
Abstract
Isahaya Bay in Japan has suffered from severe red tides. A previous study reported that red tides continued under dissolved inorganic nitrogen (DIN) depleted conditions in the bay. One possible explanation for this phenomenon is that regenerated NH4+, through organic matter decomposition, facilitates the persistence of red tides in the bay. Direct utilization of dissolved organic nitrogen (DON) by phytoplankton isolated from other coastal ocean areas has been reported. No study has been conducted on the direct utilization of DON by the phytoplankton community in Isahaya Bay. However, if phytoplankton in the Bay can utilize DON directly, DON probably contributes more to red tide formation and persistence than previously thought. In this study, field sampling and DIN and DON (in the form of amino acids and urea) enrichment bioassays were conducted to test whether growth of phytoplankton in Isahaya Bay can be stimulated through the addition of amino acids and urea. Phytoplankton growth rates in both the presence and absence of erythromycin, a bacterial inhibitor, were stimulated by NH4+, NO3−, and urea addition. The results suggest that phytoplankton in Isahaya Bay can be stimulated urea directly and indirectly. On the other hand, concentrations of DIN and urea were often depleted in the bay station throughout study period. Thus, DIN regeneration process might be an important process supporting red tides under DIN-depleted conditions. Notably, urea might prolong red tides through contribution to the NH4+ regeneration process, as urea is hydrolyzed to NH4+ by ureases produced by bacteria.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Red tides are a significant threat to fisheries and local and world economies. Due to human activities such as eutrophication and global warming, the distribution and frequency of red tides in coastal seas have increased worldwide over recent decades (Anderson 2009). In Japan, red tides have been occurring in many semi-enclosed bays, such as Tokyo Bay and the Ariake Sea, and have caused severe damage to coastal ecosystems and fisheries (Matsuoka 2003; Jia et al. 2018; Nakada et al. 2021).
The Isahaya Bay is located on the west coast of the Ariake Sea in western Japan and has suffered from severe red tides (Matsuoka 2003). In 1997, the inner part of Isahaya Bay was separated from the sea by dikes. Eutrophic high-turbidity water from a reservoir is frequently discharged into the bay. Since the Isahaya Bay Reclamation Project began in the early 1990 s, the frequency and intensity of red tides and hypoxia have become more severe in the Bay (Matsuoka 2003; Kim et al. 2018). Those environmental problems have also caused serious social problems (Hayami and Hamada 2016). However, the red tide formation mechanisms in the Bay are not fully understood.
A previous study has reported that red tides continued under dissolved inorganic nitrogen (DIN)-depleted conditions in Isahaya Bay (Li et al. 2016). One possible explanation of this phenomenon is that the red tides utilized regenerated NH4+ from the decomposition of organic matter in the drainage water (Takasu et al. 2019). We estimated that under the influence of the drainage water, the NH4+ regeneration rate increased by 31.5–46.8% in the water column in the bay (Takasu et al. 2019). Thus, regenerated NH4+ can support the persistence of red tides in the bay. On the other hand, direct utilization of dissolved organic nitrogen (DON) by isolated phytoplankton has been reported (Carpenter et al. 1972; Flynn and Bulter 1983). However, there are a few studies about direct utilization of terrestrial DON by natural phytoplankton in coastal systems (Fagerberg et al. 2009; 2010). No study has assessed the direct and indirect utilization of DON by phytoplankton in Isahaya Bay. If phytoplankton in the Bay can utilize DON directly, terrestrial organic matter loading from drainage may contribute more to red tide formation and persistence than previously thought.
This study examined DON utilization by phytoplankton assemblages in Isahaya Bay. Dissolved free amino acids (DFAA) and urea are two major components of terrestrial DON (Berman and Bronk 2003). Therefore, we selected alanine, glycine, and urea as DON for addition to the phytoplankton assemblages in Isahaya Bay. We found that urea may be as potentially important as DIN in providing a nitrogen source to phytoplankton assemblages in Isahaya Bay.
