
Abstract
Celiac disease (CD) is a chronic immune-mediated intestinal inflammatory disorder afflicting genetically susceptible individuals triggered by the consumption of dietary cereals with high gluten content. As with many other organ-specific autoimmune diseases, the dominant tissue-destructive inflammation in CD is T cell-mediated. The proinflammatory cytokine IL-15 that is overexpressed in the intestinal epithelium of CD patients has emerged as a pivotal element that orchestrates intestinal inflammation and T cell-mediated autoimmune tissue destruction. Although no animal model exists that recapitulates the full spectrum of CD pathophysiology, we have previously reported that transgenic mice that overexpress human IL-15 in enterocytes (T3b-hlL-15 Tg) display many of the T cell-mediated pathologic features seen in CD. Extending these observations, we now report that T3b-hlL-15 Tg mice in addition to recapitulating T cell-mediated effects also display autoantibodies including those against tissue transglutaminase 2 and extensive lamina propria plasmacytosis, all of which are characteristic of CD, thereby reflecting the possibility that locally expressed IL-15 drives both T and B cell pathologic effects seen in CD. More importantly, these findings support the validity and utility of T3b-hlL-15 Tg mice as a reasonable model to investigate not only tissue-destructive pathologic processes in CD, but also to explore novel therapeutic modalities for the treatment of this disease.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Celiac disease (CD) is a chronic intestinal inflammatory disease that develops in genetically susceptible individuals bearing the HLA class II DQ alleles that encode DQ2 or DQ8 molecules triggered by the exposure to dietary cereals with gluten proteins [1]. The genetic and environmental/dietary elements that precipitate CD have been extensively characterized [2]. As with several other organ-specific immune-mediated diseases, CD is primarily a T cell-mediated immune disorder where gluten-specific CD4+ T cells that recognize gluten peptide in the context of HLA class II molecules initiate the pathogenic cascade. However, these gluten-specific CD4+ T cells are not direct mediators of the extensive intestinal tissue damage seen in CD, but it is massively infiltrating CD8+ intraepithelial lymphocytes (IEL), a hallmark of CD, that cause extensive tissue damage [3]. These infiltrating CD8+ lymphocytes, unlike the proliferating lamina propria CD4+ T cells that display exquisite gluten specificity, lack demonstrable gluten specificity and appear to cause extensive enterocyte and underlying tissue damage, via T cell receptor (TCR)-independent mechanisms with the involvement of NKG2D and other coactivator NK cell receptors [4]. In addition to these prominent T cell-mediated features, the appearance of antiendomysial antibodies, one of the earliest predictors that presage clinical disease as well as the presence of massive plasmacytosis with a preponderance of IgA-secreting plasma cells in the laminar propria, attests to B cell elements associated with CD [5].
The proinflammatory cytokine IL-15 has emerged as a central driver that orchestrates T cell-mediated tissue damage in CD. It is essential for the generation of IEL and sustains these cells perhaps by protecting them from activation-induced cell death [6]. IL-15 promotes CD8+ T cell proliferation and enhances their effector cytolytic activities in addition to reprogramming IEL to lymphokine-activated killer cells or NK-like cells that undergo oligoclonal expansion to perpetuate a massive population of CD8+ T cells that target enterocytes independent of TCR engagement in a process that is further facilitated by the induction of the NKG2D pathway [7]. In completing this cycle of sustained tissue damage, IL-15 also induces cognate receptors of NKG2D such as MICA/B, ULBPs, and HLA-E on enterocytes [reviewed in refs. 2, 4]. In addition, IL-15 in concert with retinoic acid has been shown to disrupt immune homeostasis in the gut, resulting in breaching of immune tolerance to dietary antigens [8]. Finally, IL-15 also disables operational anti-inflammatory checkpoints in the intestinal mucosa by inhibiting Smad-dependent TGF-β signaling thereby further facilitating intestinal tissue destruction in CD patients [9].
