
Abstract
Two alkylated 1,4-benzoquinones were identified from the defensive secretion produced by the neotropical harvestman Goniosoma longipes (Gonyleptidae). They were characterized as 2,3-dimethyl-1,4-benzoquinone and 2-ethyl-3-methyl-1,4-benzoquinone. We tested the effectiveness of these benzoquinone secretions against several predator types, including invertebrates and vertebrates. Different predators were exposed to the harvestmen's gland secretion or to distilled water in laboratory bioassays. Our results indicate that secretions containing the 1,4-benzoquinones released by G. longipes can be an effective defense against predation, and that the effectiveness of the secretion is dependent on the predator type. The scent gland secretion repelled seven ant species, two species of large wandering spiders, and one frog species, but was not an effective defense against an opossum. Our study also demonstrates that the scent gland secretion of G. longipes can work as a chemical shield preventing the approach of three large predatory ants for at least 10 min. The chemical shield may protect the harvestman against successive attacks of the same ant worker and also allow the harvestman to flee before massive ant recruitment. Our data support the suggestion that chemical defenses may increase survival with some but not all potential predators. This variation in defense effectiveness may result from many interacting factors, including the attack strategy, size, learning ability, and physiology of the predators, as well as the chemical nature of the defensive compounds, type of emission, and amount of effluent released by the prey.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The species of the order Opiliones are characterized by the presence of a pair of exocrine glands located at the anterior margins of the cephalothorax near the base of the second pair of legs (Shultz, 1990). These glands produce a variety of volatile secretions that are released under the threat of predation (Eisner et al., 1978). To date, nearly 30 species have had their secretions chemically characterized, and a phylogenetic pattern may be observed. Species belonging to the suborder Eupnoi produce a variety of acyclic compounds, such as short-chain ketones and alcohols among the sclerosomatids and naphthoquinones among the phalangiids (references in Roach et al., 1980). In the suborder Laniatores, species of the superfamily Travunioidea produce mainly terpenoids, whereas among the Gonyleptoidea, the secretion is composed of phenols and alkylated 1,4-benzoquinones (references in Eisner et al., 1978; Acosta et al., 1993; Gnaspini and Cavalheiro, 1998). Quinones are widely distributed in the scent gland secretions of arthropods such as insects and millipedes (Blum, 1981), but the particular alkylated quinones produced by harvestmen seem to be of more restricted occurrence (Eisner et al., 1978).
Although much is known about the chemical nature of the compounds produced by harvestmen, knowledge about their biological role is still scarce. Several authors suggest that the scent gland secretions in harvestmen could be used for intraspecific communication, including trail marking, sexual recognition, mutual attraction, and alarm (see Holmberg, 1986; Machado et al., 2002). However, the main function attributed to the scent secretion is defense against predators (review in Holmberg, 1986). Laboratory observations have shown that ants are repelled by the secretion released by the cosmetid Vonones sayi (Eisner et al., 1971), the leiobunine Leiobunum formosum (Blum and Edgar, 1981), and the gonyleptid Acanthopachylus aculeatus (Eisner et al., 2004). Anecdotal observations indicate that some spiders may be also deterred by harvestman secretions (e.g., Cloudsley-Thompson, 1958; Juberthie, 1976), but others are not (e.g., Bristowe, 1949; Immel, 1954). Recently, Eisner et al. (2004) demonstrated that the scent gland secretion of the harvestman A. aculeatus was unable to deter the attack of a lycosid spider. The effectiveness of the scent gland secretion against vertebrates is also controversial because some species of frog seem to present a strong aversive response (e.g., Edgar, 1971), whereas some mammals feast on harvestmen (e.g., Pelegatti-Franco and Gnaspini, 1996). Despite these field and laboratory observations, the defensive role of the secretion against invertebrate and especially vertebrate predators remains to be experimentally tested through a rigorous protocol.
In this study, we chemically characterized the scent secretion of Goniosoma longipes (Roewer, 1913; Gonyleptidae) and conducted a series of bioassays to test the effectiveness of the 1,4-benzoquinone secretions produced by this neotropical harvestman against several predator types. This is a comprehensive study on the defensive role of the fluids released by harvestmen and the first to assess experimentally that the specific alkylated benzoquinone secretions released by harvestmen can deter vertebrates. Because after releasing the odoriferous secretion, residual effluents remain on the harvestmen's body (Eisner et al., 1971), we also tested if the secretion can also work as a “chemical shield”.
Methods and Materials
Studied Species
The harvestman G. longipes is conspicuous (Figure 1a, b) and widespread in forested areas of southeastern Brazil. This species was chosen as the source of secretion because it is abundant in our study site and because individuals produce a great amount of secretion when disturbed (see Machado et al., 2000). Moreover, the ecology and behavior of the species were previously studied (see Machado and Oliveira, 1998; Machado et al., 2000), aiding the selection of more realistic potential predators. The external morphology of the ozopore of G. longipes has been described (see Figures 19 and 20 in Hara and Gnaspini, 2003), and the mode of emission occurs as follows: (a) first, a large transparent droplet of enteric fluid composed exclusively of water is regurgitated, slips along the lateral area of the dorsal scute, runs by capillarity in tegumentary grooves, and accumulates in the posterior portion of the body (Figure 1a); (b) if the harvestman is persistently disturbed, a second fluid may be secreted by the odoriferous glands, adding to the previously colorless fluid a yellowish coloration and a characteristic sour smell (Figure 1b). Sometimes, the secretion may also be sprayed as a jet directly toward the aggressor without mixing with enteric fluid (Machado et al., 2000; Hara and Gnaspini, 2003).

Adult male of the harvestman G. longipes (A) with two droplets (arrows) of aqueous enteric fluid and (B) with a droplet of secretion (arrows) formed by the mixture of enteric fluid and defensive secretion (photos: B.A. Buzatto). The circles indicate the external opening of the scent gland. (C–F) Model predators used in the bioassays: (C) the hunting spiders E. cyclothorax (photo: R.J. Sawaya) and (D) T. biocellata (photo: G. Machado); (E) the small horned frog Pr. boiei (photo: I. Sazima) and (F) the white-eared opossum D. albiventris (photo: E.G. Martins).
