
Abstract
The inner part of Isahaya Bay was converted to a freshwater reservoir following the closure of the land claim dike in 1997. Turbid water drains into Isahaya Bay when water levels increase. We investigated whether particulate organic matter (POM) from the reservoir in Isahaya Bay has caused bottom organic enrichment in the northern part of Ariake Bay. Using potential end-members from before to after the rainy seasons, during which a frequent discharge from the reservoir was expected, stable isotope analyses were performed on sediments collected from Isahaya Bay and northern Ariake Bay. Each end-member was isotopically differentiated by δ13C and δ15N (riverine POM: −28.5 to −27.2‰ and 3.3–4.6‰; reservoir POM: −25.7 to −25.3‰ and 7.4–8.4‰; marine POM: −21.8 to −19.7‰ and 6.7–7.6‰; microphytobenthos estimated from consumers: −16.1 to −15.9‰ and 5.2–6.1‰, respectively). Sediment isotopic signatures fell within the mixing space defined by the signatures of the end-members. Marine POM contributed greatly to bottom sediments in both seasons in Isahaya Bay and Ariake Bay, ranging from ca. 60–70 and 40–60%, respectively. Reservoir POM contributed around 10% to bottom sediments. This percentage slightly increased in the sediment of Isahaya Bay after the rainy season, but decreased in the sediment of Ariake Bay. Thus, most of the POM discharged from the reservoir would not reach the northern part of Ariake Bay and would not be a major contributor to organic enrichment. This study is the first to quantitatively describe the contribution of drained reservoir POM outside Isahaya Bay.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Estuaries often constitute a huge ecosystem composed of local habitats that are linked with the transportation of materials and organisms. Some tidal-flat zones fuel the production of adjacent subtidal zones by the flushing of organic matter (Yoshino et al. 2012; Kang et al. 2015; Willems et al. 2016), whereas others function as water-purification systems based on filtration by clams inhabiting the system (Prins et al. 1996; Newell 2004; Grizzle et al. 2008). Several saltmarshes and reed beds are used as feeding grounds for fishes during high tide, which also links the trophic pathway between habitats and adjacent waters (Whitfield 2017 and the references therein). Estuarine habitats have been exposed to human activities worldwide (reviewed by McLusky and Elliot 2004). Thus, anthropogenic impact should be carefully evaluated to implement effective conservation and for the wise use of estuarine environments by taking habitat connectivity into account. Since the magnitude of human activity and habitat connectivity differs depending on the estuary, anthropogenic effects might also vary from one region to another.
Here, we focused on how anthropogenic activities affect the transportation of suspended particles between habitats in Ariake Bay. Ariake Bay is the largest bay in Kyushu, Japan, with the northwestern part being called Isahaya Bay. Although a bare mudflat zone once existed in the inner part of Isahaya Bay, a reclamation dike constructed in 1997 closed this section, making way for new agricultural land and preventing disasters that resulted from high tides related to storm events. Before reclamation, many large filter feeders were distributed in the mudflat, including the arcid bivalve Tegillarca granosa (Sato 2006). Thus, the mudflat functioned as a water-purification site, due to the presence of the filter feeder, and as a trophic subsidy due to the outwelling of microphytobenthos, similar to that observed for the mudflat of in the northern part of Ariake Bay (Yokoyama et al. 2009; Yoshino et al. 2012). However, these ecosystem functions have been completely lost since reclamation.
Reclamation caused problems beyond the simple loss of the mudflat. Although the inner part of Isahaya Bay has now become a freshwater reservoir (salinity below 1.5), the water is highly turbid (Umehara et al. 2012), with harmful cyanobacterial blooms frequently occurring in the reservoir (Umehara et al. 2012, 2015, 2017; Takahashi et al. 2014). The reservoir water is occasionally drained from the northern and/or southern gates during low tides to maintain freshwater conditions, as well as a constant water level. As a result, the organisms from all levels of the food web outside the reservoir are polluted by the water that is drained, as it contains the toxins of cyanobacteria (microcystin; Umehara et al. 2012, 2015; Takahashi et al. 2014).