Despite extensive gains in understanding the pathophysiology of CD, much remains to be explored and the absence of a small animal model that recapitulates the full spectrum of human disease has not only been a major obstacle in this regard, but has also seriously impacted the exploration of new therapeutic modalities for CD for which the only treatment available now is a lifelong gluten-free diet. Recently, we reported that transgenic mice that express human IL-15 in enterocytes from an enterocyte-specific T3b promoter (T3b-hlL-15 Tg mice) manifest almost all salient intestinal pathologic features of CD including anatomically demarcated duodenojejunal inflammation with massively infiltrated NKG2D-expressing CD8+ IEL along with severe villous atrophy and crypt hyperplasia [10]. The blockade of locally expressed IL-15 in these T3b-hIL-15 Tg mice resulted in complete resolution of the intestinal pathology thus strengthening the notion that locally expressed IL-15 is causally linked to T cell-mediated intestinal tissue damage in CD [10]. With further evaluation of T3b-hIL-15 Tg mice as a relevant animal model that closely resembles the intestinal pathology of CD, we now report that T3b-hIL-15 Tg mice in addition to recapitulating T cell-mediated effects also display autoantibodies including antibodies against tissue transglutaminase 2 (TG2) and extensive lamina propria plasmacytosis, features that are characteristic of CD, thereby raising the possibility that locally expressed IL-15 drives both T and B cell pathologic effects seen in CD. More importantly, these findings support the validity and utility of T3b-hIL-15 Tg mice as a reasonable model to investigate not only tissue-destructive pathologic processes in CD, but also to explore novel therapeutic modalities for the treatment of this disease, especially those that target IL-15.
Materials and Methods
Mice
The generation of T3b-hIL-15 Tg mice has been reported previously [11]. All of the animal experiment protocols were approved by the Tokyo Metropolitan Institute of Medical Science Animal Care and Use Committee. T3b-hIL-15 Tg mice that were over 8 months of age were used for the experiments.
Cell Preparations
Peripheral blood mononuclear cells and splenocytes were prepared as described previously [11]. Isolation of lamina propria lymphocytes in the small intestine (SI-LPL) by Percoll gradient centrifugation has been described elsewhere [11].
ELISA
Total IgM, IgG, and IgA levels in the sera of T3b-hIL-15 Tg mice and their wild-type (WT) littermates were measured by a sandwich ELISA as described previously [12]. The detection of anti-TG2 antibodies and rheumatoid factor (RF) were performed as described [13, 14]. The level of anti-dsDNA antibodies was measured using a commercially available ELISA kit (Shibayagi, Japan) according to the manufacturer’s instructions.
A Flow Cytometry Analysis
The frequency of lymphocytes was analyzed by flow cytometry using standard protocols. Briefly, cells were stained with FACS buffer containing 10% FBS and 2 mM EDTA. Next, Fc receptors of lymphocytes were blocked with anti-CD16/32 antibody for 20 min at 4°C, and lymphocytes were then stained with combinations of fluorochrome-conjugated antibodies to CD3E-PE-Cy7, CD138-PE, CD19-FITC (BD Pharmingen, USA), and B220-eFluor450 (e-Bioscience, USA) for 20 min at 4°C. The stained lymphocytes were then analyzed with a FACS CANTO II flow cytometer (BD Biosciences, USA) using the FlowJo software (Tree Star Inc., USA). The number of B cells, plasmablasts, and plasma cells were quantitated by counting the number of CD3ε− CD19+ B220+ cells (for B cells), CD3ε− B220+ CD138+ cells (for plasmablasts), and CD3ε− B220− CD138+ cells (for plasma cells) present in one million cells from the spleen and lamina propria of the small intestines isolated from each mouse, respectively, and these numbers were then used to calculate the absolute number of B cells, plasmablasts, and plasma cells for that mouse based on the total number of cells isolated from the spleen and lamina propria of the mouse.
Tissue Staining and Histology
Immunostaining of OCT-mounted fresh tissues was performed as previously described [15]. For the staining of B cells, biotinylated anti-B220 and streptavidin–PE antibodies and anti-CD8α-FITC antibody to stain T cells (BD Biosciences, USA) were used. For the detection of binding of autoantibodies, 1:4 diluted serum samples from WT mice and T3b-hIL-15 Tg mice were used as primary antibodies, and a 1:1,000 diluted FITC-conjugated goat antimouse IgG antibody (Jackson Immunoreseach laboratories, USA) was used as a secondary antibody.