2 Methods
2.1 Sampling
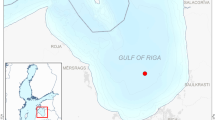
Water samples were collected with a small fishing vessel (F/V Hougei-maru) at fixed stations in the central part of Isahaya Bay on 20 April, 1 and 28 June, 21 July, 27 August, and 27 September, 2021 (Fig. 1). Water temperature and salinity were measured with a multi-parameter water quality meter (WQC-24; DKK-TOA, Tokyo, Japan). Surface water samples were collected using a 3 L Van Dorn water sampler (RIGO, Tokyo, Japan). Seawater samples for DIN, DFAA, urea, and dissolved inorganic phosphorus (DIP) analyses were filtered onboard the ship using pre-combusted 0.7 μm GF/F filters (Cytiva, Marlborough, MA, USA) and stored in 125 mL acid-washed high-density polyethylene bottles. Samples for chlorophyll-a (Chl a) analysis were poured into 125 mL amber polyethylene bottles. Samples for incubation experiments were poured into a 7 L high-density polyethylene tank through a 300 μm nylon mesh to remove mesozooplankton. Polyvinyl chloride gloves were worn during sample collection and processing to avoid contamination.

Maps of the Japanese Islands (a), Kyushu Island (b), and Ishaya Bay (c). Plots indicate sampling stations in the Ariake Sea and the reservoir. The incubation experiments were performed using seawater collected from the bay station
We also collected water samples for DIN, DFAA, urea, and Chl a analyses from the north and south discharge gates in the reservoir using a plastic bucket. Sample processing was done in the same manner as seawater.
2.2 Estimation of efficiency of the bacterial inhibitor
An antibiotic agent (erythromycin) was used during incubation to inhibit DIN regeneration through bacterial organic matter decomposition of DON to determine whether phytoplankton in Isahaya Bay can directly utilize amino acids and urea. During April sampling, we verified the efficiency of erythromycin as an inhibitor for bacterial organic matter decomposition. The screened seawater was filtered through pre-combusted 0.7 μm GF/F filters (General Electric, Coventry, UK) to remove particulate organic matter. Glucose (Fujifilm Wako Pure Chemical Industries, Osaka, Japan) was added to the filtrate at a final concentration of 5 mg L−1. The filtrate was poured into 100 mL acid-washed biological oxygen demand (BOD) bottles. Triplicate samples were used to estimate the initial dissolved oxygen (DO) concentration. Triplicate assays were spiked with erythromycin (Fujifilm Wako Pure Chemical Industries, Osaka, Japan) at a final concentration of 20 mg L−1 and incubated in the dark at in situ temperature. Ultrapure water (Milli-Q, Direct-Q UV3; Merck Millipore, Burlington, MA, USA) was used in place of erythromycin in the control experiment. After 2 days of incubation, samples were collected to measure DO concentration. The inhibition efficiency of erythromycin (I, %) was estimated from the following equation:
where DOint is the initial DO concentration (mg L−1), and DOfin-e and DOfin are the final DO concentration (mg L−1) with and without erythromycin, respectively.
2.3 Incubation experiments to estimate phytoplankton growth rates with various nitrogen sources
In the laboratory, 100 mL aliquots of seawater were filtered through 0.7 μm GF/F filters, and the filters were stored at− 20 °C until further analysis to estimate the initial Chl a concentration. The screened water was poured into acid-washed 250 mL polycarbonate bottles (100 mL in each bottle). Triplicate assays were spiked with NH4Cl (NH4+), KNO3 (NO3−), alanine, glycine, and urea (Fujifilm Wako Pure Chemical Industries, Osaka, Japan) at a final concentration of 25 μmol L−1 N, and ultrapure water was used in the control experiment. An identical series of triplicate assays was prepared and spiked with erythromycin (a final concentration of 20 mg L−1). The assays were incubated at in situ temperature, a light intensity of 3000 lx, and a 12:12 h light:dark cycle for 72 h. This incubation duration was selected, because we confirmed that the exponential growth phase of phytoplankton is maintained at least 72 h in Isahaya Bay (author’s unpublished data); this incubation duration was selected. The bottles were mixed once a daily to suspend the phytoplankton. After incubation, the samples were filtered and stored as above until further analysis.
