
Abstract
Forest productivity is driven by a suite of direct climatic and non-climatic factors that are transient or permanent. The kind of productivity driver and the nature of their effects vary by species, and scale dependencies potentially complicate these relationships. This study explored productivity-driver relations in eastern Boreal Canada and determined spatial effects in productivity control when expressed with stand dominant height at a reference age (site index). Data from 4,217 temporary sample plots obtained from boreal mixedwood and conifer bioclimatic domains, and with varied species composition, were used in this study. A single-level global model that assumes equal sensitivities across spatial scales was calibrated and compared with three alternative models reflecting different hypotheses on possible spatial heterogeneities. Alternative models were calibrated by plot-level soil deposit types (microscale), landscape dominant deposits (mesoscale) and bioclimatic domains (macroscale). A marked difference between the global and alternative models was observed, suggesting that a single global model does not sufficiently reflect existing heterogeneity in productivity-driver relationships. A combination of macro- and microscale models provided the best explanation of site index. Results further showed that site index is mainly driven by species composition (complementarity effects of aspen and jack pine compositions) and stand diameter structural diversity effects. It is concluded that successional changes, more than direct climatic effects, drive productivity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forest productivity, defined as the potential of a particular forest stand to produce above-ground biomass, is driven by a suite of direct climatic and non-climatic factors (Landsberg and Sands 2011). These may include effects from drought, frost, insect damage, nutrient deposition, etc. In addition, indirect effects complicate productivity-environmental factor relationships. For example, productivity in boreal stands is high directly following a severe fire and decreases with time and with the accumulation of organic material on the soil surface (e.g. Simard et al. 2007). Structural and compositional changes, and stand ageing can induce feedback effects on productivity through changes in litter decomposition and canopy species displacements (Prescott et al. 2000). Both the nature and rate of these dynamics differ by soil deposit types (Belleau et al. 2011). The kind of productivity drivers and the nature of their effects can vary by species partly as a result of differences in the topographic gradients they occupy (e.g. Vallet and Pérot 2011).
Productivity drivers could be transient (e.g. Wang et al. 2013) or permanent (Laliberté et al. 2013), and in the latter case they provide a geographical basis for ecological classification. There is consensus on the fact that fine mixes of transient and permanent site factors explain geographical patterns of species distribution and determine range limits (e.g. Tang et al. 2012) and, while equilibrium between the two is possible, it is rather rare and an exception in nature. Non-equilibrium between these factors (transient vs. permanent) justifies the need for niche models that attempt to explain the differential contributions of these factors in studies on species distribution and abundance and on range limits (e.g. Tang et al. 2012; Iversion and Mackenzie 2013) .
Scale-dependencies and temporal variability of drivers of forest productivity are increasingly being reported in the literature. Spatial variability in species-specific growth sensitivities to climate factors has been attributed to the existence of ample genetic variations among populations (O’Neill et al. 2008). Heterogeneity in growth-climate associations across latitudinal and longitudinal gradients may take effect from the various sources of air masses (e.g., maritime versus polar and dry versus moist) across a given species’ range. Because productivity drivers are scale-dependent, the scale (i.e. site vs. landscape versus regional levels) at which productivity models are calibrated may account for significant differences in the inclusion or not of explanatory variables for forest productivity predictions (Aersten et al. 2011). The application of models on extents which do not overlap with the extent at which they were developed (either through extrapolation in time or in space) should result in inadequate predictions.
Site index (SI), defined as the mean dominant height at a reference stand age, is the most widely used site productivity evaluation method in North America (Monserud 1984). Application of existing SI models have contributed to the understanding of productivity dynamics (e.g. McKenney and Pedlar 2003; Pinto et al. 2008; Aersten et al. 2011; Coops et al. 2011 for a review). These models, however, remain species-specific and in most cases rely on permanent productivity drivers. Additionally, SI models are difficult to apply in mixed stands because height and age estimates of dominant trees must be available for each species in a stand and these are generally not available due to the expense of collecting such data (Weiskittel et al. 2011). Also, because of mortality or displacement of the dominant cohort through time (Mailly et al. 2009), capturing the dynamics of the actual site potential by species is almost impossible. An estimate of site potential that is independent of species could potentially address this limitation (Curtis and Post 1964).
The objectives of this study were to determine (1) to what extent transient and permanent site factors influence site productivity and, (2) spatial differences in SI control in eastern boreal dense coniferous and mixedwood landscapes. Four hypotheses were verified in this regard. First, both the nature and rates of stand structural and species compositional changes differ by stand-level soil deposit types. Also, there could be accumulation of organic material through time driven by local topography and climate, it is therefore hypothesized that productivity is influenced by local transient and permanent site effects to different extents (H1—microscale hypothesis). Because disturbance events create mosaics driven by topography, regional relief, regional climate etc., and these occur at scales larger than plot-level, it is also postulated that stands of close proximity/latitude, and therefore of similar regional climate or with same parent material or of same disturbance regime, do not significantly differ in their productivity dynamics (H2—mesoscale hypothesis). At the landscape-level, conifer landscapes tend to be of less species richness compared to landscapes dominated by mixedwood (NFI 2009) largely due to differences in climate which thereby has consequences for productivity (Vila et al. 2013). It is therefore also expected that there will be differences in productivity controls by bioclimatic domain (H3—macroscale hypothesis). Finally, in view of the suspected spatial and temporal variability, a single global model should be less adequate when compared to a combination of models calibrated along various heterogeneous gradients (H4—mix of scales hypothesis).