Results
Elevated Autoantibody Levels Are Observed in the Sera of T3b-hIL-15 Tg mice
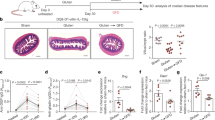
Because elevated serum immunoglobulin levels are a consistent finding in CD patients, we measured the serum immunoglobulin levels in the T3b-hIL-15 Tg mice. Elevated levels of total IgM and IgG were observed in the sera of T3b-hIL-15 Tg mice in comparison to their littermate WT controls, and these differences were statistically significant (Fig. 1a, b). It should be noted that, when we assessed the serum IgA levels, most T3b-hIL-15 Tg mice displayed relatively higher levels of serum IgA in comparison to WT controls, but these differences did not achieve statistical significance (Fig. 1c). Because the presence of autoantibodies is observed in CD against certain self-antigens, for example, tissue TGs [16], dsDNA [17], RF [18], actin [19], desmin [20], etc., we examined whether T3b-hIL-15 Tg mice display any autoantibodies against these self-antigens in their sera. Multiple autoantibodies were detected in T3b-hIL-15 Tg mice including those of anti-dsDNA antibody, RF, and anti-TG2 antibody, again drawing close parallels with those auto antibodies seen in CD (Fig. 1d–f). Although there were measurable levels of elevated antibodies against the cytoskeletal components actin and desmin, the levels of these antibodies were not statistically significant when compared to the levels observed in WT littermates (data not shown).

Sera of T3b-hIL-15 Tg mice display elevated levels of immunoglobulins and the presence of autoantibodies. The levels of serum IgM (a), IgG (b), IgA (c), anti-dsDNA (d), RF (e) and anti-TG2 (f) were measured by an ELISA
Accumulation of B Cells in the Small Intestinal Lamina Propria Displaying Inflammatory Lesions in T3b-hIL-15 Tg Mice
Another consistent finding in the affected intestinal mucosa of CD patients is the accumulation of B cells including differentiated plasma cells in the lamina propria, although what drives this plasmacytosis of small intestinal lamina propria remains unclear [5, 21, 22]. Having demonstrated that T3b-hIL-15 Tg mice have higher levels of serum immunoglobulins and multiple species of autoantibodies, we examined whether there were any alterations in the B cell profiles of these mice both in the periphery and in the affected intestinal submucosa initially by flow cytometry. The total numbers of lymphocytes isolated from the spleen and the lamina propria of T3b-hlL-15 Tg mice were 88-fold and 24-fold higher, respectively, than the total numbers of lymphocytes isolated from the WT littermates. As shown in Fig. 2a, the absolute numbers of B cells (CD3ε− B220+ CD19+) in the spleen and small intestinal lamina propria of the T3b-hIL-15 Tg mice were significantly higher in comparison to their WT littermates. Furthermore, in order to determine the frequency of antibody-secreting cells, we investigated the expression of CD138 (Syndecan-1) as a cell surface maker of plasma cells on the splenocytes and small intestinal lamina propria cell in T3b-hlL-15 Tg mice. Fully differentiated immunoglobulin-secreting plasma cells, the cell population with CD3ε− B220− CD138+ phenotype, and plasmablasts, which are the precursor cells of plasma cells with CD3ε− B220+ CD138+ phenotype, were identified by flow cytometry, and the absolute numbers of plasma cells and plasmablasts were calculated. The numbers of plasma cells and plasmablasts in the affected lamina propria of T3b-hIL-15 Tg mice displayed a tenfold and a fivefold increase, respectively, over their WT littermates (Fig. 2). However, the absolute number of plasmablasts in the spleens of T3b-hTL-15 Tg mice was approximately twofold less in comparison to WT littermates. As has been observed in CD patients, a reduction in the number of plasma cells in the periphery and spleen may be a reflection of compartmentalization of these plasma cells in the lamina propria that display florid inflammation in T3b-hIL-15 Tg mice. Confirming the increased B cell accumulation in the lamina propria of T3b-hIL-15 Tg mice as determined by FACS analyses (Fig. 2), when we examined the accumulation of B lymphocytes in the lamina propria by immunostaining of the proximal small intestinal sections with a B220-specific antibody, there was abundant B220-specific fluorescence in the lamina propria of the intestinal sections of T3b-hIL-15 Tg mice, whereas the sections derived from WT littermates displayed much reduced fluorescence (Fig. 3). It should be noted that, in addition to readily demonstrable plasmacytosis, as we have previously shown, there was massive infiltration of CD8+ T cells into the small intestinal mucosa that displayed extensive villous atrophy in these T3b-hIL-15 Tg mice as well (Fig. 3).

