
Abstract
The intricate interplay of plant-soil-microbial formulations was explored using cyanobacterium-based mixes (Anabaena laxa C11, Nostoc carneum BF2 and Anabaena laxa RPAN8) as seed coating, followed by soil drench at selected growth stages in spinach (variety Pusa All green). A field experiment under shade net conditions was conducted and data recorded 2, 3, 5 WAS (Weeks after sowing) and harvest stage. Cyanobacterial treatments led to significantly higher pigment content (0.58-1.29-fold), along with substantial enhancement in the enzyme activities of nitrate reductase (0.33 - 0.49-fold), glutamine synthetase (0.35 - 0.53-fold) and phosphoenol pyruvate carboxylase activity (0.17 -0.38-fold) in leaves. RPAN8 treatment led to a notable increase in spinach yield, while BF2 treatment stimulated ascorbic acid (0.37-fold), antioxidants (0.36-fold) and β-carotene (0.41-fold) in fruits, over control. Overall, C11 treatment consistently exhibited significantly superior results in terms of growth, soil microbiological, yield and quality attributes. Principal Component analyses illustrated a strong positive correlation of fruit ascorbic acid with soil chlorophyll and available iron. Path coefficient analysis revealed positive direct effects of available nitrogen with PEPCo, nitrate reductase, chlorophyll a, carotenoids, leaf area and antioxidants at different sampling intervals. Such organic formulations can be a valuable resource-efficient strategy for improving spinach yield and quality.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Spinach (Spinacia oleracea) is a highly popular nutritious and versatile leafy vegetable crop that is widely consumed worldwide due to its health benefits, vitality, and flavor. In developing countries, including India, phytonutrients such as β-carotene are of immense significance, 95% of β-carotene comes from fruits and vegetables and, among these, leafy greens comprise a substantial portion of about 52% of its total consumption (Gopalan 1992; Anjana et al. 2007). Among them, spinach boasts of an impressive array of properties, from being antimicrobial to anticarcinogenic and antioxidant rich (Vázquez et al. 2013). In addition, spinach contains important constituents, vitamins such as ascorbic acid, folic acid, riboflavin and minerals, including iron, calcium, (Prakash and Pal 1991), making it an excellent nutraceutical and culinary option (Miller et al. 1995).
The art of nurturing seedlings is a cornerstone in successful vegetable crop establishment. Furthermore, it is crucial to comply with all relevant regulations like providing optimal growing conditions; hence, ecological factors need to be prioritized as well to produce high-value vegetable seedlings. In modern agriculture, farmers have become reliant on synthetic fertilizers, pesticides, intensive ploughing, and intensive irrigation. While these approaches have been essential in keeping up with food demands of developing nations, they have raised some environmental and health concerns as well. The use of organic fertilizers, particularly biofertilizers or microbial consortia to enhance plant productivity are sustainable options to reduce the pressure on the environment (Cakmakci et al. 2009; Dineshkumar et al. 2019; Kholssi et al. 2021). Leveraging the beneficial effects of interactions between plants and microbes can lead to greater nutrient availability and their mobilization, which can lead to better growth of plants. This can be a boon for developing environmentally sustainable solutions for tackling the global challenge of dwindling crop yields.
Among photosynthetic prokaryotes, cyanobacteria with their multifaceted abilities, including beneficial effects on plant and soil health can help to promote eco-friendly farming practices for sustaining soil fertility and plant nutrition (Gupta et al. 2013; Renuka et al. 2018). When integrated effectively into agriculture, cyanobacteria hold significant potential for substantially augmenting crop yields (Higa and Wididana 1991). Incorporation of various cyanobacterial species into the soil illustrated their impact on growth, health and productivity across diverse crops (Garlapati et al. 2019; Kholssi et al. 2021; Prasanna et al. 2021). These positive effects are conventionally attributed to the provisioning of nutrients essential for the growth of plants and improvement in soil tilth, infiltration of water and nutrients and storage capacity of soil (Singh et al. 2016; Carillo et al. 2020). Several researchers also have highlighted the role of cyanobacteria in nutrient cycling and use efficiency, emphasizing their ability to enhance mineral content in soils and promote plant growth (Rana et al. 2012; Prasanna et al. 2018; Shivay et al. 2022).
Although there is evidence supporting the efficacy of cyanobacteria in enhancing yields in various vegetable and fruit crops (Geries and Elsadany 2021), studies investigating their impact on the commercial and qualitative aspects of spinach remain limited. The application of aqueous extract of Anabaena sphaerica on spinach resulted in enhanced plant dry weight, improved nutrient absorption and elevated protein content in the leaves (Mahmoud et al. 2019). Salamah et al. (2019) used Nostoc sp. SO-A31 to bring about significant enhancement in the growth of water spinach. Such interventions led to improvement in biomass yield, leaf numbers and vigorous stem and root development, even under nitrogen deficit conditions.
Besides nutritional inputs, vegetables also need to be protected from excessive sunlight and other environmental stressors, for which shading nets are commonly used in tropical and subtropical regions (Fallik et al. 2008). Shade net cultivation also can help growers to overcome the challenges of unfavorable natural conditions by enhancing the thermal climate, leading to improved crop protection against excessive radiation, insects, and environmental fluctuations (Kittas et al. 2008) and produce high-quality vegetables more efficiently and profitably.