All individuals of G. longipes used in this study were collected inside caves at the Parque Florestal do Itapetinga (23°15′S, 46°45′W, 900- to 1300-m altitude, 1600 mm rain/year), Atibaia, São Paulo state, southeastern Brazil (for details on the study site, see Machado and Oliveira, 1998 and Machado et al., 2000). In the laboratory, adult G. longipes were maintained in a communal terrarium (60 × 40 cm base, 35 cm high) containing cotton wetted with water to maintain the humidity and fed with dead insects twice a week.
Analyses of Defensive Secretions
Chemical secretions from G. longipes (about 8 mg/individual) were collected by pressing cotton wools (50–70 mg) onto the gland openings. The exudates were then either extracted with ethyl acetate (about 5 ml) and analyzed by gas chromatography–mass spectrometry (GC-MS) or with CDCl3/tetramethylsilane (TMS) (600 μl) for the NMR experiments. GC-MS analyses were carried out using an HP 6890/5973 system equipped with an HP5 fused silica capillary column (30 m × 0.25 mm × 0.25 μm). Oven temperatures were programmed from 50 to 200°C at 10°C min−1, and subsequently from 200 to 290°C at 16°C min−1. The injector temperature was 250°C. Helium was used as carrier gas at a flow rate of 1 ml min−1. The electron impact mass spectrum (EIMS) were taken at 70 eV. Scanning speed was 3.54 scans s−1 from m/z 40 to 450. High-resolution mass spectrum was obtained from VG AutoSpec EI (70 eV) equipment. Nuclear magnetic resonance (NMR) spectral data were acquired from Varian Inova spectrometer, operating at 499.88 MHz for 1H-NMR and 125.71 MHz for 13C-NMR. CDCl3 was used as solvent and TMS as an internal reference (δ = 0.0 ppm). Chemical shifts δ were recorded in ppm and coupling constants J in Hz. Thin-layer chromatography analyses were performed on Merck F254 Aluminum Sheets, and the spots were visualized by UV light (254 nm). All solvents were of high analytical grade, bidistilled before using. Cotton wools were successively extracted with ethyl acetate, and the solvent was evaporated under vacuum.
Bioassays
The experiments were conducted from October 2002 to May 2003 and were performed between 19:00 and 23:00 hr under dim light conditions. Immediately before each trial, an adult individual of G. longipes was milked of secretion by seizing it by hand. Males and nonovigerous females were milked because the total amount of secretion released by them is not different (8.3 ± 4.4 mg). The exudate was collected with capillary tubes and diluted into a syringe with 100 μl of distilled water. The secretion was mixed with water because many harvestman species, including G. longipes, dilute their own gland secretion in aqueous enteric fluid (Eisner et al., 1971, 1978; Acosta et al., 1993; Gnaspini and Cavalheiro, 1998; Hara and Gnaspini, 2003; see also Figure 1a). Different syringes were used for each experimental group. A different unmilked individual was used in each trial because repeated milking of the same individual could reduce the concentration and quantities of the secretion (Eisner et al., 1971). Different model predators were exposed to the harvestmen's gland secretion (treatment) or to distilled water (control).
Most laniatorean harvestmen, including G. longipes, have spines covering the body and legs and present a variety of defensive behaviors that may be divided in evasive responses (fleeing and dropping from the substrate) and aggressive responses (Machado, 2002). The latter includes several mechanisms that threaten, harm, or deter aggressors, such as attacking with the pedipalps, biting with the chelicerae, pinching with the sharp projections of the femora and coxae IV, and releasing scent gland secretions (Machado, 2002; Hara and Gnaspini, 2003). To test the potential defensive role of the glandular secretion only, and prevent other possible defensive mechanisms, individuals of G. longipes were not offered to the predators. Rather, the common cricket Gryllus gryllus (nearly 1 cm of body length) was used as model of prey in the experiments with spiders, frogs, and marsupials (see details below).
Tests with Ants
Two bioassays were conducted with ants: one in the field to test the potential of the scent gland secretion as a repellent and another in the laboratory to test the effectiveness as a “chemical shield”. The field experiment consisted of the presentation of filter paper baits imbibed with honey solution, placed on plastic dishes (5-cm diam). These baits were randomly distributed on the forest floor 5 m from each other. In the treatment group, when ants had come to feed at the margin of the bait, the glandular secretion of one individual of G. longipes diluted in 100 μl of water was discharged in the center of the filter paper with a syringe. A similar procedure was used in the control group, but only 100 μl of water were discharged in the bait. In both experimental groups, the number of ants in contact with the bait was counted before and 5 sec after presentation of either secretion or water. Five ant species attended the baits in the field and were used in this experiment: (1) Formicinae—Camponotus lespesi (four baits; two controls and two treatments); (2) Myrmicinae—Pheidole sp. (eight baits; four controls and four treatments); (3) Ponerinae—Gnamptogenys sp. (eight baits; four controls and four treatments), Odontomachus chelifer (six baits; three controls and three treatments), and Pachycondyla striata (six baits; three controls and three treatments). After square-root transformations of the data, a repeated-measures ANOVA was performed on the number of workers of the five ant species in contact with the baits before and after the treatment in both experimental groups (Pagano, 1994).
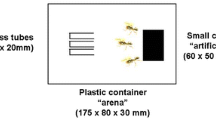
Three ant species were used in the laboratory experiments: Pachycondyla villosa (one colony), Odontomachus hastatus, and Camponotus crassus (three colonies each). Inside the tray of each colony, we presented a glass coverslip (1 × 5 cm) divided in two sides randomly designated by the flip of a coin as treatment or control. The treatment consisted of a filter paper (1 cm2) wetted with 200 μl of a saturated sugar solution mixed with the glandular secretion of one individual of G. longipes. The control side contained only a filter paper (1 cm2) wetted with 200 μl of a saturated sugar solution. The ant's response was evaluated by counting the total number of workers feeding at the baits at 1-min intervals during 10 min after the first contact.