Detection of increased B lymphocytes in the spleen and small intestinal lamina propria of T3b-hlL-15 Tg mice by flow cytometry. Splenocytes and SI-LPL were isolated from four individual mice from each group and analyzed by flow cytometry for quantitation of B cells (CD3ε− CD19+ B220+), plasmablasts (CD3ε− B220+ CD138+), and plasma cells (CD3ε− B220− CD138+). For flow cytometric analysis, 1 × 106 cells were used and the number of B cells, plasmablasts, and plasma cells present were determined, and then from these numbers, the absolute numbers of B (left panel), plasmablasts (center panel), and plasma cells (right panel) that were present in the spleens and small intestinal lamina propria of T3b-hIL-15 Tg mice and their WT littermates were calculated

Enhanced accumulation of CD8+ T cells and B cells in the proximal small intestines of T3b-hIL-15 Tg mice as detected by immunohistochemistry. Immunostaining was performed on sections from the duodenojejunal region of the small intestines with FITC-conjugated CD8α antibody and biotinylated B220 antibody. PE-conjugated streptavidin was used to detect bound biotin. The data shown are representative of three independent experiments
Small Intestinal Tissue Reactive Autoantibodies Are Present in the Sera of T3b-hIL-15 Tg Mice
The appearance of autoantibodies including those that react with endomysium and tissue TG is a consistent early manifestation of CD that presages the onset of clinical disease, and in fact, these antibodies are utilized in the serologic testing and identification of individuals prone to develop clinical disease. However, any role for these autoantibodies in the pathologic tissue-destructive process remains unproven largely due to the unavailability of suitable model systems to study the effects of these autoantibodies in the pathologic process. In an attempt to address whether autoantibodies present in the sera of T3b-hIL-15 Tg mice could contribute to the tissue-destructive process, we examined whether the sera of T3b-hIL-15 Tg mice had any reactivity against small intestinal tissues. As shown in Fig. 4, the sera from T3b-hIL-l5 Tg mice displayed reactivity against intestinal tissues of normal mice, whereas the sera from WT littermates failed to elicit any appreciable reactivity against intestinal tissues.