Materials and methods
The hypotheses were verified using data from a network of temporary sample data in Quebec, Canada. SI was estimated with the dominant height model of Pothier and Auger (2009). An approach where the global mean of SI from the calibration data is modified by explanatory variables was applied in model building. We verified plot-level-substrate SI-driver relationships (micro, H1) and regional level correlations i.e. dominant deposits by ecological districts (meso-scale, H2) and then bioclimatic scale relationships (macro-scale, H3). The multi-scale approach enabled us to verify the drivers peculiar to a scale and to understand the nature of cross-scale productivity drivers. Finally, we compared a model calibrated using the entire dataset (global model), with a combination of models calibrated by plot-level substrates, regional substrates or bioclimatic domains (H4).
Study area
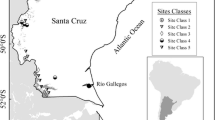
The study site is a 3 × 3 degree wide area situated in western Québec that spans from the southern mixedwood to the northern conifer dominated limit of commercial forests. Coarse deposits dominate the southern section of the study region as well as the northeastern areas, while the northwest is predominantly clay. Mor is the most frequent humus type. The region is characterized by a generally flat topography especially in the western and northern areas and is slightly hilly (≈400 m a.s.l.) and undulating (≈11 % slope) in the southwest. Drainage is generally moderate to poor. The study area covers balsam fir (Abies balsamea (L.) Mill.)—white birch (Betula papyrifera Marsh.) and black spruce (Picea mariana (Mill.) B.S.P.)—feather moss bioclimatic domains (Robitaille and Saucier 1998), which are also respectively mixedwood and conifer regions (Fig. 1). The mixedwood region lies in the southern zone of Québec’s boreal forest where pest outbreak (spruce budworm defoliation) is the main disturbance event (Jasinski and Payette 2005). However, fire also plays an important role in forest dynamics especially in the western part of the domain where fire cycles are relatively shorter when compared to the eastern parts due to higher precipitation amounts in the latter (Jasinski and Payette 2005). Annual mean temperature for the region ranges from −1.7 to 2.2 °C and it receives a mean annual total rainfall of 883 mm year−1 (sd. 47 mm year−1).

Study area map; dots indicate study plots, the shaded region is the mixedwood and the unshaded region is the conifer zone. Grey lines indicate ecological districts and the red line is the limit of commercial forest. (Color figure online)
Databases
The study data comprises 4,217 temporary sample plots selected from the third inventory program conducted by Quebec’s Ministry of Natural Resources (MRN). In 1970, the MRN started a 10-year province-wide measurement program, where circular plots of size 0.04 ha were positioned in forested lands through a stratified sampling scheme. The aim was to adequately sample the most common forest types in a given sampling unit and this was done by aggregating most similar polygons into forest strata. In every plot, all trees >9 cm DBH (diameter at breast height, 1.3 m) were measured by species, diameter, height, and age at 1 m (from ring counting) were recorded for one to five study trees (usually three individuals). During its third inventory program (1992–2002), measurements were extended to include soil features which previously were only part of permanent sample plot inventories (Rouleau 1994).
Forest fire data was also obtained from the Ministry of Natural Resources and Wildlife of Québec (MRNF). The database contains information on the location, date of detection, size (ha), and cause (lightning or human) of all fires recorded in the province of Quebec. We considered all lightning and human-caused fires from 1924 to 2009.
Potential drivers of productivity
Soil features
Table S1 provides the list of site variables measured during the inventory process and considered in this study. Drainage classes included excessive (0), rapid (1), good (2), moderate (3), poor (4), bad (5), very bad (6). There were six humus types; mull (MU), moder (MD), mor (MR), peat (TO), ‘Ammoor’ (AN) and organic (SO) (Rouleau 1994). Re-classification and grouping of some variables were done in order to validate assumptions of their influences on productivity. Soil deposit variables were regrouped first into coarse, till, clay and organic types given that successional pathways differ among these classes as reported by Belleau et al. (2011) and could have consequences for productivity. Secondly, Mansuy et al. (2010) suggested that surface deposit and drainage combinations lead to identifying areas of peculiar burn potentials. We verified if particular burn potentials were linked to productivity by regrouping 54 deposit types into 7 groups coded VAVC, MM, MAM, MAC, ROC, ORG and SAN. VAVC denotes moraine and juxta-glacial deposits that are characterized by a very high abundance of stones and boulders and a very coarse texture with extremely high potential for drying. MM refers to undifferentiated thick (>1 m) tills with moderate stoniness, while MAM are undifferentiated thin (0.25–1 m) tills with moderately abundant stoniness. MAC refers to outwash with moderately abundant stoniness (sandy to sandy silt texture) and xeric drainage. ROC are rock outcrops with excessive drainage but with a thin organic or till layer and ORG are organic deposits that characterize peatlands. SAN is a new category created for sand dune deposit types that were not in any of the established classes. Humus forms change with succession (Trap et al. 2011), as well as depth of organic layer and drainage classes; other soil features were considered permanent site variables.
Climatic variables
Overall, 38-climate variables (Table S1) obtained with the BioSIM model (ver. 8, Régnière and Saint-Amant 2008) for all study plots were considered in this study. Monthly water balance is the difference between monthly Thornwaite’s potential evapotranspiration and monthly precipitation and it is equated to zero if negative (Dunne and Leopold 1978). Studies (e.g. Vila et al. 2013) reported dominance of annual climate variables on growth relative to seasonal estimates, but there is also evidence of seasonal climatic influence on ecosystem processes (e.g. Wang et al. 2013). Therefore, we considered annual climate as well as spring (April, May, June) and summer (July, August, September) periods. BioSIM interpolates climate from a specified number of weather stations nearest to the point of interest and accounts for differences in elevation and aspect in relation to those of the weather stations. It also accounts for the distance of weather stations to the point of interest by assigning greater interpolation weights to nearer stations. The five nearest weather stations were used for interpolations of climatic variables (Anyomi et al. 2012). We obtained 50-year averages so that estimated climate values were within the same time frame as that of SI. Climate variables were considered transient variables in this study.