2.4 Chemical and biological analyses
DO concentrations were measured by the Winkler titration method (Dickson 1994) using an automated titration system (AT-710; Kyoto Electronics, Kyoto, Japan).
DIN concentrations were determined colorimetrically with an ultraviolet–visible (UV–Vis) spectrophotometer (V-730; JASCO, Tokyo, Japan) using the cadmium column-diazotization method for NO3− + NO2− (Wood et al. 1967) and the phenol-indophenol method for NH4+ (Sagi 1966).
DFAA was determined through the fluorometric method with a fluorometer (FP-8300; JASCO, Tokyo, Japan) using the o-phthalaldehyde (OPA) derivatization method (Parsons et al. 1984). The standard curve for DFAA was determined using glycine, and total DFAA was calculated as a glycine equivalent. Notably, NH4+ interferes with this DFAA determination method (Tada et al. 1998). Thus, our DFAA values must be overestimated. Urea was determined by the colorimetrically with a UV–Vis spectrophotometer (V-730; JASCO, Tokyo, Japan) according to the method of Revilla et al. (2005), which is an improved method of the diacetyl monoxime method of Newell et al. (1967).
Dissolved inorganic phosphorus (DIP) concentrations were determined colorimetrically with a UV–Vis spectrophotometer (V-730; JASCO, Tokyo, Japan) using the molybdate method (Murphy and Riley 1962).
The Chl a concentration was determined from samples collected on GF/F filters and extracted with N, N-dimethylformamide (Fujifilm Wako Pure Chemical Industries, Osaka, Japan), and analyzed using fluorometry (FP-8300; JASCO, Tokyo, Japan), according to the method of Welschmeyer (1994).
2.5 Phytoplankton growth rate
We found a significant positive correlation between phytoplankton cell number and Chl. a concentration in the northern part of Ariake Sea (r2 = 0.89, p < 0.01, n = 7, author’s unpublished data) when Skeletonema sp. was dominant (mean ± sd, 76 ± 28%). Although we did not analyze phytoplankton composition in this experiment, it has reported that Skeletonema sp. is often dominant in the surface layer during summer in Isahaya Bay (Nakajima et al. 2016). Thus, we used Chl. a as a proxy for phytoplankton biomass. The growth rates (μ, days−1) of phytoplankton were calculated from the Chl a concentrations at the start and end of the incubation experiment, assuming that phytoplankton growth follows the exponential model:
where t is the duration of incubation (days), and N0 and Nt are the Chl a concentrations (μg L−1) at the beginning and end of the incubation, respectively.
2.6 Statistical analysis
Statistical differences in phytoplankton growth rates were analyzed using one-way analysis of variance (ANOVA), and the differences among means were analyzed using the Tukey–Kramer multiple comparison test. All statistical analyses were performed using R ver. 3.3.3 (R Development Core Team, 2017).
3 Results and discussion
3.1 Occurrence of red tides, and DIN, DFAA, and urea concentrations in the reservoir and Isahaya Bay
In Isahaya Bay, Chl a concentrations were higher than 10 μg L−1 except in April (Table 1). According to the definition of Tsutsumi et al. (2015), a Chl a concentration higher than 10 μg L−1 is considered a red tide. Thus, the red tides occurred from June through September during our study period.
In both Isahaya Bay and the reservoir, DIN (NH4+ + NO3+2−) concentrations were generally higher than DFAA and urea (Table 1). However, urea concentrations were comparable to DIN’s in the reservoir on 20 April and 1 June. Concentrations of DIN, urea, and DIP were often higher in the reservoir than in Isahaya Bay, suggesting that this drainage might be an important source of DIN, urea and DIP for the Bay throughout the study period. Because urea accounts for 40% of total nitrogen fertilizer used globally (Matthews 1994), runoff from agricultural areas may be an important source of urea in the reservoirs.