Tests with Spiders
Enoploctenus cyclothorax (Ctenidae) is a large (3- to 5-cm body length) wandering spider that is commonly found near cave entrances (Figure 1c) and occasionally preys on adult G. longipes (Machado et al., 2000; Machado, personal observations), as well as other gonyleptid harvestmen (Willemart and Kaneto, 2004). Trechalea biocellata (Trechaleidae) is also a large (2- to 3-cm body length) wandering spider, abundant in the study site (Figure 1d), which is generally found near river margins or in other moist habitats, such as caves. An adult male of this spider was once observed preying on a harvestman, probably a Discocyrtus (Gonyleptidae), at night (Machado, personal observations). Individuals of both spiders were collected at the Parque Florestal do Itapetinga, from October 2002 to May 2003. They were maintained in individual cages (20 × 10 cm base, 15 cm high) containing cotton wetted with water to maintain the humidity and starved for 5–6 d before the experiments. Only subadults and adults of both sexes of T. biocellata (N = 40) and E. cyclothorax (N = 20) were used in the experiments. Individuals were offered a cricket, which they promptly took as prey. To ensure that the crickets were unable to promote injuries on the spiders, their hind legs (armed with several spines) were removed just before the experiment. After the spiders had started feeding, each individual was stimulated with either secretion of one harvestman diluted in 100 μl of water (treatment) or the same volume of distilled water (control), applied with a syringe directly to the base of the chelicerae. Individuals that extricated the chelicerae and abandoned the prey within 5 min were scored as respondents (cf. Eisner et al., 1997). The number of spiders that released the prey was compared between the two experimental groups using a Fisher exact test (Pagano, 1994). The time between the presentation of secretion and the response of the spiders was compared between species using a Mann–Whitney U-test.
Tests with Frogs
The effectiveness of the scent secretion against frogs was tested using adults (16 males and 4 females) of the small horned frog, Proceratophrys boiei (Leptodactylidae). The species is a leaf-litter dweller whose adults may reach 5–7.5 cm of body length (Figure 1e). Because of its large mouth, the diet of this species includes large ground arthropods, such as spiders, cockroaches, beetles, and crickets (Giaretta et al., 1998; Teixeira and Coutinho, 2002). Although there is no record of harvestmen among the food items of Pr. boiei, other species of the genus, such as Pr. apendiculata, include harvestmen in their diet (Machado, personal observations). Small horned frogs (N = 20) were collected at the Parque Florestal do Itapetinga during the breeding season from November 2002 to January 2003. They were maintained in a communal terrarium (60 × 40 cm base, 35 cm high) and starved 3 d before the experiments. They were offered a cricket, which they promptly took as prey. As soon as a frog swallowed the cricket, we caught it by hand, forced its mouth open, and used a syringe to inject either secretion of one harvestman diluted in 100 μl of water (treatment) or the same volume of distilled water (control). Anurans generally catch their prey by a quick flip of their sticky tongue and may swallow chemically protected arthropods, such as millipedes or beetles, before the secretion of the prey is discharged (Eisner and Meinwald, 1966). Hence, the secretion might be released outside the mouth, inside the mouth, or even inside the stomach. Our experimental protocol simulated the mouth scenario. Individuals that regurgitated the prey within 5 min were scored as respondents. The number of frogs that regurgitated the prey was compared between the two experimental groups using a Fisher exact test (Pagano, 1994).
Tests with Opossums
The white-eared opossum Didelphis albiventris (Didelphidae) is a medium-sized mammal (0.5–2.0 kg; Figure 1f), whose omnivorous diet includes harvestmen (Cáceres and Monteiro-Filho, 2001; Cáceres, 2002). Although D. albiventris occurs at the Parque Florestal do Itapetinga, experimental individuals were collected at the Clube Náutico Araraquara (21°43′S, 48°01′W, 653-m altitude, 1300 mm rain/year), a forested area near Américo Brasiliense, São Paulo state. In this locality, the species is abundant, and juvenile individuals (150–450 g) can be trapped using Sherman live traps. The opossums (N = 8) were maintained in individual cages for 12 hr after collection, and 30 min before the experiment, they were transferred to a dark closed box (60 × 40 cm base, 35 cm high) and fed with a piece of banana to bring all of them to a similar hunger level.
The presentation of the prey was made by using a tube inserted in the box's wall that placed the cricket in the center of the cage. The first cricket introduced in the cage had a 4 × 8 mm piece of filter paper carefully placed under its wings (no glue was used), wetted with the glandular secretion of one individual of G. longipes diluted in 100 μl of water (treatment). Two minutes after the introduction of the first cricket, a second cricket with a piece of filter paper wetted with 100 μl of distilled water (control) under its wings was placed in the cage. Prey were offered in this sequence because if the first cricket was rejected and not the second, it would be possible to identify a rejection response. This second cricket was also a control of the hunger level of the opossums. We recorded the time elapsed between the presentation of the crickets and the attack of the opossums, as well as the behavioral response of the animals to each type of prey. Because species of the genus Goniosoma are capable of ejecting their secretion forcibly as a spray (Machado, 2002; Hara and Gnaspini, 2003), one adult individual of G. longipes was also introduced in each cage after the presentation of the crickets to investigate the response of the opossums to this special type of emission.
Vouchers
One of the trials with frogs and all trials with the marsupials were video recorded, digitalized, and are available from the authors upon request. Voucher specimens of all studied species (except D. albiventris) were deposited at the Museu de História Natural da Universidade Estadual de Campinas and Museu de Zoologia da Universidade de São Paulo, Brazil.