With this background, the primary objective of the present investigation was aimed towards evaluating the efficacy of combining cyanobacteria-based mixes as seed coating along with drench application in stimulating the growth, yield and quality aspects of Pusa All Green variety of spinach at different stages of maturity, when grown under shade net conditions. Moreover, the novelty of this research lies in developing a nutrient-enriching strategy as an environmentally friendly organic option for boosting spinach cultivation.
Materials and methods
Cultivation of cyanobacterial strains for drench application and their maintenance
Anabaena laxa C11, Nostoc carneum BF2 and Anabaena laxa RPAN8 belong to the Division of Microbiology, IARI, New Delhi, India, and are maintained routinely by culturing in a nitrogen-free BG11 medium in Haffkine flasks (Stanier et al. 1971) at 27±1 °C under 16 h light and 8 h darkness. The cultures (thereafter referred to as C11, BF2 and RPAN8) were grown in stationary mode, and subjected to white light with an intensity of 50-55 μmol photons m-2 s-1. These cultures are deposited in the National Agriculturally Important Microbial Culture Collection (NAIMCC), ICAR-National Bureau of Agriculturally Important Microorganisms (NBAIM), India.
These cyanobacterial cultures serve as the basis for developing seed coating and soil drenching formulations. The cultures were harvested, meticulously rinsed with distilled water to eliminate any residual media components, and then blended with a carrier mixture containing vermiculite and paddy straw as its constituents (1:1 mix of Vermiculite:paddy straw compost, having a C/N ratio of 16.22:1) The humus content of paddy straw compost is 13.8%, and pH is 7.34 to which the dry powdered biomass of cyanobacterial cultures was added. The amended mix contained 2-4 μg chlorophyll g-1 carrier. Chlorophyll was used as an index of biomass, as the CFU of filamentous cyanobacteria is often erratic. The treatments for the spinach variety (Pusa All green) included - T1-Control, T2-A. laxa C11, T3-N. carneum BF2 and T4-A. laxa RPAN8, as both seed coating and drench formulations. The drench formulations of C11, BF2 and RPAN8 were prepared by blending 300 g of carrier mixture and those amended with cyanobacteria in 3 L of tap water. After thorough blending, the mixture was allowed to incubate in darkness for 48 h, with periodic agitation. Subsequently, after filtration through a muslin cloth, as formerly optimized (Prasanna et al. 2018; Bhardwaj et al. 2024), these drench formulations were stored at 4°C for further analyses and use.
Analyses of drench
The drench formulations were also analyzed in relation to the content of total, released and reducing sugars, proteins, indole acetic acid (IAA), total phenolic and glomalin content. The total and released sugar content were expressed as µg mL-1, after being assayed in drench formulations using the method of Dubois et al. (1956). The reducing sugars assay was carried out using 3,5-dinitrosalicylic acid (DNSA) reagent by the method of Miller (1959) and the values expressed as µg mL-1. In the drench formulations, the total soluble proteins were quantified spectrophotometrically (Herbert et al. 1971) with bovine serum albumin as standard. The IAA in the formulations was quantitatively determined by the Salkowski method through spectrophotometric measurement at 530 nm (Gordon and Weber 1951). Total phenols in the drench formulations was quantified, following Singleton et al. (1999) using Folin–Ciocalteu reagent and spectrophotometric analysis, where the development of blue color at 660 nm was measured. The total phenolic content was expressed in μg caffeic acid equivalent (CAE) per mL of formulation mix. Glomalin-related soil proteins in the drench formulations were extracted and quantified, as per the standard procedure (Wright and Upadhyaya 1996).
Experimental set-up
The performance of the carrier-based mixes of selected cyanobacterial strains was assessed in a field trial involving spinach (Spinacea oleracea; variety Pusa All Green) grown in 12 beds, each accommodating 1000 plants per bed, within a 4 m2 area. The experiment was set up in the shade nets covered polyhouse belonging to the Centre for Protected Cultivation and Technology (CPCT), ICAR-Indian Agricultural Research Institute (ICAR-IARI), New Delhi, situated at 28.4° N latitude, 77.2° E longitude, having an altitude of 228.6 m above mean sea level, in relation to the Arabian Sea. Prior to sowing, spinach seeds were initially coated with cyanobacterial carrier mix, involving a mechanized process known as seed-coating, carried out 24 h before sowing. The air-dried seeds were sown at the rate of 50 g per bed. At the time of sowing, the sandy loam soil had 166.67 kg ha-1 available N, available Fe and Zn as 1.13 and 0.95 mg kg-1, with dehydrogenase activity of 84.64 µg TPF g-1 soil d-1 and 84.77 µg g-1soil microbial biomass carbon.
The experimentation involved four treatments, arranged in a randomized block design (RBD), each with four replicates: T1 Control; T2 C11; T3 BF2; T4 RPAN8. The recommended chemical fertilizer doses for spinach crop were 50 kg ha-1 nitrogen, applied as two split doses of urea, with only one dose being given to treatments (T2-T4). At the time of sowing, single super phosphate (SSP), to provide 25 kg ha-1 phosphate and muriate of potash (MOP) applied to provide 50 kg ha-1 potassium were given as single basal doses. Irrigation was through drip system on alternate days, while fertigation was done twice a week, at 3-day intervals. The experimental area comprised a total of 12 beds, each containing 1000 seeds/saplings per bed, with an inter-row spacing of 1 m between each bed; the total row length was 4 m and the width was 1 m. Soil drenching using the formulations was initiated at 2 WAS (Weeks after sowing) and continued until 5 WAS, at 10-day intervals; application rate was 1000 mL bed-1, approximately 1 mL plant-1. Plant as well as rhizospheric soil samples were collected at 2, 3, 5 WAS and at the harvest stage. Analyses for various parameters, including soil microbiological attributes (soil chlorophyll, soil microbial biomass carbon, soil dehydrogenase), nutrient levels (soil organic carbon, available nitrogen, available phosphorus, available iron and zinc), leaf pigment composition, enzymatic assays (glutamine synthetase, nitrate reductase and phosphoenol pyruvate carboxylase activity), leaf nitrogen and calcium analyses. Plant yield as well as quality traits were recorded at harvest stage (60 days after sowing). All the plant and soil samples were analyzed immediately or stored at 4°C in sterile polybags for subsequent analysis.