3.2 Phytoplankton growth rates with various nitrogen source
In the erythromycin addition experiment, DOint was 8.12 mg L−1, whereas DOfin and DOfin-e were 7.95 and 8.09, respectively (Table 2). Therefore, 82.4% of bacterial respiration was inhibited by the erythromycin at 20 mg L−1 (Table 2). This concentration can be regarded as effective in inhibiting nitrogen regeneration through bacterial organic matter decomposition.
We did not added DIP to the experimental bottles, and, therefore, phytoplankton might have been under DIP limitation during the incubation period (Table 1). However, we consider the following discussion is based on the finding that phytoplankton growth rates differed among treatments (Fig. 2). In all experiments, phytoplankton showed negative growth in the control (Fig. 2), probably due to nitrogen depletion during incubation. The addition of both DIN and DON stimulated phytoplankton growth in most experiments (Fig. 2). In all erythromycin-spiked all experiments, phytoplankton growth rates decreased sharply, except for the August experiment. Thus, regeneration of NH4+ through bacterial organic matter decomposition must be an important nitrogen source for phytoplankton, and this result is supported by a previous report (Takasu et al. 2019). On the other hand, phytoplankton growth rates in experiments with NH4+ and NO3− were strongly inhibited by erythromycin addition (Fig. 2). This finding might be explained by an inhibitory effect of erythromycin on eukaryotic phytoplankton, as erythromycin has been described as harmful to some phytoplankton, especially diatom (Sendra et al. 2018). Thus, the direct stimulation of phytoplankton growth caused by DIN and DON addition in this study must have been underestimated.

Phytoplankton growth rates with various nitrogen sources from samples taken on 20 April (a), 1 (b) and 28 June (c), 21 July (d), 27 August (e), and 27 September (f). Ala alanine, Gly glycine, Ery erythromycin. Error bars represent the standard deviation. Values labeled with the same letter were not significantly different at p < 0.05
In the August experiment, phytoplankton growth rates in experiments without erythromycin showed negative growth and were significantly lower than experiments with erythromycin. One possible explanation for this phenomenon is nitrogen depletion in the bottles during incubation. Bacteria rapidly consume nutrients when they decompose organic matter (Kirchman 2012; Takasu and Uchino 2021). Although we did not measure dissolved organic matter (DOM) concentration and bacterial production, Chl a concentration was extremely high in seawater sampled in August (Table 1). Thus, in August experiments, bacteria might compete with phytoplankton for nitrogen due to the active decomposition of phytoplankton-derived DOM.
In most experiments, phytoplankton growth rates in experiments with NH4+, NO3−, and urea did not significantly differ, with a few exceptions (Fig. 2), suggesting that phytoplankton growth in Isahaya Bay can be stimulated by urea in as manner similar to DIN, both directly and indirectly. As urea is the simplest form of DON, urea may be utilized by phytoplankton similarly to DIN in this study. On the other hand, phytoplankton growth rates in experiments with added alanine and glycine showed a different pattern to the NH4+, NO3−, and urea experiments. Although alanine and glycine also generally stimulated phytoplankton growth, the degree of stimulation was statically lower than NH4+, NO3−, and urea in the samples without erythromycin. However, phytoplankton growth rates in experiments with alanine and glycine addition did not significantly differ from experiments with NH4+, NO3−, and urea addition, with a few exceptions in erythromycin addition experiments (Fig. 2). In general, DFAA in seawater is rapidly used by bacteria (Nagata 2008), and therefore, phytoplankton could not use the alanine and glycine in experiments without erythromycin. Because bacterial activity is generally higher in warmer seasons (Kirchman 2012), DFAA might be quickly consumed by bacteria in summer season. Thus, DFAA may not contribute to red tide formation in Isahaya Bay as strongly as NH4+, NO3−, and urea. Some phytoplankton species prefer to use urea under nitrogen-starved growth conditions (Tada 2019). In Isahaya Bay, DIN concentration was often depleted (Table 1; Takasu et al. 2019). In addition, the concentration of urea was also often depleted and was lower than that of DIN at the bay station throughout the study period (Table 1). Thus, the NH4+ regeneration process (Takasu et al. 2019) might be more important for supporting red tides under DIN-depleted condition. In contrast, urea may prolong red tides through its contribution to the NH4+ regeneration process, as urea has two nitrogen atoms in one molecule and is hydrolyzed to NH4+ by ureases produced by bacteria. The DIN and urea concentrations in Isahaya Bay are similar to those in the Seto Inland Sea (Tada et al. 1998). Thus, the results of this study are applicable to similar coastal regions that receive terrestrial inflows.