Results
Chemistry
Two alkylated 1,4-benzoquinones were identified from the defensive secretion of the harvestman G. longipes (Figures 2 and 3):

GC-MS analysis of male G. longipes defensive secretion. Peaks corresponding to compounds 2,3-dimethyl-1,4-benzoquinone and 2-ethyl-3-methyl-1,4-benzoquinone are indicated as (1) and (2), respectively.

Chemical defense compounds identified from the scent secretion of the harvestman G. longipes. The origin of the fragment m/z 54 in the EIMS spectra is also shown.
2,3-Dimethyl-1,4-benzoquinone (compound 1). GC-MS m/z 136 (100), 108 (57), 107 (56), 82 (44), 79 (52), 65 (5), 54 (50); HRMS (EI, 70 eV): M+ 136.05392 (C8H8O2; calc. 136.05244); 1H-NMR (CDCl3, TMS, 499.88 MHz) δ (ppm) = 2.04 (s, H-7 and H-8, 6H), 6.72 (s, H-5 and H-6, 2H); 13C-NMR δ (ppm) = 187.39 (C-1, C-4), 140.99 (C-2, C-3), 136.24 (C-5, C-6), 12.18 (C-7, C-8). Other spectroscopic methods as 13C-NMR DEPT-135 and DEPT-90, 1H–1H gCOSY, 1H–13C 1J HSQC, and 1H–13C nJ gHMBC were also employed (data not shown).
2-Ethyl-3-methyl-1,4-benzoquinone (compound 2). GC-MS m/z 150 (100), 135 (10), 122 (37), 121 (18), 107 (79), 82 (21), 79 (38), 67 (12), 54 (22); HRMS: 150.06824 (C9H10O2; calc. 150.06810). For further spectroscopic data as 1H-NMR (CDCl3, TMS, 499.88 MHz), 13C-NMR, DEPT-135 and DEPT-90, 1H–1H gCOSY, 1H–13C 1J HSQC, and 1H–13C 1J gHMBC, see Table 1.
The fragmentation pattern of compound 1 under EIMS was compared with literature (Budzikiewicz et al., 1967), and it was identical to the spectrum reported for 2,3-dimethyl-1,4-benzoquinone. Because of molecular symmetry, compound 1 displayed two singlets in 1H-NMR spectrum: at δ = 2.04 ppm for six methyl protons and at δ = 6.72 ppm for the other two aromatic protons. This spectral simplicity allowed the structural identification of the other component in the mixture.
Compound 2 fragmentation pattern under EIMS did not match any mass spectral data available for other 1,4-benzoquinones derivatives identified in arthropods. The use of one and bidimensional NMR techniques allowed almost complete chemical shift assignments for this compound, which was identified as 2-ethyl-3-methyl-1,4-benzoquinone. The molecular formula C9H10O2 was confirmed by high-resolution mass spectrometry (M+ at m/z 150.06824; calc. 150.06810). A CH2CH3 substituent was easily characterized by the A3X2 spin system (δ = 2.51 ppm for 2H and δ = 1.06 ppm for 3H) in the 1H-NMR spectrum, which was further confirmed by 1H, 1H homonuclear correlations spectrum (gCOSY). The substitution pattern of compound 2 was first suggested by the presence of the intense fragment at m/z 54 in the low-resolution mass spectrum, confirmed by HRMS (observed m/z 54.01036; calc. for C3H2O m/z 54.01056; Figure 3). This was corroborated by the H-7/C-3, H-7/C-1, H9/C-2, and H-9/C-4 3J correlations observed by 1H, 13C heteronuclear long-range correlations spectrum (gHMBC). Further spectroscopic methods as gHSQC (1H,13C 1J), DEPT-135, and DEPT-90 were also employed in chemical shift assignments for compound 2 (Table 1).
The chemical composition of defensive secretions from male and female individuals was the same in G. longipes, but the proportion of the two components is slightly different. Compound 2 always displayed smaller relative abundance in GC-MS analyses. This feature was also explored in the interpretation of the NMR experiments, in which compound 2 signals were always less intense than those from compound 1.
Tests with Ants
The presentation of scent gland secretion in the treatment group induced a marked reduction in the number of ants tending the sugar baits in the field (Figure 4). This reduction was not observed in the control baits (Figure 4). The repellent effect was observed for recruiting species such as Pheidole sp., C. lespesi, and Gnamptogenys sp. and also for solitary hunters such as O. chelifer and P. villosa. After leaving the treatment baits (but not the control baits), workers of all tested species typically cleaned their antennae and front legs.

Number of ants in the field tending sugar baits before (time = 0 sec) and after (time = 5 sec) stimulation with harvestman secretion (treatment) or distilled water (control). Data combined from 32 trials (16 controls and 16 treatments) on five species of ants. There was a significant effect of time (repeated-measures ANOVA, F = 13.77, P < 0.001) and experimental group (repeated-measures ANOVA, F = 39.61, P < 0.001).
In the laboratory, the number of ants tending control baits generally increased during the 10-min period, whereas few or no ant contacted treatment baits (Figure 5). The workers that touched the filter papers of the treatment baits presented a behavioral response similar to that observed in the field experiment, i.e., they immediately cleaned their antennae and front legs and did not return to the bait.

Number of visits over time of three ant species tending control baits of sugar solution (circular dots, full line) and treatment baits of sugar solution mixed with harvestman secretion (square dots, dotted line) in the laboratory. The numbers at the right lateral of the graphics indicate the size of the ant colonies (for C. crassus and Odontomachus, three colonies each, and for P. villosa, one colony). Because the number of ants in contact with treatment baits was generally zero, it is not possible to distinguish the three dotted lines.
Tests with Spiders
The frequency of individuals of E. cyclothorax (Fisher exact test, P = 0.003) and T. biocellata (Fisher exact test, P = 0.008) that released the prey in the treatment group was significantly higher than in the control group (Figure 6a, b). There was no difference between species in the frequency of individuals that released the prey in the treatment group (Fisher exact test, P = 0.43). Respondent individuals of both species extricated their fangs, dropped the prey, and rubbed their mouthparts against the substrate. The time elapsed between the presentation of the secretion and the release of the prey also did not differ between species (U = 23.0; P = 0.37) and ranged from 1 to 30 sec in E. cyclothorax and from 1 to 50 sec in T. biocellata.

