hood et al. (2015) demonstrates that non-lethal, low-severity wildfire can increase resin duct production and provide improved physical defense against attack over fire-excluded trees—nor were other insects and pathogens found in the study landscapes (Loehman et al. 2016). Model results are influenced by stochastic elements of fire spread and tree regeneration (Keane et al. 2011), as well as parameter settings and simulation design. Although burdened by the axiom “all are wrong but some are useful (Sterman 2002),” models are important (and perhaps the only) tools for simulating emergent ecological behaviors, identifying vulnerable landscapes, and understanding complex ecological interactions across large areas and over long time periods. Accounting for disturbance interactions will be especially critical in the coming decades, as projections of warmer temperatures and increased drought stress suggest that the total area susceptible to or affected by disturbances—and in particular by beetle outbreaks and large or severe fires—may increase rapidly (Williams et al. 2012).
References
Abatzoglou JT, Brown TJ (2012) A comparison of statistical downscaling methods suited for wildfire applications. Int J Climatol 32(5):772–780
Agee J (1993) Fire ecology of Pacific Northwest forests. Island Press, Washington, DC
Agee JK (1998) The landscape ecology of western forest fire regimes. Northwest Sci 72:24–34
Albini, Frank A. Estimating wildfire behavior and effects (1976) General Technical Report INT-30. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, p 92
Allen CD (2007) Interactions across spatial scales among forest dieback, fire, and erosion in northern New Mexico landscapes. Ecosystems 10(5):797–808
Bachelet D, Lenihan JM, Daly C, Neilson RP (2000) Interactions between fire, grazing and climate change at Wind Cave National Park SD. Ecol Model 134(2):229–244
Beh MM, Metz MR, Seybold SJ, Rizzo DM (2014) The novel interaction between Phytophthora ramorum and wildfire elicits elevated ambrosia beetle landing rates on tanoak, Notholithocarpus densiflorus. For Ecol Manag 318:21–33
Bentz B, Régnière J, Fettig C, Hansen E, Hayes J, Hicke J, Kelsey R, Negrón J, Seybold S (2010) Climate change and bark beetles of the western United States and Canada: direct and indirect effects. BioScience 60(8):602–613
Brown CD, Johnstone JF (2012) Once burned, twice shy: repeat fires reduce seed availability and alter substrate constraints on Picea mariana regeneration. For Ecol Manag 266:34–41
Brown TJ, Hall BL, Westerling AL (2004) The impact of twenty-first century climate change on wildland fire danger in the western United States: an applications perspective. Clim Chang 62(1):365–388
Buma B (2015) Disturbance interactions: characterization, prediction, and the potential for cascading effects. Ecosphere 6(4):1–15
Buma B, Wessman C (2012) Differential species responses to compounded perturbations and implications for landscape heterogeneity and resilience. For Ecol Manag 266:25–33
Campbell EM, Antos JA (2000) Distribution and severity of white pine blister rust and mountain pine beetle on whitebark pine in British Columbia. Can J For Res 30(7):1051–1059
Coops NC, Wulder MA, Waring RH (2012) Modeling lodgepole and jack pine vulnerability to mountain pine beetle expansion into the western Canadian boreal forest. For Ecol Manag 274:161–171
Clark JA, Loehman RA, Keane RE (2016) Climate changes and wildfire alter vegetation of Yellowstone National Park, but forest cover persists. In Review, Ecosphere
Edburg SL, Hicke JA, Brooks PD et al (2012) Cascading impacts of bark beetle-caused tree mortality on coupled biogeophysical and biogeochemical processes. Front Ecol Environ 10(8):416–424
Fettig CJ, Klepzig KD, Billings RF, Munson AS, Nebeker TE, Negrón JF, Nowak JT (2007) The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For Ecol Manag 238(1):24–53
Flannigan MD, Amiro BD, Logan KA, Stocks BJ, Wotton BM (2006) Forest fires and climate change in the 21st century. Mitig Adapt Strateg Glob 11(4):847–859
Fule PZ (2008) Does it make sense to restore wildland fire in changing climate? Restor Ecol 16(4):526–531