Soil microbiological and nutrient analysis
Soil chlorophyll content was quantified utilizing soil cores collected from a depth of 0 -15 cm. These cores were treated with a mixture of acetone and DMSO (1:1) added at the rate of 4 mL per g of soil. After mixing, the samples were kept under dark at normal room temperature for 48 h. Periodic shaking at 24-h intervals ensured complete chlorophyll extraction. Spectrophotometric readings were taken at 663, 645, 630, and 775 nm. The chlorophyll content in the soil samples was as quantified, represented the abundance of photosynthetic biomass (Nayak et al. 2004). The soil chlorophyll content was calculated using the SCOR/UNESCO formula and expressed in mg g-1 soil.
To assess the carbon content as microbial biomass (SBMC), soil samples equivalent to 3.5 g in dry weight were added to glass vials. In one batch of vials, chloroform was added, followed by sealing the lids tightly and vigorous shaking. The other batch of vials has no chloroform added. Thereafter, the vials were incubated in dark for a day before being flushed with N2 under a fume hood at 40-50°C to remove chloroform completely. Upon complete removal of chloroform, both fumigated and un-fumigated samples were supplemented with 0.5 M K2SO4. These vials were shaken for 30 min and the resultant extracts were filtered through Whatman No.42 filter paper, the optical density at 280 nm was measured (Nunan et al. 1998), and microbial biomass carbon calculated using the following formula:
where ECF and ECNF represent the values of extractable carbon in both the fumigated (ECF) and non-fumigated soil samples (ECNF) respectively, with kEC having a value of 0.45.
Dehydrogenase activity was determined by incubating 6 g soil with a 3% triphenyl tetrachloride solution and measuring the absorbance at 485 nm (Casida et al. 1964). The values were expressed as μgtriphenyl formazan (TPF) g-1 soil d-1.
For macro and micronutrient analysis, rhizospheric soil samples were collected at 0 -15 cm depth using a soil auger and subsequently subjected to airdrying, grinding and sieving. Soil organic carbon content (SOC) was determined (Walkley and Black 1934) and expressed as a percentage. Available nitrogen (Subbiah and Asija 1956) and phosphorus content (Olsen 1954) in soil were also quantified. Available iron and available zinc were evaluated using a wet digestion method with a di-acid mixture and the concentrations measured at 248.4 nm and 213.7 nm, using an Atomic Absorption Spectrophotometer (Prasad 2006). The nitrogen content of spinach leaves was assessed at 2, 3, 5 WAS and harvest stage, through a process of digestion with H2SO4 and subsequent distillation; the quantity of acid consumed in the titration was measured, and the nitrogen content in the spinach leaves expressed as percent (%).
Estimation of pigment content in leaves
Spinach leaf samples were collected for photosynthetic pigment estimation, as per the standard method (Lichtenthaler 1987). For the pigment extraction process, 100 mg of fresh spinach leaves (without the roots) were weighed; an aliquot of 10 mL dimethyl sulphoxide (DMSO) was added and kept overnight in dark at room temperature. The volume was then made up to 10 mL using DMSO and optical density recorded at 480, 510, 645 and 663 nm. The formulae of Arnon (1949) for chlorophyll a, chlorophyll b, carotenoids and total pigments were used and expressed as mg g-1 fresh weight (FW):
where, V represents the total volume of DMSO added and W denotes the weight of spinach leaf sample in g.
Plant enzymatic assays
The activity of glutamine synthetase (GS) in leaf tissues was determined by measuring the absorbance at 540 nm denoted as µmoles γ-glutamyl hydroxamate produced per g fresh weight of leaves per min, following the method of Shapiro and Stadtman (1970). Nitrate reductase activity (NR) was determined as per the method of Snell and Snell (1961) and expressed as µmoles nitrite per g fresh weight. Phosphoenol pyruvate carboxylase activity (PEPCo) was determined following the protocol of Wu and Wedding (1985) and expressed as micromoles per g protein weight per min.
Yield characteristics
At harvest stage (120 days after sowing), spinach plants were randomly selected from each experimental plot, in triplicates, to record the biometric and yield parameters, including plant height (cm), root length (cm), number of branches, number of leaves, leaf area (m2 plant-1), yield (kg m-2) and leaf weight (g plant-1). Plant height was measured using a ruler scale at harvest stage for each individual plant. The height was assessed from the plant’s base to the tip of the main branch in 15 randomly tagged plants. Root length was the distance from the root's starting point at the base of the plant to its endpoint, and the mean data for 15 plants were documented. The number of branches was measured from the base of the stem where the branches originate. The individual number of branches was counted and their total number recorded at harvest stage. Healthy plants with an adequate number of leaves were chosen. Any deceased or impaired leaves were eliminated. The number of leaves was then subsequently ascertained by tallying them from the uppermost part of the spinach plant to the lowest. The leaf area of three plants was measured using an automated leaf area meter, and the mean leaf area was computed and documented at harvest stage as m2 per plant. The spinach plants were harvested at maturity phase and the weight of fresh leaves from each plant was measured in g. Subsequently, the relative yield was computed and expressed in kg m-2.