Indices of stand evolution
To the extent that stand composition and structure change through time, indices of stand evolution were considered as transient variables in this study. Stand structural and compositional changes are correlated with productivity (Boucher et al. 2006). Therefore, we estimated Shannon index, Shannon evenness index, Simpson index (Lexerød and Eid 2006) and basal area proportions as indices of stand structural and compositional changes. Basal area proportion that is spruce is hereafter simply referred to as spruce basal area proportion, and similarly for other species. Productivity is unstable through time either due to organic layer accumulation that leads to soil cooling and reduced mineralization (Simard et al. 2007) or effects of climate warming (Xu et al. 2013) or atmospheric nutrient deposition (Bontemps et al. 2009). It could imply an ageing constraint related to limiting soil nutrients as soils change (Laliberté et al. 2013). Consequently, stand age is an index of stand evolution; we used age at 1 m height in this study.
Site index modeling
Estimating site index
In order to estimate SI, we modified the stand-level growth model of Pothier and Auger (2009) to provide estimates of mean dominant height at a reference age of 50-years which is technically site index. The model was originally designed to estimate dominant height \(H_{d,2}\) (m) from an earlier estimate of dominant height \(H_{d,1}\) after a period of \(\Delta t\) years as illustrated in Eq. 1. The parameters \(\beta_{1}\), \(\beta_{2}\) depend on stand diversity (\(f_{sh}\)), asymptotic height (\(f_{{\bar{h}_{\hbox{max} } }}\)) and time lapse (\(f_{\Delta t}\)). The variables \(b_{1}\) and \(b_{2}\) are co-efficients, e is the base of natural logarithm and \(\varepsilon\) is model error. Dominant height estimated with this model is not species specific.
where \(\beta_{1} = b_{2} \,f_{{\bar{h}_{\hbox{max} } }} f_{\Delta t} f_{sh}\) and \(\beta_{2} = b_{1} \beta_{1}\) \(f_{{\bar{h}_{\hbox{max} } }} = 1 + c_{{\tilde{h}_{\hbox{max} } }} \left( {\frac{{\tilde{h}_{\hbox{max} } - \bar{\tilde{h}}_{\hbox{max} } }}{{\bar{\tilde{h}}_{\hbox{max} } }}} \right)\), where \(\tilde{h}_{\hbox{max} } = 1 + \left[ {\frac{{H_{d,1} - 1}}{{1 - e^{{ - b_{2} A_{1} }} }}} \right]^{{b_{2} b_{1} }}\) is asymptotic height, and \(\bar{\tilde{h}}_{\hbox{max} }\) is the mean of asymptotic height estimated across all plots but specific to ecological sub-region. \(f_{\Delta t} = 1 + c_{\Delta t} \left( {\frac{{\Delta t - \overline{\Delta t} }}{{\overline{\Delta t} }}} \right)\), where \(\overline{\Delta t}\) is mean time lapse between two measurements. \(f_{{S_{h} }} = 1 + c_{{S_{h} }} \left( {\frac{{S_{h} - \bar{S}_{h} }}{{\bar{S}_{h} }}} \right)\), where \(S_{h} = \frac{{ - \sum\nolimits_{i = 1}^{s} {p_{i} \ln (P_{i} )} }}{\ln (k)}\) is the Shannon index and \(\bar{S}_{h}\) is the mean Shannon index estimated across all plots but specific to each ecological sub-region. \(i\) is merchantable (>9 cm) diameter class, \(p_{i}\) is the proportion of basal area in diameter class \(i\). \(k\) corresponds to the 96th percentile of the number of diameter classes observed in a plot (13 classes) (Pothier and Auger 2009). \(\text{In}\) is the natural logarithm.
In obtaining SI, one could use the asymptotic height which is by definition the maximum height attainable at a site for a maximum stand age i.e. from Eq. 1, \(A_{1} + \Delta t\) = asymptote (Raulier et al. 2003). Alternatively, given that SI is mean of dominant height at a reference age of 50-years, mean height estimate \(H_{d,2}\) (m) obtained at stand age of 50 years could equally be applicable (i.e. \(A_{1} + \Delta t\) = 50; Ouzennou et al. 2008). In this study, we applied the latter approach in obtaining an index of productivity (S) at a reference age of 50 years thus, Eq. (2).
All parameters remain as described earlier.
Stand dominant height \(H_{d}\)(m), is the mean height of the four trees of largest diameters per 400 m2, and which corresponds to the 100 largest trees per hectare. By the inventory design, an insufficient number of top height trees was sampled per plot and as a consequence, stand dominant height was estimated using Eq. 2a below.
where \(H_{d}\) is predicted dominant height (m), \(\bar{D}_{4}\) is mean diameter (cm) of the four largest trees on a plot, \(\bar{D}\) and \(\bar{H}\) are respectively mean diameter and height of all merchantable trees for which height was measured, \(\lambda_{2}\) is a parameter specific to each species group (Pothier and Auger 2009).
Modeling approach
A variance analysis was first carried out in pre-selecting the variables that were significantly correlated (P < 0.05) with site index using a combination of mixed linear models and general-purpose regressions. The GLIMMIX procedure in SAS was used in verifying and accounting for spatial autocorrelation. As a way of introducing a pre-selected explanatory variable into the model and in determining its effects on the response variable, we modified the global mean value of SI observed in the calibration data with selected explanatory variables as illustrated in Eq. 3.
where \({\bar{S}}\) is global mean site index estimated from the data set. The global mean SI is modified by the product of n selected explanatory variables and each modifier has a value close to unity when the variable \(x_{i}\) equals to its global average \(\bar{x}_{i}\) value observed with the calibration data, and increases or decreases when moving further away from this average as illustrated in Eq. 3a below.