Gibson K, Kegley S, Bentz B (2009) Mountain pine beetle, vol 2. USDA Forest Service, Forest Insect and Disease Leaflet, Portland
Goetz S, Mack M, Gurney K, Randerson J, Houghton R (2007) Ecosystem responses to recent climate change and fire disturbance at northern high latitudes: observations and model results contrasting northern Eurasia and North America. Environ Res Lett 2(4):045031
Grime JP (1973) Competitive exclusion in herbaceous vegetation. Nature, UK 242(5396):344–347
Gross JE, Tercek M, Guay K, Talbert M, Chang T, Rodman A, Thoma D, Jantz P, Morisette JT (2016) Analyses of historical and projected climates to support climate adaptation in the Northern Rocky mountains. Climate change in wildlands: pioneering approaches to science and management, p 55
Hansen AJ, Phillips LB (2015) Which tree species and biome types are most vulnerable to climate change in the US Northern Rocky Mountains? For Ecol Manag 338:68–83
Hansen A, Ireland K, Legg K, Keane R, Barge E, Jenkins M, Pillet M (2016) Complex challenges of maintaining whitebark pine in greater Yellowstone under climate change: a call for innovative research, management, and policy approaches. Forests 7(3):54
Heyerdahl EK, Morgan P, Riser JP (2008) Multi-season climate synchronized historical fires in dry forests (1650–1900), northern Rockies USA. Ecology 89(3):705–716
Hicke JA, Johnson MC, Hayes JL, Preisler HK (2012) Effects of bark beetle-caused tree mortality on wildfire. For Ecol Manag 271:81–90
Hobbs RJ, Abbott I, Burrows N (2003) How fire regimes interact with other forms of ecosystem disturbance and modification. In: Fire in ecosystems of south-west Western Australia: impacts and management. Symposium proceedings (vol I), 16–18 April 2002, Backhuys Publishers, Perth, pp 421–436
Hoff R, Hagle S (1990) Diseases of whitebark pine with special emphasis on white pine blister rust. In: Schmidt WC (ed) Symposium on whitebark pine ecosystems: ecology and management of a high-mountain resource. USDA Forest Service, Intermountain Research Station, Ogden, pp. 335–340
Holling CS (1973) Resilience and stability of ecological systems. Annu Rev Ecol Syst 4:1–23
Holsinger L, Keane RE, Isaak DJ, Eby L, Young MK (2014) Relative effects of climate change and wildfires on stream temperatures: a simulation modeling approach in a Rocky Mountain watershed. Clim Chang 124(1–2):191–206
Hood S, Sala A, Heyerdahl EK, Boutin M (2015) Low-severity fire increases tree defense against bark beetle attacks. Ecology 96(7):1846–1855
Johnstone JF, Chapin FS, Hollingsworth TN, Mack MC, Romanovsky V, Turetsky M (2010a) Fire, climate change, and forest resilience in interior Alaska. Can J For Res 40(7):1302–1312
Johnstone JF, Hollingsworth TN, Chapin FS, Mack MC (2010b) Changes in fire regime break the legacy lock on successional trajectories in Alaskan boreal forest. Glob Chang Biol 16(4):1281–1295
Keane R, Morgan P (1994) Landscape processes affecting the decline of whitebark pine (Pinus albicaulis) in the Bob Marshall Wilderness Complex. In: Proceedings of the 12th international conference on fire and forest meteorology, pp 26–28
Keane R, Tomback D, Aubry C, Bower A, Campbell E, Cripps C, Jenkins M, Mahalovich M, Manning M, McKinney S (2012) A range-wide restoration strategy for whitebark pine (Pinus albicaulis). Gen. Tech. Rep. RMRS-GTR-279. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, p 108
Keane RE, Loehman R, Clark J, Smithwick EA, Miller C (2015a) Exploring interactions among multiple disturbance agents in forest landscapes: simulating effects of fire, beetles, and disease under climate change. In: Perera AH, Remmel TK, Buse LJ (eds) Simulation modeling of forest landscape disturbances. Springer International Publishing, Switzerland, pp 201–231
Keane RE, Loehman RA, Holsinger LM (2011) The FireBGCv2 landscape fire and succession model: a research simulation platform for exploring fire and vegetation dynamics. Gen. Tech. Rep. RMRS-GTR-255, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins
Keane RE, McKenzie D, Falk DA, Smithwick EA, Miller C, Kellogg L-KB (2015b) Representing climate, disturbance, and vegetation interactions in landscape models. Ecol Model 309:33–47