Quality traits estimation in leaves
Total soluble solids (TSS) of spinach leaves were determined by a handheld refractometer (ERMA; Japan) and expressed in °Brix, based on the method of Ranganna (1986). The firmness of spinach leaves (often denoted as film burst) was assessed by the optimized protocol of Sun et al. (2015), using a texture analyzer (model: TA.Xtplus, Stable Micro System, UK) equipped with a spherical probe. This assesses the firmness of spinach leaves through the application of a regulated force and measures the resultant resistance encountered which is then analyzed to obtain a firmness value (g m-2), which denotes the maximum force exerted during the puncture. Titrable acidity values were computed using the method given by Ranganna (1986). The extracts of the macerated spinach leaf samples in distilled water (10 g in 100 mL distilled water) were filtered through Whatman no. 1 filter paper and titrated with 0.1 N NaOH, using phenolphthalein as the indicator. Titrable acidity (%) can be expressed as:
Ascorbic acid (vitamin C) was measured by the visual titrimetric technique as outlined in AOAC (2000).
Protein content of spinach leaves was measured with bovine serum albumin (BSA) as standard (Herbert et al. 1971). The assessment of antioxidant activity in fresh spinach leaves was conducted using the CUPRAC (Cupric Reducing Antioxidant Capacity) method, following the procedure outlined by (Apak et al. 2004). Absorbance of the samples was measured at 460 nm and expressed as micromoles of Trolox equivalents per 100 g (Trolox equivalent antioxidant capacity-TEAC):
The total phenolic content in the raw spinach leaves were determined using the Folin-Ciocalteau reagent method as described by Singleton et al. (1999). The total phenolic content was expressed in mg of gallic acid equivalents (GAE) per 100 g of extract:
The β-carotene, carotenoids and vitamin A were also determined in spinach leaves using the formula given below.
The calcium content of spinach leaves was determined at harvest stage using a Flame photometer equipped with an LPG flame system, Jaisbro microprocessor and silicon photodiode detector, following the wet ashing digestion (di-acid digestion) method outlined by Raghuramulu et al. (1983). The values were expressed as mg calcium (100 g)-1 dry weight.
Statistical analysis
The experiment used a randomized block design (RBD), comprising four treatments, including control. The data obtained in triplicate, at four-different time intervals (2, 3, 5 WAS and harvest stage) were analyzed for the various soil, plant, quality and yield parameters at a probability level of 0.05 %, through ANOVA using WASP 2.0 (Web Agri Stat Package, Indian Council of Agricultural Research, India) statistical tools, with critical differences computed. Pearson’s Correlation coefficient analysis using GRAPE (GRAphics Programming Environment) software tool (v1.0.0) (Gopinath et al. 2020) was utilized. Path coefficient analysis was performed using Windostat version 8.5 software (Singh and Chaudhary 1977).
Results
Analyses of the drench formulations
The total sugar values were highest in T2 (C11) and T4 (RPAN8) and lowest in Control. As compared to control (uninoculated), the released sugars were 2- fold higher in T4 (RPAN8) over control; T2 (C11) had the highest reducing sugar content which was 2-fold higher over control. In terms of protein content, both T3 (BF2) and T4 (RPAN8) showed high values (43.39 and 36.73 µg mL-1). IAA values were 2-fold higher in T2 (C11; 39.86 µg mL-1) amended drench formulations compared to control. In terms of total phenol content, it was observed that T4 (RPAN8) showed higher values (36.43 µg CAE mL-1), as compared to control (T1). In terms of Glomalin content, T4 drench formulation (RPAN8) exhibited 18.72% higher values, as compared to control (Table 1).
Growth attributes, as influenced by cyanobacterial treatments
Total pigment content of the leaves was greater in cyanobacterium-treated plants and highest with C11 (0.95 mg g-1 FW) at 2 WAS, however, at later stages RPAN8 and BF2 were most promising respectively, as compared to control (Fig. 1a).

Influence of cyanobacterial treatments on photosynthetic pigments and enzymatic activities of spinach crop at different crop growth stages in terms of (a) Total pigments, (b) Glutamine Synthetase activity, (c) Nitrate reductase activity, (d) PEPCO. Error bars represent standard deviation of mean of three replicates. Abbreviations for treatments and attributes: T1: Control; T2:C11 (Anabaena laxa), T3: BF2 (Nostoc carneum), T4: RPAN8 (Anabaena laxa); WAS: weeks after sowing
The components of total pigments such as chlorophyll values were also higher in leaves of seeds treated with cyanobacteria +drench, as compared to the control. Chlorophyll a was higher with C11 treatment at all stages of crop growth (Fig. S1a) i.e., 2, 3, 5 WAS and harvest stage (72%, 67.74%, 1.02-fold and 38.29% in C11 treated plants respectively). Similarly, treatment with cyanobacteria resulted in the enhancement of chlorophyll b content of the leaves at all stages, which was higher in plants treated with C11 (0.50-fold increase at 2 WAS, 0.048-fold increase at 3 WAS, 1.09-fold increase at 5 WAS and 0.82-fold increase at harvest stage respectively), in comparison with control (Fig. S1b). carotenoid content showed no significant differences in the cyanobacterium treated plants as compared to the uninoculated control, particularly at harvest stage (Fig. S1c). At 2 and 5 WAS, carotenoid content was significantly higher in plants treated with C11 (62.5% and 80.76% increase over control) respectively. However, changes in carotenoids content were significantly higher at 3 WAS in cyanobacteria-treated plants, while BF2 and RPAN8 significantly enhanced the content in leaves (0.22 mg g-1 FW and 0.21 mg g-1 FW).