where \(\upbeta_{l.Xi}\) and \(\upbeta_{q.Xi}\) represent the linear and quadratic effects of the variables \(x_{i}\) on SI. Categorical explanatory variables (soil surface deposit types, texture etc.) were coded with a combination of binary variables so that they could be used in the NLMIXED procedure of SAS (\(\sum\nolimits_{{\text{j} = \text{1}}}^{\text{m}} {\upbeta_{\text{j}} \text{z}_{\text{j}} }\)) as follows (Eq. 4):
where \(\beta_{\text{j}}\) represents the effect of a categorical variable \(z_{\text{j}}\) on productivity and i and j represent the number of selected explanatory variables and deposit types respectively. As explanatory variables, we retained the variables that provided the highest correlation with SI, together with the lowest root mean square error (RMSE) and, subsequently, added other variables one by one. Inclusion of covariates in the model was in two stages: (a) we first studied model residuals to verify which variables were most significantly (P < 0.05) related with SI and (b) through a stepwise procedure with forward selection, we selected the variable combination that most explained the variability in SI. In avoiding model over-fitting, a covariate was only retained in the model if it was significant (P < 0.05) in the model and additionally contributed substantially (at least 1 %) to the variance explained (or in reducing RMSE). Entry or exit of a modifier was determined with a likelihood-ratio test. In order to capture random variation, we further introduced a random error (\(s\)) variable into Eq. (3) to account for homogeneity associated with stands sharing a common property, such as belonging to the same landscape (Anyomi et al. 2013), thereby obtaining Eq. (5).
The parameters of Eq. (5) were estimated with the NLMIXED procedure. Moran’s I test was conducted to verify model residuals for spatial autocorrelation (Dormaan et al. 2007) using the VARIOGRAM procedure in SAS.
Evaluation of hypotheses
Model calibration for each hypothesis
H1
Productivity is influenced by local (microscale) transient and permanent site effects to different extents
In verifying this hypothesis, variance explained individually by plot-level permanent and transient variables was compared using the MIXED procedure in SAS in combination with REG procedure. We then kept soil surface deposit type (permanent site feature) constant, given that successional dynamics differ by soil deposit (Belleau et al. 2011) and determined variance explained by temporal site factors (i.e. calibrated models for each deposit).
H2
Productivity is homogenous within a regional unit (mesoscale model)
In verifying hypothesis H2, we calibrated models by surface deposit type dominating within an ecological district. Ecological districts are portions of land characterized by a distinctive pattern of relief, geology, geomorphology and regional vegetation expressed at a scale of 1:250,000 (Bergeron et al. 1992) and were found to be appropriate units for characterizing regional-level productivity (Anyomi et al. 2013). For the disturbance regime, average burn rate per year was obtained at regional landscape units because districts were too small relative to fire events. The regional landscape units are defined on the basis of similarity of biophysical factors and vegetation type (i.e. portions of the territory with a recurrent arrangement of the main permanent biophysical and vegetation factors) (Robitaille and Saucier 1998).
H3
Productivity is driven differently by bioclimatic domains (macroscale model)
Here, we verified productivity-driver relations by bioclimatic domain thus, conifer and mixedwood landscapes.
H4
A single global model is inadequate (mix-of-scales hypothesis)
Here a model was first calibrated using all 4,217 plots hereafter called the global model and then rated against four alternative model sets using Akaike information criterion (AIC). Two types of alternative sets were considered; the first type consisted in combining all the models of one hypothesis (H1–H3) thus, three alternative sets and the second type of set consisted of finding the best combination of models from any hypothesis with an integer programming procedure. Predicted values of alternative models were chosen with binary variables in a nested manner, giving the selection priority first to the values predicted with models from the microscale, then mesoscale and finally macroscale hypotheses. Integer programming was used to estimate the set of binary variables (and consequently the combination of models) that provided the lowest mean square error for the whole plot dataset. No further constraints were considered.
Results
Globally, a mean SI of 16 m (std: 3.48 m) was observed for the entire study area, albeit spatial variability between ecological districts existed (AIC = 21,744, P < 0.0001, Fig. 2). Shannon index (r = 0.61, AIC = 20,788, P < 0.0001), stand age (r = −0.30, AIC = 22,343, P < 0.0001) and aspen (Populus tremuloides Michx.) basal area proportion (r = 0.58, AIC = 21,035, P < 0.0001) exhibited moderate associations with SI by ecological district (Fig. 2). No substantial differences in AIC were observed and coefficients of determination remained the same when relationships that accounted for spatial autocorrelation were compared with those that did not (Table S2). Also, we did not observe major differences in SI-annual climate correlations when compared with SI-seasonal climate correlations (Fig. S1). Moran’s I test shows that residuals of the global model did not exhibit significant spatial autocorrelation (Z = 0.699, P = 0.4848; Fig. S2). Calibrated models can be found in Table S3, and parameter values in Table S4 while model fits are illustrated in Fig. S3.

Heterogeneity in A site index and site index drivers; B Shannon index, C aspen basal area proportion and D stand age in years
H1
SI is influenced by local transient and permanent site effects to different extents
Total annual precipitation, basal area proportions of jack pine (Pinus banksiana Lamb) and thuya (Thuya occidentalis Linnaeus) were the transient variables found not to correlate (P > 0.05) with site index as single-level variables. Humus type (AIC = 21,749), depth of soil organic layer (AIC = 21,968, r = −0.37), drainage (AIC = 22,337), Shannon index (AIC = 20,788, r = 0.61), Simpson index (AIC = 21,213, r = 0.55), spruce and aspen basal area proportions (AIC = 21,321, r = −0.53; AIC = 21,035, r = 0.58 respectively), were the major (explaining at least 5 % of variability in site index) transient site variables identified in this study. The major permanent site variables included soil surface deposit (AIC = 22,197), and soil texture of the B-horizon (AIC = 22,156).
Once soil surface deposit was controlled for with different models (Table S3, Eqs. 6a–d), variability in site index was explained by transient factors only (Shannon index, stand age, aspen and jack pine basal area proportions).