Number of individuals of the spiders (A) E. cyclothorax and (B) T. biocellata that released (white bars) or not (black bars) the prey after stimulation with harvestman secretion (treatment) or distilled water (control). (C) Number of individuals of the small horned frog Pr. boiei that regurgitated (white bars) or not (black bars) the prey after stimulation with harvestman secretion (treatment) or distilled water (control).
Tests with Frogs
The frequency of individuals of Pr. boiei that regurgitated the prey in the treatment group was significantly higher than of those in the control group (Fisher exact test, P = 0.004; Figure 6c). Frogs in the treatment group presented clear aversion reactions such as scratching the tongue on the substrate, shutting the eyes, and strong abdominal contractions (Figure 7). These behavioral reactions occurred both before and after prey regurgitation. Interestingly, for all respondent individuals, the prey was regurgitated alive.

(A–H) Sequence extracted from videotape showing the reaction a small horned frog, Pr. boiei, to the scent gland secretion of the harvestman G. longipes. (A) The individual immediately before the test. (B) After the harvestman secretion was presented inside its mouth, the individual had abdominal contractions, and (C–E) regurgitated the cricket. (F–G) The frog scratched the mouth on the substrate and moved backward. (H) Nearly 30 sec after regurgitation, the individual still showed clear signs of aversion, maintaining the eyes shut and presenting strong abdominal contractions.
Tests with Opossums
All individuals of D. albiventris consumed the crickets of both experimental groups. There was no clear sign of aversion when the opossums ingested the crickets wetted with secretion. All opossums also ingested adults of the harvestman G. longipes. Four of five subadult individuals subdued the harvestmen using the front legs to step on it, which probably promoted the releasing of secretion. After that, the harvestmen were grasped with the front legs and consumed. One subadult and three juveniles subdued the harvestmen using the snout to bite the prey. The biting elicited the release of a jet of secretion that probably reached the nose and the eyes of the opossums. In these cases, the opossums responded to the discharge by releasing the prey and wiping their eyes and snout with their front legs for nearly 2 min. After this period, the opossums approached the dead body of the prey and ate it. In all cases, only the body of the harvestmen was consumed and the legs were discarded. During actual consumption, the opossum showed no clear signs of aversion, such as those presented by the small horned frogs.
Discussion
Like most representatives of the superfamily Gonyleptoidea, the defensive secretion of the harvestman G. longipes is a mixture of compounds. The occurrence of alkylated 1,4-benzoquinones in G. longipes is chemotaxonomically in agreement with the constitution of the defensive secretions previously reported for other Gonyleptidae (Estable et al., 1955; Fieser and Ardao, 1956; Roach et al., 1980; Acosta et al., 1993; Eisner et al., 2004), including congeneric species, such as Goniosoma spelaeum (Gnaspini and Cavalheiro, 1998). The derivative 2,3-dimethyl-1,4-benzoquinone has been already identified in other harvestman species belonging to the families Cosmetidae, Gonyleptidae, and Manaosbiidae (Estable et al., 1955; Fieser and Ardao, 1956; Eisner et al., 1977, 2004; Roach et al., 1980; Acosta et al., 1993), but this is the first report of 2-ethyl-3-methyl-1,4-benzoquinone as a natural product. Moreover, the use of bidimensional nuclear magnetic resonance spectroscopy and high-resolution mass spectrometry is reported for the first time for chemical characterization of substances from harvestmen, providing additional protocols for studies of scent secretions in the group.
Alkylated benzoquinones are produced by harvestmen of the suborder Laniatores, millipedes of the orders Spirobolida, Spirostreptida, and Julida, and insects of the orders Coleoptera, Dermaptera, Blattodea, Isoptera, Orthoptera, and Hemiptera (Eisner and Meinwald, 1966; Blum, 1981). The caustic, irritating, and odoriferous properties of these substances may cause invertebrate predators to desist from the attack and to perform vigorous cleaning activities (Eisner and Meinwald, 1966). Recently, Eisner et al. (2004) showed that the secretion of the gonyleptid harvestman A. aculeatus (Pachylinae) was an effective deterrent against the ant Formica exsectoides. Using a different protocol, we here demonstrate that secretions containing the 1,4-benzoquinones produced by the gonyleptid G. longipes (Goniosomatinae) effectively repel five species of ants belonging to three subfamilies. This result is expected because 1,4-benzoquinones from the secretion of several arthropods, including cockroaches, carabid, and tenebrionid beetles, have been shown to be highly repellent to ants (Eisner, 1958a,b; Peschke and Eisner, 1987).
As long as their defensive stores last, chemically defended arthropods can repel potential predators, but when exudates are depleted, they may quickly fall prey (Whitman et al., 1994). Many harvestmen, including representatives of the families Biantidae, Cosmetidae, Gonyleptidae, Manaosbiidae, Phalangodidae, and Sclerosomatidae, have mechanisms of emission that increase the evaporating surface of the defensive secretion (reviewed in Acosta et al., 1993; Hara and Gnaspini, 2003). These mechanisms are divided in two main types: (1) emission of secretion as fine spray that moistens the dorsum of the harvestman and (2) displacement of the secretion along the lateral area of the dorsal scutum. Both types of emission are supposed to create a “chemical shield” around the body of the harvestman protecting the animal after the emission of secretion (Acosta et al., 1993; Hara and Gnaspini, 2003). Our study demonstrates that secretions containing the 1,4-benzoquinones released by G. longipes in fact work as a “chemical shield” preventing the approach of three large predatory ants for at least 10 min. The “chemical shield” may protect the harvestman against the successive attacks of the same ant worker and also allow it to flee before massive ant recruitment. Because the families that have mechanisms to increase the evaporating surface of the defensive secretion are not closely related, it is possible that the “chemical shields” have evolved independently many times in the order Opiliones, and it may be related to the constant exposure to swarming insects, mainly ants.