Activity of plant enzymes
A gradual reduction in glutamine synthetase activity in leaves was recorded with increasing age of the plants, and all three cyanobacterial treatments brought about a positive influence. Values ranged from 4.88 to 7.23 µmol γ-glutamyl hydroxamate g-1 FW min-1 at 2WAS, with highest in C11 (7.23 µmol γ-glutamyl hydroxamate g-1 FW min-1), followed by BF2 (6.05 µmol γ-glutamyl hydroxamate g-1 FW min-1) and RPAN8 (6.20 µmol γ-glutamyl hydroxamate g-1 FW min-1) respectively (Fig. 1b). In terms of cyanobacterial treatments, RPAN8 treatment recorded significantly higher values of 5.40 µmol γ-glutamyl hydroxamate g-1 FW min-1 respectively. At 5 WAS, the values ranged from 2.79 to 3.77 µmol γ-glutamyl hydroxamate g-1 FW min-1 and among the treatments, the top performers were C11 and BF2, respectively (3.77 and 3.73 µmol γ-glutamyl hydroxamate g-1 FW min-1). The cyanobacterial treatments, in particular, C11 considerably enhanced the glutamine synthetase activity even at harvest stage, by 53.02% compared with Control, followed by 37.2% enhancement by RPAN8, respectively.
The activity of nitrate reductase (NR) in leaf samples ranged from 183.09 to 256.80 µmol NO2 g-1FW at 2 WAS, 157.49 to 210.27 µmol NO2 g-1FW at 3 WAS, 132.34 to 180.31 µmol NO2 g-1FW at 5 WAS and 114.64 to 171.53 µmol NO2 g-1FW at harvest stage respectively (Fig. 1c), while among treatments (Fig. 1c), the highest values were observed in C11 (256.80, 194.64, 179.37 and 161.81 µmol NO2 g-1FW) and RPAN8 (250.35, 210.27, 180.31 and 171.53 µmol NO2 g-1FW) at all 4 stages respectively. Overall, C11 and RPAN8 were the top performers, at all stages of growth.
Phosphoenol pyruvate carboxylase (PEPCO) activity determined in leaves at 2, 3, 5 WAS and harvest stages were enhanced by all cyanobacterial treatments, up to harvest stage (Fig. 1d). BF2 was the top ranked, followed by C11 and RPAN8 at all stages, except at 2WAS, in which RPAN8 exhibited a greater increment.
Soil microbiological analyses
The soil microbial biomass carbon in treatments receiving cyanobacterial treatment (seed coating and drench) ranged from 94.47 to 132.47; µg g−1 soil, with highest in C11 (132.47 µg g−1 soil), followed by BF2 (117.29 µg g−1 soil), as given in Table 2. SMBC content at 3 WAS significantly increased in cyanobacterial treatments, as compared to control, with highest values recorded in C11 and RPAN8 203.81 and 223.63 µg g−1 soil, followed by BF2 (137.52 µg g−1 soil).A similar trend was also observed at 5 WAS, wherein T2 (C11) and T4 (RPAN8) showed highest total SMBC content (316.35 and 320.37 µg g−1 soil), followed by T3 (BF2; 238.60 µg g−1 soil) respectively. SMBC content showed a progressive increase in RPAN8 treatment throughout the period of the experiment. Dehydrogenase activity in soil enhanced with cyanobacterial treatments at all 4 stages. Among the cyanobacterial treatments, dehydrogenase activity in T2 (C11) was almost 40.72% higher at 2WAS, with 46.09% enhancement at 3 WAS, 35.80% at 5 WAS and 30.33% enhancement at harvest stage, respectively, compared to the control (Table 2). Chlorophyll content was highest in T4 (RPAN8) treatment at both 2 WAS (0.48 mg g-1) and harvest stage (0.67 mg g-1) respectively (Table 2). Among the cyanobacterial treatments evaluated at 3 WAS, the highest chlorophyll values of 0.56 mg g-1 was observed in T2 (C11), followed by 0.51 mg g-1 in RPAN8. The soil chlorophyll content in all the treatments at 5 WAS varied from 0.24 to 0.63 mg g-1, with all the cyanobacterial treatments (C11, BF2 and RPAN8) exhibiting the highest values (0.63, 0.53 and 0.52 mg g-1).