The extent of SI control by factors changed from one deposit type to another (Table 1).
H2
SI is homogenous within regional units
Even though we had fire data spanning from 1924 to 2009, we did not observe a major (R2 = 0.3 %) contribution of burn rate to SI in the study region at that scale of analysis. Mean vapour pressure deficit at landscape scale (R2 = 5 %) and mean monthly water balance (R2 = 7 %) were major climatic drivers of SI. Spruce and aspen plot proportions within the landscape (R2 = 11, 13 % respectively) and landscape mean age (R2 = 9 %) were the major transient variables. Dominant soil surface deposit (R2 = 6 %) and dominant soil texture (R2 = 6 %) were the most important permanent site factors that explained SI. By controlling for landscape-level dominant surface deposit types with different models, we observed that a combination of transient factors (Shannon index, stand age, aspen and jack pine basal area proportions and degree days) best explained the observed variability in SI (Table 2):
The extent of control by SI drivers (absolute amounts of explained variance) as well as relative contributions to total explained variance (Table 1 vs. Table 2) varied when the two sets of models were compared, showing the importance of scale consideration in SI dynamics.
H3
SI is driven differently by bioclimatic domain
Within the mixedwood region, a combination of transient site factors (Shannon index, stand age, basal area proportions of aspen and jack pine) explained 69 % of the variability in productivity (Table S3, Eq. 8a, Table 3). Within conifer landscapes, we did not observe a major (<5 %) contribution of permanent site features to the total explained variance either. Instead, Shannon index, basal area proportion of aspen, stand age, depth of organic layer and 50-year mean growing degree-days accounted for SI dynamics (Table S3, Eq. 8b; Table 3):
Also within mixedwood, aspen basal area proportion was more important than Shannon index but within conifer landscapes the inverse was the case (Table 3, also Fig. 2a, b). Thus, SI seems to be driven to different extents by transient site features and also differently by bioclimatic domain (species richness).
H4
A single global model is inadequate
When soil or bioclimatic groupings were not taken into account, variability in SI was explained by soil surface deposit type, Shannon index, stand age, aspen and jack pine basal area proportions (Table S3, Eq. 9), which we refer to as the global model.
Optimization with integer programming indicated that alternative models were distinct from the global model (Table S3, Eq. 9); this was the case for alternative model sets combining plot-level soil deposit types (Table S3, Eqs. 6a–6d), dominant soil deposit types (Table S3, Eqs. 7a–7d) and also true for regrouping by bioclimatic domains (Table S3, Eqs. 8a, b) which respectively led to differences in AIC of 34, 19 and 285 relative to the AIC value of the global model (Table S5). The distinction between bioclimatic domains seems important in explaining SI. Furthermore, a combination of models by plot-level soil deposit types (till and clay) and bioclimatic domains led to the maximum difference (Table S5) in AIC relative to the global model. In this latter optimization procedure, plot-level coarse and organic soil deposits types were not highlighted, showing that when bioclimatic domain (species richness) is already accounted for in a productivity model, taking plot-level till and clay deposits into account further improved the model.
Discussion
Site index control differs with scale
Results show differences in total variance explained by SI with changes in substrate types and also with bioclimatic domain (species richness) (Fig. 2). Mean SI was highest (17 m) on clay surface deposit type and lowest on organic deposits (12 m) and varied in-between these extremes on coarse (15 m) and till (13 m) deposit types. Long-term (50 year) mean of monthly moisture balance was slightly higher (26 mm month−1) on clay deposit type relative to the others (coarse; 23 mm month−1, till and organic; 25 mm month−1). Also, mean depth of organic layer was highest on organic deposits (77 cm) and lowest on clay (14 cm) and coarse deposits (13 cm). Consequently, specific area of mineral soil layer, a variable that is linked to mineral weathering and therefore directly related to productivity (Hamel et al. 2004) was found to be highest (73,403) on clay deposit type, and lowest on organic deposits (6,144) while moderate on till (51,895) and coarse (28,167) deposits.
Delineation by substrate type adheres with earlier observations that reported differing successional changes among soil surface deposit types, with higher transition rates on cochrane tills (according to Dubé-Loubert et al. 2013, cochrane tills were deposited by eastward to southeastward ice re-advances of the Hudson Dome into the Lake Ojibway basin around the end of the deglaciation, across the Paleozoic sedimentary rocks of the Hudson platform) and slower transition rates on coarse deposit types (Belleau et al. 2011). The diversity-pedogenesis hypothesis stipulates that plant diversity is influenced by soil type and stand age (Laliberté et al. 2013). Variability in total variance explained by bioclimatic domain is thus due to regional differences in substrate (Table 1), but climatic and disturbance factors that result in differences in stand dynamics between these domains (Jasinski and Payette 2005) also play major roles. For instance, annual sums of degree days explained substantial variability in SI and was retained in the conifer model (Table S3, Eq. 8b) but not the model for mixedwoods. Also, mean moisture balance (25 mm month−1) was higher within mixedwoods compared to conifer dominated landscapes (21 mm month−1) and correspondingly, a slightly higher mean SI was observed in the former (16 m) relative to the latter (15 m).
We observed differences in productivity control by scale (Tables 1, 2, 3). Similarly, the relative contribution of drivers to total explained variance also consistently varied with scale, and this is the first time such spatial differential effects in SI responses to indirect climatic and site factors have been reported. Signal changes on dominant coarse and organic sites relative to stand-level coarse and organic substrate types point to a regional effect of dominant substrate types. Succession rates were reported to be slowest on coarse deposit types due to the xeric nature of such soils as observed in this study. Organic soil deposits are known to be prone to successional paludification (Simard et al. 2007) and as reported earlier, the average organic layer thickness on this substrate type is about five times the mean calculated across all sites. Soil water regime coupled with reduced mineralization due to paludification therefore explains the regionality in the substrate effect on stand-level productivity. Weakening of productivity-driver relationships when regional till sites were taken into account might be related to the relatively high rate at which cohort transitions occur on this substrate type (Belleau et al. 2011), such that accounting for regional substrate could mean combining stands at varied stages of succession, thereby leading to a decrease in correlation. Clay is known to be a productive substrate for boreal forest stands, but it is not entirely clear why scale effect was not obvious on this substrate type.