This pollution also spreads beyond Isahaya Bay. Recently, Umehara et al. (2017) detected microcystin from the surface sediments in the northern part of Ariake Bay. Although microcystin is a soluble material, the suspended particles from the drainage are also transported in a northerly direction out of Isahaya Bay. Severe summer hypoxia has frequently occurred since (at least) the 2000s in the northern part of Ariake Bay (Tsutsumi 2006; Hamada et al. 2007; Tokunaga et al. 2011), causing the benthic ecosystem to deteriorate (Yoshino et al. 2007, 2010; Orita et al. 2016; Yamada et al. 2016). A potential cause of frequent hypoxia is an increase in organic loading (see Hayami 2007 and references therein). The increase in the organic loads is caused by a recent increase in the frequency of red tides in the northern part of Ariake Bay (Tsutsumi et al. 2007). However, if suspended particles are transported to the northern part of Ariake Bay, reservoir drainage might facilitate bottom organic enrichment and hypoxia. However, the particles suspended in freshwater might be quickly deposited when they meet seawater (Little 2000; Komorita et al. 2014). Cyanobacterial cells would quickly breakdown after drainage due to a sudden rise in salinity (Preece et al. 2017). Microcystin, which is detected from the sediments in northern Ariake Bay (Umehara et al. 2017), might be present because it is transported as a dissolved element to the northern part of Ariake Bay, followed by subsequent adsorption to sediments (Song et al. 2014). Thus, the transportation of suspended particles from the reservoir to the northern part of Ariake Bay requires investigation.
Stable isotopes are a powerful tool for evaluating the transport of materials because the isotopic signatures of source materials reflect their origin (Fry and Sherr 1984). When considering the target signatures, the proportional contribution of each source using mixing models based on the mass balance could be estimated (reviewed by Phillips et al. 2014). Stable isotope analyses of sediments have been performed in both Isahaya Bay and the inner part of Ariake Bay (Okamura et al. 2005; Yamamoto et al. 2006; Yokoyama and Ishihi 2009); however, these studies did not evaluate the effect of the drainage from the reservoir on the environment of northern Ariake Bay, because of the lack of information on reservoir sediment and particulate organic matter (POM). In this study, we measured the stable isotopes of POM in the water and surface sediments of the reservoir and the stations that are cross-sectionally arranged from Isahaya Bay to the northern part of Ariake Bay. The stable isotopes were measured before and after the rainy seasons, during which a high amount of discharge was expected. We evaluated the transportation of suspended particles from the reservoir to the bottom of the northern part of Ariake Bay, due to drainage caused by seasonal changes in the proportional contribution of the reservoir POM to the sediment of the northern part of Ariake Bay.
2 Materials and methods
2.1 Study site and sample collection
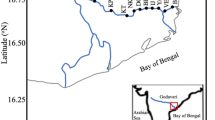
POM and sediment samples were collected in June and August 2016, before and after the rainy seasons, respectively. The samples were collected from the reservoir on June 7 and August 6, from Isahaya Bay and the northern part of Ariake Bay on June 14 and August 10, and from an upper reach of the Chikugo River on June 17 and August 8 (Fig. 1). The Chikugo River is the largest river discharging into Ariake Bay. Its annual average flow rate is 115.1 m3/s, whereas that of other rivers is 40 m3/s (Sato and Takita 2000). Thus, most riverine materials move into Ariake Bay from the Chikugo River. The average flow rates in July and August 2016 were 293.4 and 61.7 m3/s, respectively (http://www.qsr.mlit.go.jp/chikugo/gaiyou/suishitsu/index.html). A high amount of drainage also occurred during the study period from the north gate of the reservoir (Fig. 2). The northern part of Ariake Bay is a macrotidal estuary in which the tidal range extends up to approximately 7 m (Sato and Takita 2000). While salinity intrudes about 15–20 km upstream from the river mouth in the Chikugo River (Suzuki et al. 2009), the effect of tide reaches up to 23 km upstream where the dam, Chikugo Oozeki (N33.3039, E130.4824, not shown in Fig. 1), is located (Sato and Takita 2000). To collect riverine POM, surface water was collected by casting a bucket with a rope from the shore about 500 m upstream of the dam. As with other POMs, we sampled water at 1 m depth using a Van Dorn water sampler at the reservoir station near the north gate of the reclamation dike, station Is2 from Isahaya Bay, and station S3 from the northern part of Ariake Bay. A 1.5–2.5 L volume of water was filtered onto precombusted (450 °C for 2 h) and weighed Whatman GF/F filters on the same day as collection and was frozen at −30 °C until analysis.