Koerner SE, Collins SL (2014) Interactive effects of grazing, drought, and fire on grassland plant communities in North America and South Africa. Ecology 95(1):98–109
Kulakowski D, Jarvis D, Veblen TT, Smith J (2012) Stand-replacing fires reduce susceptibility of lodgepole pine to mountain pine beetle outbreaks in Colorado. J Biogeog 39:2052–2060
Kurz WA, Dymond CC, Stinson G, Rampley J, Neilson ET, Carroll AL, Ebata T, Safranyik L (2008a) Mountain pine beetle and forest carbon feedback to climate change. Nature 452(7190):987–990
Kurz WA, Stinson G, Rampley GJ, Dymond CC, Neilson ET (2008b) Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proc Natl Acad Sci USA 105:1551–1555
Larson ER (2011) Influences of the biophysical environment on blister rust and mountain pine beetle, and their interactions, in whitebark pine forests. J Biogeogr 38(3):453–470
Leirfallom SB, Keane RE, Tomback DF, Dobrowski SZ (2015) The effects of seed source health on whitebark pine (Pinus albicaulis) regeneration density after wildfire. Can J For Res 45(11):1597–1606
Lewis KJ, Lindgren BS (2002) Relationship between spruce beetle and tomentosus root disease: two natural disturbance agents of spruce. Can J For Res 32(1):31–37
Loehman R, Bentz B, Keane RE (2016) Effects of Climate Change on Ecological Disturbance. In: Halofsky JE, Peterson DL, Dante-Wood SK, Hoang L (eds) Climate change vulnerability and adaptation in the Northern Rocky Mountains. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ft. Collins
Loehman RA, Clark JA, Keane RE (2011a) Modeling Effects of Climate Change and Fire Management on Western White Pine (Pinus monticola) in the Northern Rocky Mountains USA. Forests 2(4):832–860
Loehman RA, Corrow A, Keane RE (2011b) Modeling climate change and disturbance interactions: effects on whitebark pine (Pinus albicaulis) and implications for restoration, Glacier National Park. In: Keane RE, Tomback DF, Murray MP, Smith C (eds) The future of high-elevation, five-needle white pines in Western North America: proceedings of the high five symposium, Missoula, MT, 28–30 June 2010, Proceedings RMRS-P-63. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, pp 176–189
Loehman RA, Reinhardt E, Riley KL (2014) Wildland fire emissions, carbon, and climate: seeing the forest and the trees—A cross-scale assessment of wildfire and carbon dynamics in fire-prone, forested ecosystems. For Ecol Manag 317:9–19
Lynch HJ, Renkin RA, Crabtree RL, Moorcroft PR (2006) The influence of previous mountain pine beetle (Dendroctonus ponderosae) activity on the 1988 Yellowstone fires. Ecosystems 9(8):1318–1327
Mahalovich MF, Burr KE, Foushee DL (2006) Whitebark pine germination, rust resistance, and cold hardiness among seed sources in the Inland Northwest: planting strategies for restoration. In: Riley LE, Dumroese RK, Landis TD (eds) National Proceedings: forest and conservation nursery associations—2005. Proceedings of RMRS-P-43. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, pp 91–101
Marlon JR, Bartlein PJ, Gavin DG, Long CJ, Anderson RS, Briles CE, Brown KJ, Colombaroli D, Hallett DJ, Power MJ (2012) Long-term perspective on wildfires in the western USA. Proc Natl Acad Sci USA 109(9):E535–E543
Matson E, Bart D (2013) Interactions among fire legacies, grazing and topography predict shrub encroachment in post-agricultural páramo. Landscape Ecol 28(9):1829–1840
McDonald GI, Hoff RJ, Wykoff W (1981) Computer simulation of white pine blister rust epidemics: Model formulation. Res. Pap. INT-RP-258. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden
McDowell N, Barnard H, Bond B, Hinckley T, Hubbard R, Ishii H, Köstner B, Magnani F, Marshall J, Meinzer F (2002) The relationship between tree height and leaf area: sapwood area ratio. Oecologia 132(1):12–20
McKenzie D, Kennedy MC (2011) Scaling laws and complexity in fire regimes. In: McKenzie D, Miller C, Falk DA (eds) The landscape ecology of fire. Springer, New York, pp 27–49
McKenzie D, Ze’ev G, Peterson D, Mote P (2004) Climatic change, wildfire, and conservation. Conserv Biol 18(4):890–902