Deposits tend to be spatially aggregated in different regions such that a co-variation of temperature with soil deposit (R2 = 7 %, P = 0.000) was observed, with higher means on coarse deposits but higher variability on organic deposit types (Fig. 3a). Warming therefore, caused changes in productivity-driver relationships especially on moisture stressed and xeric (coarse deposits) sites relative to e.g. organic sites. Areas of similar disturbance regime also exhibited some homogeneity in productivity that influenced correlations at the regional level such that combining areas of varied fire regimes led to a decrease in variance explained. Indeed, we observed that dominant deposit type co-varied with fire cycle (R2 = 31 %, P = 0.000, Fig. 3b).

Spatial distribution of dominant deposit types (A), mean temperature in °C (B) and fire cycle in years (C). Till and clay deposit types were spatially aggregated in a way that no significant differences in annual mean temperatures were observed. Fire cycle varied substantially from one deposit type to another (P < 0.01)
A combination of models better reflects existing spatial heterogeneity in productivity
Irrespective of the cause of the differences in correlation by scale discussed in earlier paragraphs, there is a need for a modelling approach that accounts for this multiple/hierarchical variability in sensitivities. Existing models (e.g. McKenney and Pedlar 2003; Pinto et al. 2008; Coops et al. 2011) are single-level models that assume equal sensitivities across spatial scales and therefore, impose an equality of means on the calibration data. Variable sensitivity means that the use of a single model would be inadequate since it leads to underestimating productivity on till sites where accounting for regional effects leads to a decrease in signal and overestimating productivity for coarse and organic deposit types where sensitivities increase. We observed a marked difference in AIC between alternative models and the global model. To maximize the difference in AIC between the global and an alternative model, the latter must account for heterogeneity as a result of plot-level deposit types, particularly till and clay deposits and bioclimatic domain (i.e. species richness). Indeed, there were major differences in productivity drivers between the models by bioclimatic domains. In the region with less species richness (conifer-dominated), stand diameter structure was the main driver of productivity and the proportion of basal area that is aspen was the major productivity driver within mixedwood (species-rich region, Table 2). We also saw distinct variability in productivity with ageing. For northern latitudes and particularly conifer-dominated landscapes, one would have expected that either annual or seasonal temperature would be the most limiting growth factor (Xu et al. 2013) but this was not the case. As structural and compositional changes are dynamics that characterize stand succession, it is inferred that successional changes, more than climate, drive productivity when measured with SI. Given that potential height growth differs among species, we suspect that this could influence the effect of species abundance on SI. While Shannon index-height growth relationship has been recognized (Ouzennou et al. 2008; Anyomi et al. 2013), we suspect that the importance of Shannon index could be slightly inflated, due to the nature of the site index model (Eq. 1). But the fact that neither species abundance nor stand structural effects was consistently the first entry variable across model (Table 3) shows there was no systematic bias. Finally, results also demonstrate heterogeneity in productivity–driver relationships to the extent that a combination of models calibrated at identified gradients of heterogeneity performed better when compared to a single (global) model. There is a school of thought that envisages forest ecosystems as composed of several components that interact with each other and the external environment in a way that produces heterogeneous structures that are capable of adapting through time (Puettmann et al. 2013, p. 6). Our results demonstrate the existence of heterogeneity in forest productivity due to species interaction (stand dynamics) and, therefore, concur with some aspects of this concept. But further research will be needed in order to verify if such a system could be described as a complex adaptive one.
Implications within the context of climate change
Positive effects of aspen and jack pine proportions on productivity adds to a growing body of evidence on complementarity among species within natural boreal ecosystems and yet differently because a long-term productivity measure (site index) was used in this study. The presence of one species improves the growing conditions that benefit other species or a scenario where co-existing species occupy different niches. Either way, complementarity has implications for ecosystem resilience to disturbances (Turner et al. 1999). IPCC (2007) projected about 10–20 % increase in precipitation and 3–4.8 °C increase in warming for eastern North America by the end of the 21st century. A recent study projected conifer forest contraction and an expansion in oaks, hickories, birch and aspen forests by the end of the 21st century (Tang et al. 2012) due to a warming climate, but there could also be non-climatic constraints (Iversion and Mackenzie 2013). Natural disturbances and forest management activities have also altered species composition over time. For instance, Kelly et al. (2013) reported high burn rates within the boreal forest in recent decades and since aspen is an early successional and shade intolerant species, largely propagated by fire, it is expected that aspen forests should expand. Given that hardwoods are less flammable relative to conifers, there are calls for the use of mixedwood and hardwood forests in dealing with fire risks in the boreal forest (Girardin et al. 2013). An expansion of hardwood forests, whether as result of direct effects of warming or through climate change driven increased biomass burning or as a result of management intervention (assisted expansion), would likely result in an increase in the proportion of aspen. Therefore, we postulate that boreal forest site productivity will significantly increase as a consequence.
In this study, correlations between SI, climate, soil and stand attributes were explored for conifer and mix of conifer and deciduous landscapes in Quebec. Correlative approaches may not always be associated with clear causal processes but they provide important hints as to the processes that produce observable patterns across landscapes. SI, a widely used measure of productivity can be derived from stem analysis data. It can also be estimated from trees that have been in dominant canopy position and therefore have not experienced previous growth suppression. SI can also be modelled using largest diameter trees within a stand. SI obtained by any of these approaches are comparable even though some differences might exist (Mailly et al. 2004). It is not clear how estimating SI using one or another of the above methods might introduce errors in SI estimation. But a species aspecific SI, as estimated here, is expected to better reflect site productive potential regardless of the approach used. The use of chronosequence data with temporary sample plot data to infer stand dynamics of forest stands rather than long-term measurements of vegetation changes could potentially affect our results (Johnson and Miyanishi 2008). Since inventory design was such that the most common forest types were sampled, regions with higher species richness were likely to have higher plot densities relative to areas of less species richness (Fig. 1), and this could potentially affect the results.