3.3 Conclusions
We found that urea concentration was often higher in the reservoir than in Isahaya Bay. Furthermore, phytoplankton growth rates in the presence or absence of erythromycin were stimulated by NH4+, NO3−, and urea addition. These results suggest that the growth of phytoplankton in Isahaya Bay can be stimulated by urea both directly and indirectly. On the other hand, the concentration of urea was often depleted and lower than that of DIN at the bay station throughout the study period (Table 1). Thus, the NH4+ regeneration process might be more important to red tides under DIN-depleted conditions. On the other hand, urea might prolong red tides through its contribution to the NH4+ regeneration process, as urea is hydrolyzed to NH4+ by ureases produced by bacteria. These results indicate that reducing both DIN and DON input from drainage is necessary to control the red tides in Isahaya Bay. To understand fully the mechanisms of red tide formation in Isahaya bay, further study is needed to estimate quantitatively the contribution of DON as a nitrogen source to red tide formation.
References
Anderson DM (2009) Approaches to monitoring, control and management of harmful algal blooms (HABs). Ocean Coast Manag 52:342–347
Berman T, Bronk DA (2003) Dissolved organic nitrogen: a dynamic participant in aquatic ecosystems. Aquat Microb Ecol 31:279–305
Carpenter EJ, Remsen CC, Watson SW (1972) Utilization of urea by some marine phytoplankters. Limnol Oceanogr 17:265–269
Dickson AG (1994) Determination of dissolved oxygen in sea water by Winkler titration. WOCE operations manual: the observational programme, section 3.1: WOCE hydrographic programme, part 3.1.3: WHP operations and methods, vol 3. WHP Office Report WHPO 91–1, WOCE Report No. 68/91 November 1994, Woods Hole. https://www.nodc.noaa.gov/woce/woce_v3/wocedata_1/whp/manuals. htm Accessed 18 Mar 2017
Fagerberg T, Carlsson P, Lundgren M (2009) A large molecular size fraction of riverine high molecular weight dissolved organic matter (HMW DOM) stimulates growth of the harmful dinoflagellate Alexandrium minutum. Harmful Algae 8:823–831
Fagerberg T, Jephson T, Carlsson P (2010) Molecular size of riverine dissolved organic matter influences coastal phytoplankton communities. Mar Ecol Prog Ser 409:17–25
Hayami Y, Hamada T (2016) Isahaya bay, freshwater lake to an estuary again. Mar Poll Bull 102:250–255
Jia R, Lei H, Hino T, Arulrajah A (2018) Environmental changes in Ariake Sea of Japan and their relationships with Isahaya Bay reclamation. Mar Poll Bull 135:832–844
Kim S, Hayami Y, Tai A, Tada A (2018) The mechanism of bottom water DO variation in summer at the northern mouth of Isahaya Bay, Japan. J Oceanogr 74:595–605
Kirchman DL (2012) Processes on microbial ecology. Oxford University Press, Oxford, p 312
Li H, Matsushita K, Kanzaki M, Sugihara Y, Matsunaga N (2016) Formative process of large scale red tides in Isahaya Bay. J Jpn Soc Civ Eng B2 72:I_1243-I_1248 (in Japanese with English abstract)
Matsuoka K (2003) Recent changes in red tide-causing plankton in Isahaya Bay. Kaiyo Monthly 35:246–251 (in Japanese)
Matthews E (1994) Nitrogenous fertilizers: global distribution of consumption and associated emissions of nitrous oxide and ammonia. Global Biogeochem Cycles 8:411–439
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chem Acta 27:31–36