The efficiency of the defensive secretion of harvestmen seems to vary among different spider families and genera. The ctenid Ctenus fasciatus is an important predator of gonyleptids in the wild (Gnaspini, 1996; Machado et al., 2000), and, therefore, it seems that the 1,4-benzoquinone secretions released by their prey have no effect on this spider. In captivity, individuals of the wolf spider Lycosa ceratiola (Lycosidae) were minimally affected by the effluent of the harvestman A. aculeatus, showing little response when the effluent was applied to their mouthparts as they fed on mealworms (Eisner et al., 2004). Using a similar protocol, we demonstrate here that secretions produced by G. longipes, whose composition is almost the same of A. aculeatus, are able to repel two large species of wandering spiders, E. cyclothorax (Ctenidae) and T. biocellata (Trechaleidae).
Contrary to what would be expected, Eisner et al. (2004) reported that live individuals of A. aculeatus were consistently rejected by spiders immediately after contact, before they were even prompted to release the defensive secretion. Three previous studies have reported that individuals of E. cyclothorax attack gonyleptid harvestmen but immediately retreat, avoiding biting the prey (Sabino and Gnaspini, 1999; Machado et al., 2000; Willemart and Kaneto, 2004). Eisner et al. (2004) discuss that A. aculeatus may contain additional chemical factors on the tegument that are repellent to spiders. It is also possible that such a repellent property derives from some residual action of the secretion, which may function as a “long persisting chemical shield”, and/or that harvestmen actually release small amounts of secretion, not able to be detected visually. Experimental specimens that are seized or pressed normally produce excessive amounts of fluids because of a highly stressful situation, but minimal amounts of fluids may be secreted in “slightly unpleasant” situations before an extremely shocking situation appears (Whitman, personal communication).
As far as we know, our study is the first to evaluate the defensive role of 1,4-benzoquinone secretions produced by a harvestman species against vertebrate predators. Given that the great majority of the neotropical Laniatores are nocturnal (Machado, personal observations), we focused on two groups of vertebrates that are active at night: frogs and marsupials. Both vertebrate predators used here occur syntopically with G. longipes and may include harvestmen in their diets, thus being appropriate model organisms for the bioassays. The defensive effluent of G. longipes stimulated a strong aversive reaction on the frogs, causing them to regurgitate the prey, which left the frogs' stomach still alive. Similar results were previously obtained in experiments using millipedes and beetles that have quinone-producing glands (see Eisner and Meinwald, 1966). As occurs with spiders, the effectiveness of the harvestmen secretion in anurans may vary among different species or genera. Especially in the genera Bufo (Bufonidae) and Rana (Ranidae), which are mainly constituted by large-bodied species, there are many records of predation on harvestmen (Bristowe, 1949; Smith and Bragg, 1949; Jenssen and Klimstra, 1966; Berry, 1970; Clarke, 1974; Acosta et al., 1995). Therefore, it is possible that the size of the frog influences the resistance to the secretion, so that smaller species are more likely to be affected than the larger ones. However, this suggestion remains to be tested.
Marsupials seem to be highly resistant to the chemical defenses of arthropods (Eisner, 1965, but see Whitman et al., 1994). Moreover, field data on the diet of some Brazilian didelphids have revealed that gonyleptid harvestmen are important food items for at least three species: Philander opossum, which enter caves to feed on resting individuals of G. spelaeum (Pelegatti-Franco and Gnaspini, 1996), D. albiventris, and Didelphis aurita, in which harvestmen are the most frequent prey (Cáceres and Monteiro-Filho, 2001; Cáceres, 2002). Indeed, the tested white eared-opossum D. albiventris consumed all crickets in both experimental groups, as well as the adult harvestmen offered in our experiment. However, individuals that bit the prey were clearly affected by the spray of secretion released by G. longipes. This spray reached the eyes and the snout of the opossums, making them immediately release the dead prey and spend nearly 2 min grooming their face. Similar results were obtained by Eisner (1965) who offered the walkingstick Anisomorpha (Phasmida) to the mouse opossum Micoureus demearare. Individuals of Anisomorpha also sprayed their defensive secretion on the face of the marsupial, and, despite the “obvious discomfort” caused, the opossum attacked and consumed all walkingsticks that were introduced into the cage.
Many gonyleptids show gregarious habits, and among the representatives of the subfamily Goniosomatinae, this behavior is especially common (Machado, 2002). Recently, it was demonstrated that the defensive secretion of Goniosoma aff. proximum elicits alarming behavior among aggregated individuals and that the reaction response to the signal is negatively correlated with the size of the group (Machado et al., 2002). The time elapsed between the presentation of the secretion and the dispersion of the individuals ranged from 2 to 52 sec. If after the first strike toward a harvestmen aggregation, a naive marsupial takes up to 120 sec to recover from the exposition to the secretion, there is enough time for all aggregated individuals to disperse. Because the aggregated individuals are not close relatives, there is no apparent advantage to the attacked harvestman, which dies after the first strike of the marsupial. However, the emission of defensive secretion by one individual may retard the attack of the marsupial and enhance the escape capability of conspecifics in an aggregation of harvestmen (see Discussion in Machado, 2002). Gregariousness, therefore, may be a defensive mechanism especially important against mammal predators because harvestman secretions seem not to be so effective.
The results of our study indicate that the 1,4-benzoquinones released by G. longipes are an effective defense against predation, and that the effectiveness of the secretion depends on the predator type. The emission of scent gland secretions repelled seven species of ants, two species of wandering spiders, and one species of frog, but it was not an effective defense against an opossum. Our data support the suggestion that chemical defenses may increase survival with some but not all potential predators (e.g., Krall et al., 1999; Staples et al., 2002; Eisner et al., 2004). This variation in defense effectiveness may result from many interacting factors, including, on one hand, the attack strategy, size, learning ability, and physiology of the predators and, on the other hand, the chemical nature of the defensive compounds, type of emission, and amount of effluent released by the prey (Whitman et al., 1994). In this way, harvestmen are good model organisms because they release a great variety of compounds and are attacked by numerous potential and a few actual predators.