Location of the sampling stations. The sampling station of riverine POM on the Chikugo River is located further upstream, and is not presented on the map

Amount of drainage per day from the north gate of the reservoir from June 1 to August 9 in 2016
Target sediment samples were collected from the boat using a hand grab at six subtidal stations in Ariake Bay and Isahaya Bay, in addition to that of the reservoir. The sediment bottom at the stations in Isahaya Bay and Ariake Bay in 2016 was fine mud with around 90% silt-clay content, except for station Q, which had about 50% silt-clay content (Yoshino unpublished data). The top 1 cm of each of the three grab sediment cores (ca. 3 cm in diameter) was taken and preserved separately as a replicate. Sediment samples were also frozen at −30 °C until analysis. Several gastropods (Cerithidea largillierti, Salinator takii, and Littoraria sinensis) inhabiting the tidal flats of Kashima City (Shingomori tidal flat) were collected. C. largillierti found in saltmarsh habitats mainly feed on the diatoms of sediment (Wang et al. 2014). S. takii also feed on diatoms on the mudflat surface (Fukuda and Kosuge 2000), while L. sinensis are often found attached to rocks near the mudflat (Yoshino, personal observation). Alfaro (2008) reported that a closely related species, L. scabra, is a generalist herbivore that shifts its diet depending on the availability of diet type. L. sinensis individuals at our site were expected to feed on epilithic microphytobenthos. The average isotopic signature of their tissues was used to estimate that of microphytobenthos (hereafter MB) by subtracting general trophic enrichment by carbon and nitrogen assimilation (i.e., 0.3 and 3.4‰, respectively; Post 2002). Samples of sediment and benthos tissues were also preserved at −30 °C until analysis.
2.2 Sample processing and isotope analyses
Macrobenthos and surface sediment samples were dried at 60 °C in an oven for 24 h and were then ground to a powder. Sediment samples were also immersed in 1.2 N HCl until no gas occurred for the removal of carbonates. Tidal flat benthos tissue was immersed in a mixture of methanol and chloroform (1:2) to remove lipid components. The filtered POM samples were weighed after drying in an oven at 60 °C, and were then acidified using 12 N HCl fumes in a desiccator for 24 h to remove carbonates. All samples were again dried in an oven and placed in tin capsules (approximately 1 mg for benthos tissue, 20 mg for sediment, and whole filter for POM) before determining isotopic ratios of samples using a Thermo Fisher Scientific Delta V mass spectrometer coupled to a Thermo Finnigan Flash EA1112 elemental analyzer.
Stable isotope values were denoted as δ, which is a measure of the amount of a heavier isotope present in a sample relative to known standards. This parameter was calculated as follows:
where R indicates 13C/12C or 15N/14N, respectively. Standards for C and N were Pee Dee Belemnite and atmospheric nitrogen, respectively.
2.3 Statistical analyses
All analyses were performed using the freeware R ver. 3.3.2 (R Development Core Team 2017). We performed weighted two-way analyses of variance based on generalized least square for differences in the organic contents and isotope signatures of surface sediments between stations within seasons and for differences between seasons within stations using the function gls() of the MASS package. Multiple comparisons were performed with Dunn–Šidák type I error rate correction using the function lsmeans() of the package “lsmeans” with the sandwich estimator for considering the heterogeneity of variance among samples.
The proportional contribution of each end-member (i.e., POM and MB) to the surface sediment from each station was estimated by Bayesian mixing models using the Markov Chain Monte Carlo method. Because the isotopic signatures of POM from Isahaya Bay and Ariake Bay were similar, we combined them in the estimation to represent materials from marine production (see “Results”). The analyses were performed using the R package “simmr,” which is the upgraded version of the “siar” package (Parnell et al. 2010, 2013). In the analyses, the trophic enrichment factor was set to 0 in the nontrophic relationship among POM, MB, and sediments. Four Markov chains with lengths of 500,000 were run and the first 50,000 samples were discarded as burn-in steps. Thinning was adjusted from the default of 10–20 depending on the convergence of chains. Gelman–Rubin diagnostic statistics were less than 1.05 for all estimated parameters, which satisfied the rules of thumb for convergence assessment (less than 1.1) by Gelman et al. (2003).
3 Results
3.1 Organic contents of POM and sediment from each station
Both the carbon and nitrogen contents of POM significantly differed among sites in both seasons (Table 1). The carbon and nitrogen contents of reservoir POM and riverine POM were significantly lower than that of marine POM before the rainy season, but increased significantly after the rainy season (Table 1). While the C/N ratios of POM mostly ranged from 6 to 7 at all sites, those of the reservoir was significantly lower than at any other site in both seasons (Table 1).