Nagata T (2008) Organic matter-bacteria interactions in seawater. In: Kirchman DL (ed) Microbial ecology of the oceans, 2nd edn. Wiley, New York, pp 207–241
Nakada T, Katano T, Hashimoto K, Kagami M (2021) Phytoplankton species abundance in Tokyo Bay (Japan) from 1998 to 2019. Ecol Res 36:901–906
Nakajima M, Yokoyama Y, Uno K, Yanaga J, Takemoto M, Ono E, Honjo T (2016) Prefictions and characteristics of Chattonella and diatom red tide bloom cessation during summer in Isahaya Bay. J Jpn Soc Civ Eng B2 72:I_1249-I_1254 (in Japanese with English abstract)
Newell BS, Morgan B, Cundy J (1967) The determination of urea in seawater. J Mar Res 25:201–202
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis. Pergamon Press, Oxford, p 173
R Development Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Revilla M, Alexander J, Glibert PM (2005) Urea analysis in coastal waters: comparison of enzymatic and direct methods. Limnol Oceanogr Methods 3:290–299
Sagi T (1966) Determination of ammonia in sea water by the indophenol method and its application to the coastal and offshore waters. Oceanogr Mag 18:43–51
Sendra M, Damián-Serrano A, Araújo CVM, Moreno-Garrido I, Blasco J (2018) Erythromycin sensitivity across different taxa of marine phytoplankton. A novel approach to sensitivity of microalgae and the evolutionary history of the 23S gene. Aquat Toxicol 204:190–196
Tada K (2019) Coastal environment and phytoplankton growth: field observations and laboratory experiments. Bull Coast Oceanogr 56:97–103 (in Japanese with English abstract)
Tada K, Tada M, Maita Y (1998) Dissolved free amino acids in coastal seawater using a modified fluorometric method. J Oceanogr 54:313–321
Takasu H, Komorita T, Okano T, Kuwahara M, Hoshimoto K (2019) Influence of water discharged from a reservoir on reclaimed land into Isahaya Bay (Kyushu, Japan) on the regeneration of NH4+ in the water column. J Oceanogr 75:299–304
Takasu H, Uchino K (2021) Nutrient regeneration from riverine high-molecular-weight dissolved organic matter through marine bacterial decomposition in a eutrophic coastal system: The Ariake Sea Japan. J Sea Res 177:102114
Tsutsumi H, Takamatsu A, Nagata S, Orita R, Umehara A, Komorita T, Shibanuma S, Takahashi T, Komatsu T, Montani S (2015) Implications of changes in the benthic environment and decline of macro-benthic communities in the inner part of Ariake Bay in relation to seasonal hypoxia. Plankton Benthos Res 10:187–201
Welschmeyer NA (1994) Fluorometric analysis of chlorophyll-a in the presence of chlorophyll-b and pheopigments. Limnol Oceanogr 398:1985–1992
Wood ED, Armstrong FAJ, Richards FA (1967) Determination of nitrate in sea water by cadmium-copper reduction to nitrite. J Mar Biol Assoc UK 47:23–31
Acknowledgements
We express our deep thanks to the captain Hidenori Matsunaga of the F/V Hougei-maru for his kind assistance during the field sampling. This study was supported in part by JSPS KAKENHI grant no. 21K05748 and Planetary Health Project at Nagasaki University to HT. We are grateful to the editor and anonymous reviewers as their comments helped us to greatly improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Takasu, H., Miyake, Y., Shiragaki, T. et al. Urea is a potentially important nitrogen source for phytoplankton during red tide formation in Isahaya Bay, Japan. J Oceanogr 78, 177–183 (2022). https://doi.org/10.1007/s10872-022-00640-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10872-022-00640-x