References
L. E. Acosta T. I. Poretti P. E. Mascarelli (1993) ArticleTitleThe defensive secretions of Pachyloidellus goliath (Opiliones: Laniatores: Gonyleptidae) Bonn. Zool. Beitr. 44 19–31
L. E. Acosta F. E. Pereyra R. A. Pizzi (1995) ArticleTitleField observations on Pachyloidellus goliath (Opiliones, Gonyleptidae) in Pampa de Achala, province of Córdoba, Argentina Bull. Br. Arachnol. Soc. 10 23–28
P. Y. Berry (1970) ArticleTitleThe food of the giant toad Bufo asper Zool. J. Linn. Soc. 49 61–68
M. S. Blum (1981) Chemical Defenses of Arthropods Academic Press New York
M. S. Blum A. L. Edgar (1981) ArticleTitle4-Methyl-3-heptanone—identification and role in opilionid exocrine secretions Insect Biochem. 1 181–188 Occurrence Handle10.1016/0020-1790(71)90071-0
W. S. Bristowe (1949) ArticleTitleThe distribution of harvestmen (Phalangida) in Great Britain and Ireland, with notes on their names, enemies and food J. Anim. Ecol. 18 100–114
H. Budzikiewicz C. Djerassi D. H. Williams (1967) Mass Spectrometry of Organic Compounds Holden-Day Inc. San Francisco
N. C. Cáceres (2002) ArticleTitleFood habits and seed dispersal by the white-eared opossum, Didelphis albiventris, in Southern Brazil Stud. Neotrop. Fauna Environ. 37 97–104 Occurrence Handle10.1076/snfe.37.2.97.8582
N. C. Cáceres E. L. A. Monteiro-Filho (2001) ArticleTitleFood habits, home range and activity of Didelphis aurita (Mammalia, Marsupialia) in a forest fragment of Southern Brazil Stud. Neotrop. Fauna Environ. 36 85–92 Occurrence Handle10.1076/snfe.36.2.85.2138
R. D. Clarke (1974) ArticleTitleFood habitats of toads, genus Bufo (Amphibia: Bufonidae) Am. Midl. Nat. 91 140–147
J. L. Cloudsley-Thompson (1958) Spiders, Scorpions, Centipedes and Mites Pergamon Press London
A. L. Edgar (1971) ArticleTitleStudies on the biology and ecology of Michigan Phalangida (Opiliones) Misc. Pub. Mus. Zool., Univ. Mich. 144 1–64
T. Eisner (1958) ArticleTitleThe protective role of the spray mechanism of the bombardier beetle, Brachynus ballistarius Lec J. Insect Physiol. 2 215–220 Occurrence Handle10.1016/0022-1910(58)90006-4
T. Eisner (1958) ArticleTitleSpray mechanism of the cockroach Diploptera punctata Science 128 148–149
T. Eisner (1965) ArticleTitleDefensive spray of a phasmid insect Science 148 966–968
T. Eisner J. Meinwald (1966) ArticleTitleDefensive secretions of arthropods Science 153 1341–1350
T. Eisner A. F. Kluge J. E. Carrel J. Meinwald (1971) ArticleTitleDefense of phalangid: liquid repellent administered by leg dabbing Science 173 650–652
T. Eisner T. H. Jones K. Hicks R. E. Silberglied J. Meinwald (1977) ArticleTitleQuinones and phenols in the defensive secretions of neotropical opilionids J. Chem. Ecol. 3 321–329 Occurrence Handle10.1007/BF00988447
T. Eisner D. Alsop J. Meinwald (1978) Secretions of opilionids, whip scorpions and pseudoscorpions S. Bettini (Eds) Handbook of Experimental Pharmacology (Arthropod Venoms), Vol. 48 Springer-Verlag Berlin 87–99
T. Eisner R. C. Morgan A. B. Attygalle S. R. Smedley K. B. Herath J. Meinwald (1997) ArticleTitleDefensive production of quinoline by a phasmid insect (Oreophoetes peruana) J. Exp. Biol. 200 2493–2500 Occurrence Handle9366083
T. Eisner C. Rossini A. González M. Eisner (2004) ArticleTitleChemical defense of an opilionid (Acanthopachylus aculeatus) J. Exp. Biol. 207 1313–1321 Occurrence Handle10.1242/jeb.00849 Occurrence Handle15010482
C. Estable M. I. Ardao N. P. Brasil L. F. Fieser (1955) ArticleTitleGonyleptidine J. Am. Chem. Soc. 77 4942 Occurrence Handle10.1021/ja01623a087
L. F. Fieser M. I. Ardao (1956) ArticleTitleInvestigation of the chemical nature of Gonyleptidine J. Chem. Ecol. 78 774–781
A. A. Giaretta M. S. Araújo H. F. Medeiros K. G. Facure (1998) ArticleTitleFood habits and ontogenetic shifts of the litter dwelling frog Proceratophrys boiei (Wied) Rev. Bras. Zool. 15 385–388
P. Gnasnipi (1996) ArticleTitlePopulation ecology of Goniosoma spelaeum, a cavernicolous harvestman from Southeastern Brazil (Arachnida: Opiliones: Gonyleptidae) J. Zool. 239 417–435
P. Gnaspini A. J. Cavalheiro (1998) ArticleTitleChemical and behavioral defenses of a Neotropical cavernicolous harvestman: Goniosoma spelaeum (Opiliones: Laniatores: Gonyleptidae) J. Arachnol. 26 81–90
M. R. Hara P. Gnaspini (2003) ArticleTitleComparative study of the defensive behavior and morphology of the gland opening area among harvestmen (Arachnida, Opiliones, Gonyleptidae) under a phylogenetic perspective Arthropod Struct. Develop. 32 257–275 Occurrence Handle10.1016/S1467-8039(03)00040-9
R. G. Holmberg (1986) ArticleTitleThe scent glands of Opiliones: a review of their function Proc. 5th Cong. Int. Arachnol. 1983 131–133