Autoantibodies that react with small intestinal tissues are detected in sera of T3b-hIL-15 Tg mice. Sera obtained from 8-month-old WT mice or T3b-hIL-15 Tg mice were incubated with proximal small intestinal tissue sections obtained from 3-month-old WT mice. The bound mouse antibodies were detected using an FITC-labeled antimouse IgG as the secondary antibody. The data shown are representative of three independent experiments
Discussion
In our continuing pursuit of developing a mouse model that recapitulates human CD, we have focused on T3b-hIL-15 Tg mice that express human IL-15 locally in the intestinal epithelium due to the usage of the enterocyte-specific T3b promoter for the expression of the IL-15 transgene [11]. Although as expected these mice do not show gluten sensitivity, the locally expressed IL-15, as we have shown previously, singularly drives a spontaneous intestinal inflammatory pathology that bears striking resemblance to both macroscopic and microscopic pathologic lesions seen in human CD with 100% penetrance [10, 11]. Adult T3b-hIL-15 Tg mice manifest florid inflammatory lesions that are exclusively confined to the proximal duodenojejunal region of the small intestine with extensive villous atrophy, some degree of crypt hyperplasia, and massive accumulation of NK-like CD8+ T lymphocytes bearing NKG2D receptors, all of which are pathognomonic in human CD. Unlike the T3b-hIL-15 Tg mouse model, the double transgenic human CD4/HLA-DR3-DQ2 [13] or HCD4/DQ8 [23] mice do not display autoantibody production or enteropathy, although the HCD4/DQ8 in autoimmune-prone NOD background has been shown to develop cutaneous lesions resembling dermatitis herpetiformis seen in some CD patients upon immunization with gluten plus pertussis toxin [24]. More recently, a double transgenic mouse created by crossing humanized HLA-DQ8 with an IL-15 transgenic mouse that overexpresses mouse IL-15 from a ubiquitous MHC class I promoter but without any induced or spontaneous enteropathy has been shown to develop autoantibodies against gliadin and tissue TG2, along with intraepithelial lymphocytosis that mimic early stages of CD upon feeding gliadin. However, even in these mice, although IFN-γ-producing antigliadin T cells are readily demonstrable following the feeding of gliadin, the disease-defining villous atrophy is absent [8].
Many elegant studies have contributed to the prevailing consensus that locally hyperexpressed IL-15 is centrally involved in orchestrating the T cell-mediated inflammatory intestinal tissue damage [reviewed in refs. 1, 2, 4]. However, the formal proof of this view that IL-15 is pivotally causal rather than a mere correlate awaits the direct demonstration in a clinical trial involving CD patients that the blockade of IL-15 activity leads to amelioration or reversal of pathologic inflammatory lesions. Nonetheless, in support of this notion, we recently demonstrated that the infusion of a monoclonal antibody directed against CD122, a receptor that is critical in IL-15 signaling, results in the complete reversal of pathologic inflammatory lesions in T3b-hIL-15 Tg mice [10].
Despite T cell-mediated effects being primarily responsible for the pathologic lesions in CD, the B cell-mediated effects such as the appearance of autoantibodies including those against tissue TGs constitute the earliest signs of impending clinical disease [25]. However, the impact of these B cell effects in the pathogenesis of CD or the factors that trigger these B cell effects that presage clinical disease remain largely undefined. In this study, we have demonstrated that the overexpression of IL-15 in epithelial cells of the small intestine can induce the production of autoantibodies, some of which can react with the small intestinal tissues. We have previously reported that the administration of an anti-CD122 monoclonal antibody that ablates IL-15-dependent NK-like intraepithelial CD8+ T cells results in the complete resolution of inflammatory lesions in the small intestines [10]. Nonetheless, to elucidate whether these IL-15-induced autoantibodies are also involved in the pathogenesis of intestinal inflammatory lesions of T3b-hIL-15 Tg mice, we are currently crossing the μ chain-deficient mice which cannot produce any antibodies [26] with our T3b-hIL-15 Tg mice. It is also important to emphasize that, in these T3b-hIL-15 Tg mice, despite having elevated levels of serum immunoglobulins and autoantibodies, we have failed to detect any extraintestinal immune pathologic lesions including in the kidneys, lungs, or heart.
As we have shown in this study, the presence of autoantibodies including those against tissue TG2 (Fig. 1f) and the accumulation of plasmacytes in the lamina propria underlying the intestinal inflammatory lesions in the proximal duodenojejunal region (Figs. 2 and 3), along with the elevated levels of serum immunoglobulin (Fig. 1), not only attest to close recapitulation of prominent B cell-mediated effects seen in CD in these T3b-hIL-15 Tg mice, but also indicate that the locally expressed IL-15 in the small intestines is sufficient and capable of eliciting both T and B cell-mediated pathologic manifestations that define human CD.
Conclusions
While it is well established that the pleiotropic cytokine IL-15 has profound effects on both T and B lymphocyte functions and biology [27–29], it would have been difficult to predict that the local overexpression of this cytokine alone could bring about the full spectrum of pathologic manifestations seen in CD. More importantly, the availability of the T3b-hlL-15 Tg mouse model would now not only allow us to dissect out as to how locally expressed IL-15 in the intestine could orchestrate T and B cell pathologic events, thus providing valuable insights into the pathologic process of CD, but these mice would also serve as a reliable model system to explore novel therapeutic modalities in the treatment of this debilitating autoimmune disease that has a propensity to give rise to malignancies and considerably lowers the life expectancy of affected individuals.