Miller RF, Wigand PE (1994) Holocene changes in semiarid pinyon-juniper woodlands. Bioscience 44(7):465–474
Mladenoff DJ (2004) LANDIS and forest landscape models. Ecol Model 180(1):7–19
Parker TJ, Clancy KM, Mathiasen RL (2006) Interactions among fire, insects and pathogens in coniferous forests of the interior western United States and Canada. Agric For Entomol 8(3):167–189
Peterson DL, Johnson MC, Agee JK, Jain TB, McKenzie D, Reinhardt ED (2005) Forest structure and fire hazard in dry forests of the western United States. Gen. Tech. Rep. PNW-GTR-628. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Portland
Riahi K, Rao S, Krey V, Cho C, Chirkov V, Fischer G, Kindermann G, Nakicenovic N, Rafaj P (2011) RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Clim Chang 109(1–2):33–57
R Development Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Riggs RA, Keane RE, Cimon N, Cook R, Holsinger L, Cook J, DelCurto T, Baggett LS, Justic D, Powell D (2015) Biomass and fire dynamics in a temperate forest-grassland mosaic: integrating multi-species herbivory, climate, and fire with the FireBGCv2/GrazeBGC system. Ecol Model 296:57–78
Romme WH, Despain DG (1989) Historical perspective on the Yellowstone fires of 1988. Bioscience 39(10):695–699
Rupp DE, Abatzoglou JT, Hegewisch KC, Mote PW (2013) Evaluation of CMIP5 20th century climate simulations for the Pacific Northwest USA. J Geophys Res Atmos 118(19):10884–10906
Safranyik L, Carroll A, Régnière J, Langor D, Riel W, Shore T, Peter B, Cooke B, Nealis V, Taylor S (2010) Potential for range expansion of mountain pine beetle into the boreal forest of North America. Can Entomol 142(5):415–442
Schoennagel T, Veblen TT, Romme WH (2004) The interaction of fire, fuels, and climate across Rocky Mountain forests. BioScience 54(7):661–676
Schoettle AW, Sniezko RA (2007) Proactive intervention to sustain high-elevation pine ecosystems threatened by white pine blister rust. J For Res 12(5):327–336
Seidl R, Rammer W (2016) Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landscape Ecol. doi:10.1007/s10980-016-0396-4
Simard S (1991) Fire severity, changing scales, and how things hang together. Int J Wildland Fire 1(1):23–34
Sterman JD (2002) All models are wrong: reflections on becoming a systems scientist. Sys Dyn Rev 18(4):501–531
Taylor AH, Skinner CN (2003) Spatial patterns and controls on historical fire regimes and forest structure in the Klamath Mountains. Ecol Appl 13(3):704–719
Temperli C, Bugmann H, Elkin C (2013) Cross-scale interactions among bark beetles, climate change, and wind disturbances: a landscape modeling approach. Ecol Monogr 83(3):383–402
Tomback DF, Achuff P (2010) Blister rust and western forest biodiversity: ecology, values and outlook for white pines. For Pathol 40(3–4):186–225
Turner MG (2005) Landscape ecology: what is the state of the science? Annu Rev Ecol Evol Syst 36:319–344
Turner MG (2010) Disturbance and landscape dynamics in a changing world 1. Ecology 91(10):2833–2849
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western U.S. forest wildfire activity. Science 313:940–943
Williams AP, Allen CD, Macalady AK, Griffin D, Woodhouse CA, Meko DM, Swetnam TW, Rauscher SA, Seager R, Grissino-Mayer HD (2012) Temperature as a potent driver of regional forest drought stress and tree mortality. Nat Clim Chang 3:292–297
Acknowledgments
This research was funded in part by the Joint Fire Science Program under projects JFSP 09-3-01-17 and JFSP-09-1002-9. The authors gratefully acknowledge the contributions of the large number of skilled researchers, ecologists, and biological technicians who have participated in many phases of this research. Any use of trade names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Loehman, R.A., Keane, R.E., Holsinger, L.M. et al. Interactions of landscape disturbances and climate change dictate ecological pattern and process: spatial modeling of wildfire, insect, and disease dynamics under future climates. Landscape Ecol 32, 1447–1459 (2017). https://doi.org/10.1007/s10980-016-0414-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0414-6