V. Immel (1954) ArticleTitleZur Biologie und Phisiologie von Nemastoma quadripunctatum (Opiliones: Dyspnoi) Zool. Jahrb. (Syst.) 83 129–184
T. A. Jenssen W. D. Klimstra (1966) ArticleTitleFood habits of the green frog, Rana clamitans, in southern Illinois Am. Midl. Nat. 76 169–182
C. Juberthie (1976) ArticleTitleChemical defense in soil opiliones Rev. Ecol. Biol. Sol 13 155–160
B. S. Krall R. J. Bartelt C. J. Lewis D. W. Whitman (1999) ArticleTitleChemical defense in the stink bug Cosmopepla bimaculata J. Chem. Ecol. 25 2477–2494 Occurrence Handle10.1023/A:1020822107806
G. Machado (2002) ArticleTitleSubsociality, gregariousness, and defensive behavior in neotropical Goniosoma harvestmen (Arachnida: Opiliones) Insectes Soc. 49 388–393
G. Machado P. S. Oliveira (1998) ArticleTitleReproductive biology of the Neotropical harvestman Goniosoma longipes (Arachnida: Opiliones: Gonyleptidae): mating and oviposition behaviour, brood mortality, and parental care J. Zool. 246 359–367 Occurrence Handle10.1017/S0952836998009881
G. Machado R. L. G. Raimundo P. S. Oliveira (2000) ArticleTitleDaily activity schedule, gregariousness, and defensive behaviour in the Neotropical harvestman Goniosoma longipes (Arachnida: Opiliones: Gonyleptidae) J. Nat. Hist. 34 587–596 Occurrence Handle10.1080/002229300299453
G. Machado V. Bonato P. S. Oliveira (2002) ArticleTitleAlarm communication: a new function for the scent gland secretion in harvestmen (Arachnida: Opiliones) Naturwissenschaften 89 357–360 Occurrence Handle10.1007/s00114-002-0337-8 Occurrence Handle12435036
R. R. Pagano (1994) Understanding Statistics in the Behavioral Sciences West Publishing Company Minneapolis, USA
F. P. Pelegatti-Franco P. Gnaspini (1996) ArticleTitleUse of caves by Philander opossum (Mammalia: Didelphidae) in Southeastern Brazil Pap. Avulsos Zool. 39 351–364
K. Peschke T. Eisner (1987) ArticleTitleDefensive secretion of a beetle (Blaps mucronata): physical and chemical determinants of effectiveness J. Comp. Physiol., A 161 377–388
B. Roach T. Eisner J. Meinwald (1980) ArticleTitleDefensive substances of opilionids J. Chem. Ecol. 6 511–516 Occurrence Handle10.1007/BF01402927
J. Sabino P. Gnaspini (1999) ArticleTitleHarvestman (Opiliones: Gonyleptidae) takes prey from a spider (Araneae: Ctenidae) J. Arachnol. 27 675–678
J. W. Shultz (1990) ArticleTitleEvolutionary morphology and phylogeny of Arachnida Cladistics 6 1–38
C. C. Smith A. N. Bragg (1949) ArticleTitleObservations on the ecology and natural history of Anura, VII. Food and feeding habits of the common species of toads in Oklahoma Ecology 30 333–349
J. K. Staples B. S. Krall R. J. Bartelt D. W. Whitman (2002) ArticleTitleChemical defense in the plant bug Lopidea robiniae J. Chem. Ecol. 28 601–615 Occurrence Handle10.1023/A:1014552414580 Occurrence Handle11944836
R. L. Teixeira E. S. Coutinho (2002) ArticleTitleHábito alimentar de Proceratophrys boiei (Wied) (Amphibia, Anura, Leptodactylidae) em Santa Teresa, Espírito Santo, sudeste do Brasil Bol. Mus. Biol. Mello Leitão 14 13–20
D. W. Whitman M. S. Blum D. W. Alsop (1994) Allomones: chemicals for defense D. L. Evans J. O. Schmidt (Eds) Insect Defenses: Adaptative Mechanisms and Strategies of Prey and Predators State University of New York Albany 289–351
R. H. Willemart G. E. Kaneto (2004) ArticleTitleOn the natural history of the Neotropical spider Enoploctenus cyclothorax (Araneae, Ctenidae) Bull. Br. Arachnol. Soc. 13 53–59
Acknowledgments
We are grateful to our friends A.C. Pellegrino, B.A. Buzatto, E.G. Martins, F. Osses, G.B.T. Egito, G.S. Requena, J.M. Fanton, L.M. Rossetto, S. Koehler, and T.M. Del Corso for helping during this study, to B.A. Buzatto for the photos of the harvestman, to E.G. Martins, I. Sazima, and R.J. Sawaya for the photos of the predators, to the staff of the Clube Náutico Araraquara for logistical support, to A.J. Santos (spiders), R. Cogni (ants), and E.G. Martins (marsupials) for taxonomic identifications, and to B.A. Buzatto, G.S. Requena, I. Sazima, P. Gnaspini, D.W. Whitman, and an anonymous reviewer for comments on an early draft of the manuscript. This study was supported by fellowships from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, no. 02/00381-0 and no. 03/09357-7).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Machado, G., Carrera, P.C., Pomini, A.M. et al. Chemical Defense in Harvestmen (Arachnida, Opiliones): Do Benzoquinone Secretions Deter Invertebrate and Vertebrate Predators?. J Chem Ecol 31, 2519–2539 (2005). https://doi.org/10.1007/s10886-005-7611-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-005-7611-0