Abbreviations
- CD:
-
Celiac disease
- TG2:
-
Transglutaminase 2
- IL-15:
-
Interleukin 15
- RF:
-
Rheumatoid factor
References
Kagnoff MF. Celiac disease: pathogenesis of a model immunogenetic disease. J Clin Invest. 2007;117(1):41–9.
Abadie V, Sollid LM, Barreiro LB, Jabri B. Integration of genetic and immunological insights into a model of celiac disease pathogenesis. Annu Rev Immunol. 2010;29:493–525.
Marsh MN. Gluten, major histocompatibility complex, and the small intestine. A molecular and immunobiologic approach to the spectrum of gluten sensitivity (‘celiac sprue’). Gastroenterology. 1992;102(1):330–54.
Hue S, Mention JJ, Monteiro RC, Zhang S, Cellier C, Schmitz J, et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity. 2004;21(3):367–77.
Scott BB, Goodall A, Stephenson P, Jenkins D. Small intestinal plasma cells in coeliac disease. Gut. 1984;25(1):41–6.
Malamut G, El Machhour R, Montcuquet N, Martin-Lanneree S, Dusanter-Fourt I, Verkarre V, et al. IL-15 triggers an antiapoptotic pathway in human intraepithelial lymphocytes that is a potential new target in celiac disease-associated inflammation and lymphomagenesis. J Clin Invest. 2010;120(6):2131–43.
Meresse B, Chen Z, Ciszewski C, Tretiakova M, Bhagat G, Krausz TN, et al. Coordinated induction by IL.15 of a TCR-independent NKG2D signaling pathway converts CTL into lymphokine-activated killer cells in celiac disease. Immunity. 2004;21(3):357–66.
DePaolo RW, Abadie V, Tang F, Fehlner-Peach H, Hall JA, Wang W, et al. Co-adjuvant effects of retinoic acid and IL-15 induce inflammatory immunity to dietary antigens. Nature. 2011;471(7337):220–4.
Benahmed M, Meresse B, Arnulf B, Barbe U, Mention JJ, Verkarre V, et al. Inhibition of TGF-beta signaling by IL-15: a new role for IL-15 in the loss of immune homeostasis in celiac disease. Gastroenterology. 2007;132(3):994–1008.
Yokoyama S, Watanabe N, Sato N, Perera PY, Filkoski L, Tanaka T, et al. Antibody-mediated blockade of IL-15 reverses the autoimmune intestinal damage in transgenic mice that overexpress IL-15 in enterocytes. Proc Natl Acad Sci USA. 2009;106(37):15849–54.
Ohta N, Hiroi T, Kweon MN, Kinoshita N, Jang MH, Mashimo T, et al. IL-15-dependent activation-induced cell death-resistant Th1 type CD8 alpha beta+NK1.1+ T cells for the development of small intestinal inflammation. J Immunol. 2002;169(1):460–8.
Kweon MN, Yamamoto M, Kajiki M, Takahashi I, Kiyono H. Systemically derived large intestinal CD4(+) Th2 cells play a central role in STAT6-mediated allergic diarrhea. J Clin Invest. 2000;106(2):199–206.
de Kauwe AL, Chen Z, Anderson RP, Keech CL, Price JD, Wijburg O, et al. Resistance to celiac disease in humanized HLA-DR3-DQ2-transgenic mice expressing specific anti-gliadin CD4+ T cells. J Immunol. 2009;182(12):7440–50.
Bobe P, Bonardelle D, Benihoud K, Opolon P, Chelbi-Alix MK. Arsenic trioxide: a promising novel therapeutic agent for lymphoproliferative and autoimmune syndromes in MRL/lpr mice. Blood. 2006;108(13):3967–75.