Analysis of soil and plant nutrient attributes
The organic carbon content in soil samples ranged from 0.25 to 0.38 %, with all the cyanobacterial amendments (C11, BF2 and RPAN8) showing significantly higher values (0.38, 0.37 and 0.35%) than the control. At 2, 5 WAS and harvest stage, T2 (C11) treatment showed an increase of 52 %, 70.83% and 37.93% increase in terms of organic carbon content, in contrast to a 32%, 45.8% and 27.58 % increment caused by RPAN8 treatment (Fig. 2a). All the cyanobacterial treatments exhibited significant differences in available soil nitrogen content; soil under C11, BF2 and RPAN8 treatments experienced significantly higher available soil nitrogen content at both 2 and 5 WAS compared to the control (Fig. 2b). However, no significant differences in available nitrogen in soil was recorded at harvest stage, although C11 showed higher values. Among the treatments at 3 WAS, T4 (RPAN8) showed an 8.72% increase as compared to control (Fig. 2b). The available phosphorus content in soil at 3 WAS ranged from 6.82.2 – 11.95.2 mg kg-1; among the treatments, T2 (C11) showed higher total phosphorus content of 11.19 mg kg-1 compared to T1 (control). At 5 WAS, available phosphorus content was significantly higher in all the cyanobacterial treatments (C11, BF2 and RPAN8 -11.31, 11.27 and 11.37 mg kg-1) compared to the control (7.10.53 mg kg-1) respectively. Among the cyanobacterial treatments, the highest available phosphorus content of 12.12.07 mg kg-1 was observed in RPAN8 (Fig. 2c) at harvest stage. However, at 2WAS, no significant differences were observed among the cyanobacterial treatments.

Modulation of soil macronutrient levels as influenced by cyanobacterial treatments at different stages of growth (a) soil organic carbon, (b) available soil nitrogen, (c) available soil phosphorus. Error bars represent standard deviation of mean of three replicates. Abbreviations for treatments and attributes: T1: Control; T2:C11 (Anabaena laxa), T3: BF2 (Nostoc carneum), T4: RPAN8 (Anabaena laxa); WAS: weeks after sowing
The effect of cyanobacterial treatments (Table 3), in terms of available soil iron revealed that in all treatments (C11, BF2, RPAN8), excluding control, the highest values were recorded at 3 WAS, at 5 WAS and at harvest stage. However, no significant differences were recorded at 2 WAS. The available zinc content in soil showed significantly higher values, with a 1.22-fold and 1.20- fold increments in T4 (RPAN8) and T3 (BF2) respectively. However, all cyanobacterial treatments (C11, BF2, RPAN8), showed the highest increase of 74.33 %, 79.64% and 76.10 % at 3 WAS, (56.37 %, 64.42 % and 52.34 % increase) at 5 WAS and (34.65%, 46.59% and 36.93% increments) at harvest stage as compared to the control, respectively (Table 3). Interestingly, nitrogen content in leaves at 2 WAS exhibited significantly higher values in all the cyanobacterial treatments (C11, BF2 and RPAN8 - 3.67, 3.90 and 3.80 %), compared to control (3.40%) respectively (Table 3), with highest values recorded at harvest stage. C11 (4.18%) recorded significantly higher leaf nitrogen content at 3WAS, but at 5 WAS, RPAN8 recorded statistically at par values (Table 3). Comparison across cyanobacterial treatments at harvest stage showed significant differences (4.58 and 4.52%), with T2 (C11) and T4 (RPAN8) being the top performers. Analyses of leaf calcium content in cyanobacterial treatments illustrated that RPAN8 brought about an increase of 6.33%, while C11 and BF2 exhibited a 5.73 and 5.37% increment over control (Table 3).
Yield attributes
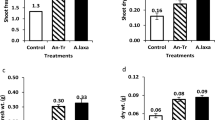
Cyanobacterial treatment resulted in a 54.35 % increase of leaf area, over the uninoculated control estimated at harvest stage (Table 4). Comparatively, the treatments C11 (35.92% increase) and RPAN8 (11.65% increase) showed better yield over control (Fig. 3). In terms of individual leaf weight, RPAN8 showed a 1.24-fold increase, followed by C11 exhibiting a 0.82-fold enhancement, respectively (Table 4). All the treatments with cyanobacteria significantly improved plant height, when compared with T1 (control) (Table 4). The root length of plants treated with C11 (14.96 cm) was higher than the control, and the number of branches were more in RPAN8 (12) by 1.19% (Table 4). The number of leaves was highest in T4 (RPAN8; 14), followed by T2 and T3, which were statistically at par (Table 4).

Influence of cyanobacterial treatments on yield and quality traits of spinach at harvest stage. Error bars represent standard deviation of mean of three replicates. Abbreviations for treatments and attributes: T1: Control; T2:C11 (Anabaena laxa), T3: BF2 (Nostoc carneum), T4: RPAN8 (Anabaena laxa)
Quality of the produce
The leaves of Pusa All Green analyzed for ascorbic acid content at harvest stage showed higher values for all cyanobacterial respectively (Fig. 3). Total soluble solids also exhibited a similar trend with C11, BF2 and RPAN8 recording an increase of 14.45, 13.98 and 16.31%, over control, respectively (Table 5). Phenol content in the leaves showed a wide range and the highest values of 391.17 mg GAE (100 g)-1 was recorded in RPAN8 treatment, which reflected 86.29% enhancement respectively (Table 5). In general, the titrable acidity content was not significantly modified by the cyanobacterial treatments (Table 5). As compared to control (uninoculated), the antioxidant activity in plants was significantly higher (1.51 µmol Trolox (100 g)-1) by the inoculation of either C11 or BF2 (Table 5). Analyses of protein content in leaves illustrated that cyanobacterial treatment (RPAN8) enhanced the content to the greatest extent, by 86.27%, followed by C11, which also brought about a significant increase in protein content in leaves (7.87 mg g-1 leaves) (Table 5). BF2 increased the content of β-carotene (Fig. 3), Vitamin A and carotenoids (Table 5) and brought about significant enhancement over the control (T1). The textural analysis in terms of firmness of spinach leaves (also referred to as film burst) revealed that C11 treatment (seed coating + soil drench) showed a 30.18% increase over Control (Table 5).This was followed by BF2, which brought about a significant increase of 17.15%.