The carbon and nitrogen contents of the surface sediments also significantly differed among stations in both seasons, but at most stations they were around 20 and 2.2 mg/g, respectively. However, nitrogen content was high in the three stations located in Isahaya Bay, as a whole (Table 2). Seasonal changes in nitrogen content were fundamentally small at most stations, but both carbon and nitrogen content significantly increased and decreased after the rainy season at stations Q (both t28 > 3.3, P < 0.003) and Is2 (t28 = 3.7, P < 0.001), respectively. Sediment C/N ratios were 10.8 or more in the reservoir, S3, and E, and were significantly higher than those at the other sites (all |t|28 > 3.6, all P < 0.018), except E before the rainy season, which showed large variance. The degree of seasonal change in the C/N ratios remained within 0.5, even though these changes were statistically significant (Table 2).
3.2 Isotopic signatures of end-members and surface sediments
The dual-isotope plot shows that the isotopic signatures of end-members clearly differed from one another in both seasons, except those from Isahaya Bay and Ariake Bay (Fig. 3). The riverine POM had the most depleted δ13C and δ15N, ranging from −28.5 to −27.2 and 3.3–4.6‰, respectively. The δ13C and δ15N of reservoir POM ranged from −25.7 to −25.3 and 7.4–8.4‰, respectively. MB had the most enriched δ13C, ranging from −16.1 to −15.9‰, while δ15N was between riverine and reservoir POM (5.2–6.1‰). The signatures of POM from Isahaya Bay and Ariake Bay differed from the other end-members, but their plots were similar to one another in both seasons. Although the δ15N of the POM in Ariake Bay was slightly enriched (7.6‰) compared with that of Isahaya Bay (6.7‰), the δ13C signatures were similar to one another (−21.8 and −21.2‰) before the rainy season. However, after the rainy season, the δ13C of the POM in Ariake Bay was more enriched (−19.7‰) than that of Isahaya Bay (−21.4‰), while the δ15N signatures were similar to one another (6.7 and 6.8‰).

Dual-isotope plots of POM, MB, and the surface sediment from each station before (upper panel) and after the rainy season (lower panel)
The isotopic signatures of the surface sediments fell within convex hulls encompassed by the range of the signatures of end-members (Fig. 3). In the reservoir, δ13C and δ15N ranged from −24.6 to −24.4 and 5.5–6.1‰, respectively, showing almost no seasonal changes. However, the enrichment in δ15N was statistically significant (t28 = 2.3, P = 0.031) and was associated with the enrichment of reservoir POM (Fig. 3). In Isahaya Bay, the δ13C and δ15N of the sediment were similar to those of POM at all stations, and had a range of −21.6 to −20.9 and 6.2–6.7‰, respectively. In Ariake Bay, the δ13C and δ15N of sediments at station Q before the rainy season were −21.5 and 6.4‰, respectively. These values were similar to those of Isahaya Bay. δ13C and δ15N at station Q were enriched by 1–1.4 and 0.8–0.9‰, respectively, when compared with values at the other two stations in Ariake Bay. However, both δ13C and δ15N at the two stations were significantly enriched after the rainy season, being similar to those at Isahaya Bay sediment (Fig. 3). Although the δ13C at station Q did not change between the two seasons, δ15N became significantly depleted by 0.8‰ after the rainy season (t28 = −3.4, P = 0.002).
3.3 Proportional contribution of POM to sediment
The isotopic signatures of POM from Isahaya Bay and Ariake Bay were similar to one another and, therefore, were combined to represent marine POM when estimating the proportional contribution to sediment composition. Figure 4 shows the proportional contribution of POM to surface sediments in Isahaya Bay based on the isotope mixing space (convex hulls). Most contributions were attributed to marine POM in both seasons, which represented more than 60% at the median in any station. The contribution of reservoir POM was less than 10% before the rainy season at all stations, but it increased after the rainy season at stations Is3 and Is2, together with an increase in the upper limits of the 95% credible interval. Is1 decreased slightly. Conversely, at all stations, the contribution of MB consistently decreased and riverine POM increased after the rainy season.

Proportional contribution of end-members to surface sediment at three stations in Isahaya Bay before (left) and after (right) the rainy seasons. Box plots are based on the posterior probability density. Error bars denote 95% credible intervals. Each value above the box indicates the median value. Reserv reservoir, MB microphytobenthos estimated from tidal flat benthos
Marine POM contributed the most to the sediment in both seasons at Ariake Bay (Fig. 5), although it tended to be lower than that at Isahaya Bay. Instead, both reservoir and riverine POM contributed a slightly higher proportion to Ariake Bay than to Isahaya Bay. However, the contribution of reservoir POM to Ariake Bay did not increase in a similar manner to that at Isahaya Bay, but rather consistently decreased with the upper limits of the 95% credible interval at all stations after the rainy season (Fig. 5). The seasonal changes in the contribution of POM at the other locations differed depending on the station. The contribution of riverine POM decreased after the rainy season at stations S3 and E, but increased at station Q. The contribution of MB minimally changed at station Q, decreased at station S3, and slightly increased at station E. The contribution of marine POM increased at stations S3 and E, but decreased at station Q.