Conclusion
This study shows that productivity dynamics can be better inferred when micro, meso and macroscale variability in driving factors are taken into consideration, demonstrating the importance of accounting for heterogeneity when inferring processes from patterns. Therefore, caution must be taken in the application of model predictions that fail to account for these heterogeneities, especially if modelled stands go through various successional stages. Results show the dominance of stand dynamics effects on productivity and a positive effect of aspen abundance on site index.
References
Aertsen W, Kint V, Muys B, Orshoven JV (2011) Effects of scale and scaling in predictive modelling of forest site productivity. Environ Model Softw 31:19–27
Anyomi K, Raulier F, Mailly D, Girardin M, Bergeron Y (2012) Using height growth to model local and regional response of trembling aspen (Populus tremuloides Michx.) to climate within the boreal forest of western Québec. Ecol Model 243:123–132
Anyomi K, Raulier F, Bergeron Y, Mailly D (2013) The predominance of stand composition and structure over direct climatic and site effects in explaining aspen (Populus tremuloides Michaux) productivity within boreal and temperate forests of western Quebec, Canada. For Ecol Manage 302:390–403
Belleau A, Leduc A, Lecomte N, Bergeron Y (2011) Forest succession rate and pathways on different surface deposit types in the boreal forest of northwestern Quebec. Ecoscience 18(4):329–340
Bergeron J-F, Saucier J-P, Robert D, Robitaille A (1992) Québec forest ecological classification program. For Chron 68(1):53–63
Bontemps J-D, Hervé J-C, Dhote J-F (2009) Long-term changes in forest productivity: consistent assessment in even-aged stands. For Sci 55(6):549–564
Boucher D, Gauthier S, De Grandpré L (2006) Structural changes in coniferous stands along a chronosequence and productivity gradient in the northeastern boreal forest of Québec. Ecoscience 13(2):172–180
Coops NC, Gaulton R, Waring RH (2011) Mapping site indices for five Pacific Northwest conifers using a physiologically based model. Appl Veg Sci 14:268–276
Curtis RO, Post BW (1964) Basal area, volume, and diameter related to site index and age in unmanaged even-aged northern hardwoods in the Green Mountains. J Forest 62:864–870
Dormaan CF, McPherson JM, Araujo MB, Bivand R, Bolliger J, Carl G, Davies RG, Hirzel A, Jetz W, Kissling DW, Kuhn I, Ohlemuller R, Peres-Neto PR, Reineking B, Schroder B, Schurr FM, Wilson R (2007) Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography 30:609–628
Dubé-Loubert H, Roy M, Allard G, Lamothe M, Veillette JJ (2013) Glacial and non-glacial events in the eastern James Bay Lowlands, Canada. Can J Earth Sci 50(4):379–396
Dunne T, Leopold LB (1978) Water in environmental planning. W.H. Freeman and Co., San Francisco, p 818
Girardin MP, Ali AA, Carcaillet C, Blarquez O, Hely C, Terrier A, Genries A, Bergeron Y (2013) Vegetation limits the impact of a warm climate on boreal wildfires. New Phytol. doi:10.1111/nph.12322
Hamel B, Bélanger N, Paré D (2004) Productivity of black spruce and Jack pine stands in Quebec as related to climate, site biological features and soil properties. For Ecol Manage 191:239–251
IPCC (Intergovernmental Panel on Climate Change) 2007. Synthesis report. Summary for Policy makers. http://www.ipcc.ch/pdf/assessment-report/ar4/syr/ar4_syr.pdf Accessed on 18 February, 2010
Iversion LR, Mackenzie D (2013) Tree-species range shifts in a changing climate: detecting, modeling, assisting. Landscape Ecol 28:879–889
Jasinski JPP, Payette S (2005) The creation of alternative stable states in the southern boreal forests, Quebec, Canada. Ecol Monogr 75(4):561–583
Johnson EA, Miyanishi K (2008) Testing the assumptions of chronosequence in succession. Ecol Lett 11:419–431
Kelly R, Chipman ML, Higuera PE, Stefanova I, Brubaker LB, Hu FS (2013) Recent burning of boreal forests exceeds fire regime limits of the past 10,000 years. Proc Natl Acad Sci USA. doi:10.1073/pnas.1305069110
Laliberté E, Grace JB, Huston MA, Lambers H, Teste FP, Turner BL, Wardle DA (2013) How does pedogenesis drive plant diversity? Trends Evol Ecol 1676:1–10
Landsberg J, Sands P (2011) Physiological ecology of forest production: principles, processes and models. Academic Press, London
Larson AJ, Lutz JA, Gersonde RF, Franklin JF, Hietpas FF (2008) Potential site productivity influences the rate of forest structural development. Ecol Appl 18(4):899–910
Lexerød NL, Eid T (2006) An evaluation of different diameter diversity indices based on criteria related to forest management planning. For Ecol Manage 222:17–28
Mailly D, Turbis S, Auger I, Pothier D (2004) The influence of site tree selection method on site index determination and yield prediction in black spruce stands in northeastern Québec. For Chron 80(1):134–140
Mailly D, Gaudreault M, Picher G, Auger I, Pothier D (2009) A comparison of mortality rates between top height trees and average site trees. Ann For Sci 66:202
Mansuy N, Gauthier S, Robitaille A, Bergeron Y (2010) The effects of surficial deposit-drainage combinations on spatial variations of fire cycles in the boreal forest of eastern Canada. Int J Wildland Fire 19(8):1083–1098