Phenotypical correlation of plant, soil, yield and quality traits at different stages of growth
To investigate the associations among plant, soil, yield and quality traits, Pearson’s correlation coefficients were computed. A strong positive and significant association was observed between the soil nitrogen with plant/soil parameters and the values in ascending order are as follows: glutamine synthetase (GS) activity (r=0.85**) at 2 WAS (Fig. S2a) < available soil phosphorus (P) at 5 WAS (r=0.86**; Fig. S2c) < glutamine synthetase activity (GS) & organic carbon (OC)/ soil chlorophyll (SC; r=0.87**) at 5, 2 WAS, respectively. The nitrogen content in leaves showed a significant positive correlation with soil parameters such as soil chlorophyll (SC; r=0.934**) at 3 WAS (Fig. S2b; Fig. S5) and available soil phosphorus (P; r=0.865**) at 3 WAS respectively (Fig. S4a). The available iron content in soil exhibited positive correlation with chlorophyll a (r=0.936**) at harvest stage respectively (Fig. S2d) and with soil parameters like soil microbial biomass carbon (SMBC; r=0.980**) at harvest stage, soil chlorophyll (r=0.942** at harvest stage) and Available soil phosphorus (P; r=0.953**) at harvest stage, respectively. The phosphorus content in soil exhibited significant positive association with chlorophyll a (0.946**) at harvest stage, GS (r=0.910**) at harvest stage, SMBC (r=0.980**) at harvest stage, soil dehydrogenase (r=0.904**) at 3 WAS (Table S.3) and soil chlorophyll (r=0.966; 0.954**) at 3 WAS and harvest stage respectively. A positive and significant association was observed between the OC with GS (r=0.950;0.909/0.971**) at 2, 5 WAS (Fig. S6) and harvest stage respectively (Fig. S3d) and with soil parameters like SMBC (r=0.901**) at harvest stage, soil dehydrogenase (r=0.919**) at 2 WAS (Fig. S4) and soil chlorophyll(r=0.924**) at 3 WAS respectively (Fig. S5). The chlorophyll content in soil exhibited significant positive correlation with phosphoenol pyruvate carboxylase activity (PEPCO; r = 0.904**) at 2 WAS (Table S.1) and SMBC (r = 0.952**) at harvest stage respectively. A positive and significant association was observed between dehydrogenase activity in soil with traits such as chlorophyll a (r= 0.941;0.903**) at 2 WAS and harvest stage, carotenoids (r= 0.911*) at 2 WAS, total pigments (r= 0.930;0.895**) at 2, 3 WAS (Table S.1), and GS (r= 0.913;0.934**) at 2 WAS and harvest stage and with SMBC (r=0.893**) at 2 WAS respectively. SMBC exhibited significant positive correlation with plant traits like chlorophyll a (r=0.923**) at harvest stage (Fig. S7), nitrate reductase (r=0.902;0.945/0.983**) at 2, 3, 5 WAS (Table S5); and GS (r=0.914**) at harvest stage respectively. Nitrate reductase activity showed a significantly positive correlation with chlorophyll a, chlorophyll b, carotenoids and total pigments content (r=0.910**; 0.821*; 0.713*; 0.899**) at 2 WAS and (r=0.772*; 0.845**; 0.822*;0 .816*) at harvest stage respectively (Table S.7).
Among yield variables recorded at harvest stage (Fig. 4a; Table S9), the leaf calcium content showed a strong significant positive correlation with parameters and the values were in ascending order are as follows: organic carbon (r=0.827**) <Chlorophyll a (CHLA; r=0.841**) < Soil Microbial Biomass Carbon (r=0.877**) < available soil phosphorus (r=0.881**) < available iron (r=0.887**) < soil chlorophyll (r=0.898**) at harvest stage (Fig. S8). There were higher correlations observed among individual plant weight/nitrate reductase activity and individual plant weight/Leaf nitrogen (r=0.933**) and (r=0.827**) at harvest stage respectively. However, there were also significant higher correlations observed between vitamin A with phosphoenol pyruvate carboxylase activity (r=0.912**) respectively (Fig. 4b; Fig. S9; Table S11). The leaf protein content showed a strong significant positive correlation with the values in ascending order: chlorophyll a, glutamine synthetase, organic carbon (r=0.84**) < soil available iron (r=0.90**) <available phosphorus (r=0.94**) < SMBC/SC (r=0.95**). Most interestingly, the ascorbic acid trait depicted a strong positive correlation with soil chlorophyll (r=0.890**), SMBC (r=0.850**) and soil available iron (r=0.851**).