Proportional contribution of end-members to the surface sediment of three stations in Ariake Bay before (left) and after (right) the rainy seasons. Box plots are based on the posterior probability density. Error bars denote 95% credible intervals. Each value above the box indicates the median value. Reserv reservoir, MB microphytobenthos estimated from tidal flat benthos
4 Discussion
Although the isotopic signatures of POM were similar in Isahaya Bay and Ariake Bay, the other end-members exhibited fundamental discrete isotopic signatures that reflected the different origins of the contributing materials. The carbon signatures of POM from Isahaya Bay and Ariake Bay were −18 to −22‰, reflecting materials stemming from marine phytoplankton (Fry and Sherr 1984). Meanwhile, riverine and reservoir POM had more depleted signatures than marine POM. Riverine POM was −27 to −28‰, with C/N ratios ranging from 6 to 8, indicating a freshwater phytoplankton source (Atkinson and Smith 1983). Reservoir POM showed a much more enriched nitrogen signature than riverine POM, and the carbon signature was also slightly enriched (−25.7 to −25.3‰), as reported in a previous study (−25.6‰, signature of marine POM near the north gate just after drainage; Komorita et al. 2014). The carbon signatures of tidal flat macrobenthos corrected for trophic enrichment were about −15.9 to −16.1‰, respectively, which was close to the general value for microphytobenthos (Fry and Sherr 1984).
In a comparison between the isotopic signatures of sediment and the end-members, the sediment from Isahaya Bay had almost the same POM values at all three stations in the bay, indicating that the origin of sediment is mostly primary production in the bay. The contribution of riverine POM was fundamentally less than 10%. This result is consistent with a previous study (Okamura et al. 2005). However, the carbon signatures of the sediment in Ariake Bay were more depleted before the rainy season, except for station Q near the mouth of Isahaya Bay. According to Okamura et al. (2005) and Yamamoto et al. (2006), the sediment in the northern part of Ariake Bay is highly affected by discharge from the Chikugo River. In fact, the mixing model showed that riverine POM contributed more than 20% to the sediment at stations S3 and E. However, the contribution of reservoir POM was around 10%.
This pattern did not change after the rainy season. We expected a shift in the isotopic signatures of Isahaya Bay and Ariake Bay sediment toward that of reservoir POM (i.e., depleted signatures) and that the contribution of reservoir POM would increase. Actually, we found slight increases in the contribution of reservoir POM to the sediment at stations Is3 and Is2 in Isahaya Bay, with a gradation from geographic proximity to the drainage gate after the rainy season. However, the carbon signature of the sediments in Ariake Bay became rather enriched after the rainy season, with reservoir POM contributions decreasing at all stations, despite an increase in the organic content of reservoir POM after the rainy season. The same pattern was found at station Is1, which was the most distant from the gate in Isahaya Bay. These results indicate that the POM draining from the reservoir did not reach the northern part of Ariake Bay. Freshwater suspended materials are likely to flocculate when they meet seawater, due to the reduced charge of the material surface (Little 2000). Reservoir POM was quickly deposited on the bottom close to the gate due to flocculation and/or particle size (Komorita et al. 2014). Alternatively, it might have been partly transported toward the southern part of Ariake Bay along the Shimabara Peninsula (Tai and Komatsu 2013).
Contribution estimates might change with variations in the isotopic signatures of each end-member. Suzuki et al. (2009) showed that the δ13C of POM in the Chikugo River ranges from −27.4 to −22.8‰ before flushing periods and −24.8 to −22.3‰ during flushing periods. These δ13Cs are enriched compared with that in our data. The δ13C of MB in this study also differed from that collected by the method used by Couch (1989) in May and July from a tidal flat off the Kase River (−22.2 to −20.3‰; Yokoyama et al. 2009) and a tidal flat off the Chikugo River (−20.3 to −15.4‰; Suzuki et al. 2009). We, therefore, re-estimated the proportional contributions of reservoir POM after the rainy season using the modified δ13Cs of riverine POM (−25.5 ± 2.6‰) and MB (−18.4 ± 2.6‰) by averaging the data above and our data. However, these parameters minimally differed to those before the rainy season (station Q: 9.7%; station S3: 13.4%; station E: 12.9%). Hence, the proportional estimation of reservoir POM would not be seriously affected by isotopic variation in the other end-members.