McKenney DW, Pedlar JH (2003) Spatial models of site index based on climate and soil properties for two boreal tree species in Ontario, Canada. For Ecol Manage 175:497–507
Monserud R (1984) Problems with site index: an opinionated review. In: Forest land classification: experiences, problems, perspectives. Proceedings of a symposium held at the University of Wisconsin, Madison. March 18–20, 1984
NFI (National Forestry Database), 2009. http://nfdp.ccfm.org/inventory/background_e.php#2.2. Last accessed on 14 August 2012
O’Neil G, Hamann A, Wang T (2008) Accounting for population variation improves estimates of the impact of climate change on species’ growth and distribution. J Appl Ecol 45:1040–1049
Ouzennou H, Pothier D, Raulier F (2008) Adjustment of the age-height relationship for uneven-aged black spruce stands. Can J For Res 38:2003–2012
Pinto EP, Gégout J-C, Hervé J-C, Dhote J-F (2008) Respective importance of ecological conditions and stand composition on Abies alba Mill. dominant height growth. For Ecol Manage 255:619–629
Pothier D, Auger I (2009) NATURA-2009: un modèle de prévision de la croissance à l’échelle du peuplement pour les forêts du Québec. Mémoire de recherche forestière no. 163. Direction de la recherche forestière p 76
Prescott CE, Zabek LM, Staley CL, Kabzems R (2000) Decomposition of broadleaf and needle litter in forests of British Columbia: influences of litter type, forest type, and litter mixtures. Can J For Res 30:1742–1750
Puettmann KJ, Messier C, Coates KD (2013) Managing forests as complex adaptive systems; introductory concepts and applications. In: Messier C, Puettmann KJ, Coates KD (eds) Managing forests as complex adaptive systems; building resilience to the challenge of global change EarthScan. Routledge, Abingdon, p 353
Raulier F, Lambert M-C, Pothier P, Ung C-H (2003) Impact of dominant tree dynamics on site index curves. For Ecol Manage 184:65–78
Régnière J, Saint-Amant R (2008) BioSIM 9 - Manuel de l’utilisateur. Ressources naturelles Canada, Service canadien des forêts. Centre de foresterie des Laurentides. Rapport d’information LAU-X-134F. ftp://cfl.scf.rncan.gc.ca/regniere/Data/Weather/. Accessed on 4 February 2011
Robitaille A, Saucier J-P (1998) Paysages régionaux du Quebec méridional. MRN (Ministère des Ressources naturelles), Gouvernement du Québec. Les publications du Québec, Québec, p 213
Rouleau R (1994) Comparaison entre les programmes d’inventaire de 1970 à aujourd’hui. Unpublished legend explaining MRNFQ’s inventory programme. p 106
Simard M, Lecomte N, Bergeron Y, Bernier PY, Pare D (2007) Forest productivity decline caused by successional paludification of boreal soils. Ecol Appl 17(6):1619–1637
Tang G, Beckage B, Smith B (2012) The potential transient dynamics of forests in New England under historical and projected future climate change. Clim Change 114:357–377
Trap J, Bureau F, Brethes A, Jabiol B, Ponge J-F, Chauvat M, Decaëns T, Aubert M (2011) Does moder development along a pure beech (Fagus Sylvatica L.) chronosequence result from changes in litter production or in decomposition rates. Soil Biol Biochem 43(7):1490–1497
Turner MG, Romme WH, Gardner RH (1999) Prefire heterogeneity, fire severity, and early postfire plant reestablishment in subalpine forests of Yellowstone national park, Wyoming. Int J Wildland Fire 9(1):21–36
Vallet P, Perot T (2011) Silver fir stand productivity is enhanced when mixed with Norway spruce: evidence based on large scale inventory data and a generic modelling approach. J Veg Sci 22:932–942
Vila M, Carrillo-Gavila A, Vayreda J, Bugmann H, Fridman J, Grodzki W, Haase J, Kunstler G, Schelhaas M, Trasobares A (2013) Disentangling biodiversity and climatic determinants of wood production. PLoS One 8(2):e53530. doi:10.1371/journal.pone.0053530
Wang Z, Grant RF, Arain MA, Bernier PY, Chen B, Chen JM, Govind A, Guindon L, Kurz WA, Peng C, Price DT, Stinson G, Sun J, Trofymowe JA, Yeluripati J (2013) Incorporating weather sensitivity in inventory-based estimates of boreal forest productivity: a meta-analysis of process model results. Ecol Model 260:25–35
Weiskittel AR, Hann DW, Kershaw JA, Vanclay JK (2011) Forest growth and yield modeling. Wiley-Blackwell, Chichester, p 415
Xu L, Myneni RB, Chapin FS III, Callaghan TV, Pinzon JE, Tucker CJ, Zhu Z, Bi J, Ciais P, Tømmervik H, Euskirchen ES, Forbes BC, Piao SL, Anderson BT, Ganguly S, Nemani RR, Goetz SJ, Beck PSA, Bunn AG, Cao C, Stroeve JC (2013) Temperature and vegetation seasonality diminishment over northern lands. Nature Clim Change. doi:10.1038/NCLIMATE1836
Acknowledgments
We sincerely thank the Ministry of Natural Resources of Quebec for inventory and fire data. We also thank Nicole Stump of Wayne National Forest, Ohio US for proof reading the paper. Funding for the study came from an NSERC (Natural Sciences and Engineering Research Council of Canada) project grant.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Anyomi, K.A., Raulier, F., Bergeron, Y. et al. Spatial and temporal heterogeneity of forest site productivity drivers: a case study within the eastern boreal forests of Canada. Landscape Ecol 29, 905–918 (2014). https://doi.org/10.1007/s10980-014-0026-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-014-0026-y