Jang MH, Sougawa N, Tanaka T, Hirata T, Hiroi T, Tohya K, et al. CCR7 is critically important for migration of dendritic cells in intestinal lamina propria to mesenteric lymph nodes. J Immunol. 2006;176(2):803–10.
Dieterich W, Ehnis T, Bauer M, Donner P, Volta U, Riecken EO, et al. Identification of tissue transglutaminase as the autoantigen of celiac disease. Nat Med. 1997;3(7):797–801.
Lerner A, Blank M, Lahat N, Shoenfeld Y. Increased prevalence of autoantibodies in celiac disease. Dig Dis Sci. 1998;43(4):723–6.
Hällgren J, Knutson F, Lavö B, Hällgren R. Increased mucosal synthesis of rheumatoid factor (RF) in coeliac disease. Clin Exp Immunol. 1996;103(1):94–8.
Granito A, Muratori P, Cassani F, Pappas G, Muratori L, Agostinelli D, et al. Anti-actin IgA antibodies in severe coeliac disease. Clin Exp Immunol. 2004;137(2):386–92.
Teesalu K, Uibo O, Kalkkinen N, Janmey P, Uibo R. Increased levels of IgA antibodies against desmin in children with coeliac disease. Int Arch Allergy Immunol. 2001;126(2):157–66.
Wood GM, Howdle PD, Trejdosiewicz LK, Losowsky MS. Jejunal plasma cells and in vitro immunoglobulin production in adult coeliac disease. Clin Exp Immunol. 1987;69(1):123–32.
Brandtzaeg P. The changing immunological paradigm in coeliac disease. Immunol Lett. 2006;105(2):127–39.
Black KE, Murray JA, David CS. HLA-DQ determines the response to exogenous wheat proteins: a model of gluten sensitivity in transgenic knockout mice. J Immunol. 2002;169(10):5595–600.
Marietta E, Black K, Camilleri M, Krause P, Rogers 3rd RS, David C, et al. A new model for dermatitis herpetiformis that uses HLA-DQ8 transgenic NOD mice. J Clin Invest. 2004;114(8):1090–7.
Salmi TT, Collin P, Jarvinen O, Haimila K, Partanen J, Laurila K, et al. Immunoglobulin A autoantibodies against transglutaminase 2 in the small intestinal mucosa predict forthcoming coeliac disease. Aliment Pharmacol Ther. 2006;24(3):541–52.
Kitamura D, Roes J, Kühn R, Rajewsky K. A B cell-deficient mouse by targeted disruption of the membrane exon of the immunoglobulin μ chain gene. Nature. 1991;350(6317):423–6.
Hiroi T, Yanagita M, Ohta N, Sakaue G, Kiyono H. IL-15 and IL-15 receptor selectively regulate differentiation of common mucosal immune system-independent B-1 cells for IgA responses. J Immunol. 2000;165(8):4329–37.
Waldmann TA. The biology of interleukin-2 and interleukin-15: implications for cancer therapy and vaccine design. Nat Rev Immunol. 2006;6(8):595–601.
Perera LP. Interleukin 15: its role in inflammation and immunity. Arch Immunol Ther Exp (Warsz). 2000;48(6):457–64.
Acknowledgements
This work was supported by the research grants “Genomic for Agriculture Innovation GMC009” from the Ministry of Agriculture, Forestry and Fisheries of Japan and a Grant-in-Aid for 2009 Multidisciplinary Research Project from MEXT in Japan from the Ministry of Education, Science, Sports, and Culture of Japan, and a grant from the Ministry of Health, Labor and Welfare of Japan awarded to T. Hiroi. L.P. Perera gratefully acknowledges the receipt of an invitational fellowship from the Japan Society for the Promotion of Science.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Yokoyama, S., Takada, K., Hirasawa, M. et al. Transgenic Mice that Overexpress Human IL-15 in Enterocytes Recapitulate Both B and T Cell-Mediated Pathologic Manifestations of Celiac Disease. J Clin Immunol 31, 1038–1044 (2011). https://doi.org/10.1007/s10875-011-9586-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-011-9586-7