While the present study showed no conspicuous effect of reservoir POM at a short timescale, there is a study looking at the long-term effect on the water quality and sediments of the northern part of Ariake Bay. According to the advection/diffusion simulation by Tai and Komatsu (2013), although the materials from the reservoir initially spread toward the southern part of Ariake Bay along the Shimabara Peninsula, a considerable amount of material is transported to the northern part of Ariake Bay over 455 days. However, the simulation assumes that the material is conservative and does not consider the extent to which it degrades during transportation. Reservoir POM was mainly composed of labile materials, with C/N ratios ranging from 6 to 8. Over such a long timescale, it is likely to be decomposed or taken up by organisms before reaching the northern part of Ariake Bay.
Finally, we must pay attention to how the results obtained from mixing models are interpreted. The proportional contribution of an end-member in a mixing model is calculated by its isotope signature under the premise that it is a real end-member; however, the estimated proportion cannot necessarily guarantee this premise. In other words, if reservoir POM was actually transported to the northern part of Ariake Bay, the contribution would be approximately 10% (Fig. 5) of all of the end-members; yet, even if transportation is not real, the mixing model would estimate 10% contribution from the isotope signature. Our main aim was to confirm the premise for the transportation of reservoir POM. We, therefore, evaluated the contribution change of reservoir POM, expecting an increase after the rainy season similar to the sediment at two stations (i.e., Is2 and Is3) in Isahaya Bay; however, such changes could not be detected at any station in the northern part of Ariake Bay. Although no change could show a real contribution, we cautiously concluded that reservoir POM is not transported to the northern part of Ariake Bay, leading to a major impact on organic loading in the region.
In conclusion, this study tested whether drainage from the reservoir affected organic loads to the northern part of Ariake Bay, as a change between before and after rainy seasons in the proportional contribution of reservoir POM to the surface sediment. We detected no increase in POM after the rainy season at any station in the northern part of Ariake Bay, unlike that of Isahaya Bay. Therefore, most of the drained reservoir POM does not reach the northern part of Ariake Bay, unlike toxic materials (Umehara et al. 2012, 2017; Takahashi et al. 2014), and does not have a major effect on bottom organic loads in this area. However, our results must be interpreted with caution, and further studies are needed to clarify the effect of drainage from the reservoir on the northern part of Ariake Bay.
References
Alfaro A (2008) Diet of Littoraria scabra, while vertically migrating on mangrove trees: gut content, fatty acid, and stable isotope analyses. Estur Coast Shelf Sci 79:718–726
Atkinson MJ, Smith SV (1983) C:N:P ratios of benthic marine plants. Limnol Oceanogr 28:568–574
Couch CA (1989) Carbon and nitrogen stable isotopes of meiobenthos and their food resources. Estuar Coast Shelf Sci 28:433–441
Fry B, Sherr EB (1984) δ13C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contrib Mar Sci 27:13–47
Fukuda H, Kosuge T (2000) Gastropods II (Makigairui II). In: Sato M (ed) Life in Ariake Bay (Ariakekai no ikimono tachi). Kaiyusha, Tokyo, pp 138–147 (in Japanese)
Gelman A, Carlin JB, Stern HS, Rubin DB (2003) Bayesian data analysis, 2nd edn. Chapman and Hall/CRC, London
Grizzle RE, Greene JK, Coen LD (2008) Seston removal by natural and constructed intertidal eastern oyster (Crassostrea verginica) reefs: a comparison with previous laboratory studies, and the value of in situ method. Estur Coast 31:1208–1220
Hamada T, Hayami Y, Yamamoto K, Yoshino K, Ohgushi K, Hirakawa R, Yamada Y (2007) The observation of growth–decay process of hypoxic water mass in the head of Ariake Bay. Ann J Coast Eng JACE 54:1121–1125 (in Japanese with English abstract)
Hayami Y (2007) Hypoxia in the inner area of the Ariake Sea. Aquabiology 173:577–584 (in Japanese with English abstract)
Kang C-K, Choy EJ, Choi K-S, Hwang K, Kim J-B (2015) Linking intertidal and subtidal food webs: consumer-mediated transport of intertidal benthic microalgal carbon. PLoS ONE 10:e0139802
Komorita T, Umehara A, Tai A, Takahashi T, Tsutsumi H (2014) Short-term variation of high-turbidity water discharged from a reservoir of reclaimed land onto Isahaya Bay, Kyushu, Japan. Oceanogr Jpn 23:1–12 (in Japanese with English abstract)
Little C (2000) The biology of soft shores and estuaries. Oxford University Press, Oxford
McLusky DS, Elliot M (2004) The estuarine ecosystem, 3rd edn. Oxford University Press, Oxford
Newell RIE (2004) Ecosystem influences on natural and cultivated populations of suspension-feeding bivalve molluscs: a review. J Shellfish Res 23:51–61
Okamura K, Tanaka K, Kimoto K, Kiyomoto Y (2005) Distribution of organic matter and organic carbon stable isotope ratios in the surface sediment of inner Ariake Bay and Isahaya Bay, Japan. Oceanogr Jpn 15:191–200 (in Japanese with English abstract)
Orita R, Umehara A, Komorita T, Choi J-W, Montani S, Komatsu T, Tsutsumi H (2016) Contribution of the development of the stratification of water to the expansion of dead zone: a sedimentological approach. Estuar Coast Shelf Sci 164:204–213
Parnell AC, Inger R, Bearhop S, Jackson AL (2010) Source partitioning using stable isotopes: cope with too much variation. PLoS ONE 5:e9672
Parnell AC, Phillips DL, Bearhop S, Semmens BX, Ward EJ, Moor JW, Jackson AL, Grey J, Kelly D, Inger R (2013) Bayesian stable isotope mixing models. Environmetrics 24:387–399
Phillips DL, Inger R, Bearhop S, Jackson AL, Moore JW, Parnell AC, Semmens BX, Ward EJ (2014) Best practices for use of stable isotope mixing models in food-web studies. Can J Zool 92:823–835
Post DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83:703–718
Preece EP, Hardy FJ, Moore BC, Bryan M (2017) A review of microcystin detections in estuarine and marine waters: environmental implications and human health risk. Harmful Algae 61:31–45
Prins TC, Smaal AC, Pouwer AJ, Dankers N (1996) Filtration and resuspension of particulate matter and phytoplankton on an intertidal mussel bed in the Oosterschelde estuary (SW Netherlands). Mar Ecol Prog Ser 142:121–134
R Development Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0. http://www.R-project.org
Sato S (2006) Drastic change of bivalves and gastropods caused by the huge reclamation projects in Japan and Korea. Plankton Benthos Res 1:123–137
Sato M, Takita T (2000) What bay is Ariake Bay? (Ariakekai towa donna tokoroka). In: Sato M (ed) Life in Ariake Bay (Ariakekai no ikimono tachi). Kaiyusha, Tokyo, pp 9–31
Song H, Reichwaldt ES, Ghadouani A (2014) Contribution of sediments in the removal of microcystin-LR from water. Toxicon 83:84–90
Suzuki K, Sugimoto R, Kasai A, Nakayama K, Tanaka M (2009) Dynamics of particulate organic matter in the estuarine turbidity maximum of the Chikugo River estuary, Ariake Sea, in summer: influence of the fluctuation of freshwater discharge. Bull Jpn Soc Fish Oceanogr 73:149–160 (in Japanese with English abstract)
Tai A, Komatsu T (2013) Process of pollutant transport from the head of Isahaya Bay and deterioration of aquatic environment in the Ariake Sea. J Jpn Soc Civil Eng (Hydraulic Enginerring) 69:1375–1380 (in Japanese with English abstract)
Takahashi T, Umehara A, Tsutsumi H (2014) Diffusion of microcystins (cyanobacteria hepatotoxins) from the reservoir of Isahaya Bay, Japan, into the marine and surrounding ecosystems as a result of large-scale drainage. Mar Pollut Bull 89:250–258
Tokunaga T, Kodama M, Kimoto K, Sibahara Y (2011) Fractionation of oxygen consumption component in the inner western area of Ariake Bay, Japan. Doboku Gakkai Ronbunshu B267:I926–I930 (in Japanese with English abstract)
Tsutsumi H (2006) Critical events in the Ariake Bay ecosystem: clam population collapse, red tides, and hypoxic bottom water. Plankton Benthos Res 1:3–25
Tsutsumi H, Tsutsumi A, Takamatsu A, Kimura C, Nagata S, Tsukuda M, Komorita T, Takahashi T, Montani S (2007) Mechanisms for the expansion of hypoxic water in the inner areas of Ariake Bay during summer. Oceanogr Jpn 16:1–20 (in Japanese with English abstract)
Umehara A, Tsutsumi H, Takahashi T (2012) Blooming of Microcystis aeruginosa in the reservoir of the reclaimed land and discharge of microcystins to Isahaya Bay (Japan). Environ Sci Pollut Res Int 19:3257–3267
Umehara A, Komorita T, Tai A, Takahashi T, Orita R, Tsutsumi H (2015) Short-term dynamics of cyanobacterial toxins (microcystins) following a discharge from a coastal reservoir in Isahaya Bay, Japan. Mar Pollut Bull 92:73–79
Umehara A, Takahashi T, Komorita T, Orita R, Choi J-W, Takenaka R, Mabuchi R, Park H-D, Tsutsumi H (2017) Widespread dispersal and bio-accumulation of toxic microcystins in benthic marine ecosystems. Chemosphere 167:492–500
Wang S, Chu T, Huang D, Li B, Wu J (2014) Incorporation f exotic Spartina alternifola into diet of deposit-feeding snails in the Yangtze River estuary salt marsh: stable isotope and fatty acid analyses. Ecosystems 17:567–577
Whitfield AK (2017) The role of seagrass meadow, mangrove forests, salt marshes and reed beds as nursery areas and food sources for fishes in estuaries. Rev Fish Biol Fish 27:75–110
Willems T, Backer AD, Kerkhove T, Dakriet NN, Troch MD, Vincx M, Hostens K (2016) Trophic ecology of Atlantic seabob shrimp Xiphopenaeus kryoyeri: intertidal benthic microalgae support the subtidal food web off Suriname. Estuar Coast Shelf Sci 182:146–157
Yamada K, Miyamoto Y, Nakano T, Okamura K (2016) Inter- and intraspecific variation in anoxic survival among three bivalve species in intertidal and subtidal areas along the coast of Japan. Plankton Benthos Res 11:49–56
Yamamoto K, Hayami Y, Kasagi T, Miyasaka H, Ohgushi K, Yoshino K, Hirakawa R (2006) Distribution of the surface sediment in the inner part of the Ariake Sea, Japan. Ann J Coast EngJSCE 53:961–965 (in Japanese with English abstract)
Yokoyama H, Ishihi Y (2009) Zoning of the inner part of Ariake Sound based on principal component analysis of sediment factors. Nippon Suisan Gakkasihi 75:674–683 (in Japanese with English abstract)
Yokoyama H, Sakami T, Ishihi Y (2009) Food sources of benthic animals on intertidal and subtidal bottoms in inner Ariake Sound, southern Japan, determined by stable isotopes. Estur Coast Shelf Sci 82:243–253
Yoshino K, Yamamoto K, Hayami Y, Hamada T, Kasagi T, Ueno D, Ohgushi K (2007) Benthic fauna of the inner part of Ariake Bay: long-term changes in several ecological parameters. Plankton Benthos Res 2:198–212
Yoshino K, Hamada T, Yamamoto K, Hayami Y, Yamaguchi S, Ohgushi K (2010) Effects of hypoxia and organic enrichment on estuarine macrofauna in the inner part of Ariake Bay. Hydrobiologia 652:23–38
Yoshino K, Tsugeki N, Amano Y, Hayami Y, Hamaoka H, Omori K (2012) Intertidal bare mudflats subsidize subtidal production through the outwelling of benthic microalgae. Estuar Coast Shelf Sci 109:138–143
Acknowledgements
We would like to thank M. Ukon and H. Doi for ship-based investigations, and K. Okamura, M. Takahashi and members of the Seikai National Fisheries Research Institute for helping with isotope analysis. We are also grateful to Kyushu Agricultural Administration Bureau for providing the data of the amount of drainage from the reservoir. This research is a part of the contracted research of the specified nonprofit corporation Ariakekai Saisei Kikou, “The Project for Considering Policies on the Restoration of Ariake Bay” funded by Saga Prefecture. This research was also supported by Grant-in-Aid for Scientific Researches (nos. 15K18731 and 18K11625) to K. Yamada, the Ministry of Education, Culture, Sports, Science and Technology, and the Cooperative Monitoring Program of Ariake Sea (COMPAS) project of Saga University. Special thanks are due to three anonymous reviewers and a guest editor in chief for their critical reading and many invaluable comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Yoshino, K., Yamada, K. & Kimura, K. Does suspended matter drained from the Isahaya freshwater reservoir cause organic enrichment in the northern Ariake Bay?. J Oceanogr 74, 619–628 (2018). https://doi.org/10.1007/s10872-018-0486-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10872-018-0486-9