Phenotypical Correlation network, depicting the relationships among plant, soil, yield and quality traits (a) yield traits with plant and soil parameters at harvest stage; (b) quality traits with plant and soil at harvest stage; P: available soil phosphorus; OC: soil organic carbon; SC: soil chlorophyll; DHY: soil dehydrogenase activity; SMBC: soil microbial biomass carbon; PEPC: Phosphoenol pyruvate carboxylase activity; GS: glutamine synthetase activity; NR: nitrate reductase activity; TP: total pigments; CAR: carotenoids; CHLB: chlorophyll b; LN: leaf nitrogen; SN: available soil nitrogen; LCA: leaf calcium; LA: leaf area; NOL: number of leaves; NOB: number of branches; PW: plant weight; IPW: individual plant weight; RL: root length; PH: plant height; CHLA: chlorophyll a; ZN: soil available zinc; FE: soil available iron; VITA: vitamin A; LP: leaf proteins; PHE: phenols; AOX: antioxidants; CAR: carotene; BCAR: beta carotene; FB: film burst; TSS: total soluble solids; TA: titrable acidity; AA: ascorbic acid
Path coefficient analysis
Significant effects were recorded for path coefficient analyses of available soil nitrogen with other parameters at 2, 3, 5 WAS and harvest stage. Positive direct effects of PEPCO (2.66), carotenoids (2.19), chlorophyll a (1.40), soil dehydrogenase activity (1.32) and Glutamine synthetase activity (1.27) besides organic carbon (0.13) and Soil Microbial Biomass Carbon (0.01) on available soil nitrogen at 2 WAS were recorded (Fig. 5a). The residual effect was 0.001, which clearly depicted that all the parameters at 2 WAS were majorly involved in enhancing the soil available nitrogen, brought about by cyanobacterial treatments (Fig. S3a; Table S2). The positive direct effect of nitrate reductase activity (10.64) and chlorophyll b (10.23) on available soil nitrogen was found to be very high at 3 WAS are observed (Table S4). The residual effect was 0.002, which shows that the increase in soil available nitrogen is due to the other attributes at 3 WAS investigated, as influenced by cyanobacterial treatments (Fig. S3b). On the other hand, chlorophyll a (7.17) in the leaves exerted a major direct effect on available soil nitrogen values at 5 WAS in inoculated treatments (Fig. S3c). Furthermore, the direct positive effect of available soil phosphorus in the soil (5.28) carotenoids (3.39) and PEPCO (3.15) was found to be stronger in the inoculation treatments (Table S.6). At harvest stage, carotenoids (3.27), soil available iron (2.83) and chlorophyll a (2.65) had positive direct effects on available soil nitrogen at harvest stage followed by glutamine synthetase activity (1.78), available soil Zn (0.98), soil dehydrogenase activity (0.53) and soil chlorophyll (0.48). The residual value for the path analysis model was 0.001 at harvest stage was also observed (Fig. S3d; Table S8). Except for total soluble solids (8.02) and leaf firmness/ film burst (9.26); the assessed traits had positive direct effects on available soil nitrogen through quality traits, including antioxidant activity (25.82), phenols (24.96), leaf proteins (21.73), beta carotene (14.89) and titrable acidity (10.74). The residual value for the path analysis model was 0.003 at harvest stage was observed for quality traits with soil available nitrogen (Fig. 5; Table S12) whereas with yield traits; Leaf area (3.99) and Individual plant weight (1.42) had the greatest positive direct effect on soil available nitrogen (Fig. S3e; Table S10).

Network path diagram depicting the association of soil available nitrogen with quality traits at harvest stage; PEPCO: Phosphoenol pyruvate carboxylase activity
Discussion
Cyanobacteria are a valuable bioresource which play a significant role in the sustainability of various sectors, particularly agriculture and industry (Rana et al. 2012; Gupta et al. 2013; Grzesik and Romanovska-Duda 2015; Kholssi et al. 2021; Shivay et al. 2022). They are valuable as biofertilizers or nutrient-mobilizing inoculants, which have been successfully utilized in various crops, including rice, wheat, maize, tomatoes, and other vegetables (Kumar et al. 2013; Prasanna et al. 2014, 2021). Several cyanobacteria are known to enhance yields in various vegetable and fruit species as documented in several studies (Geries and Elsadany 2021; Kokila et al. 2023; Bhardwaj et al. 2024), however, little research is documented on cyanobacteria as an input to enhance the commercial and qualitative attributes of spinach as a crop. In the current study, the use of carrier-based cyanobacterial formulations, as seed coating, specifically those amended with A. laxa C11, N. carneum BF2 or A. laxa RPAN8, along with their use as soil drench, in the cultivation of Pusa All Green variety of spinach in shaded net house was investigated. A range of growth-related, enzymatic, soil biological, yield, and quality parameters as indicators were evaluated the role of cyanobacteria as an organic intervention towards the production of a healthy spinach crop.
Dehydrogenase activity, microbial biomass carbon, and soil chlorophyll content serve as important soil microbiological indicators to gauge nutrient mobilization and soil quality indices as they correlate positively with the abundance of microbial numbers (Filip 2002; Hanaka et al. 2019) Soil microbial biomass plays a fundamental role in regulating the transformation processes of elements, besides governing the accumulation and decomposition of organic matter, and serving as an early indicator of perturbations due to changes in agronomic practices or contamination through xenobiotics or heavy metals. The soil microbial biomass carbon in cyanobacteria-inoculated treatments was enhanced by 2-3 fold over control, underscoring the significant impact of cyanobacterial inoculation on soil quality. Several previous studies showed an increase in microbial biomass carbon by 15-30%, highlighting the remarkable ability of cyanobacteria to substantially contribute towards soil carbon accretion (Prasanna et al. 2014, 2021; Kokila et al. 2023). Photosynthetic prokaryotes, such as cyanobacteria have the capability to capture and fix atmospheric CO2 leading to an anticipated increase in soil carbon with cyanobacterial treatment (Renuka et al. 2017), This may also lead to proliferation of soil microbial populations (Mager 2010), as recorded in cyanobacterial soil crusts of Kalahari sand, which contained large fractions of soil organic pools with carbohydrates as their major constituent. In addition, the proliferation of microalgae and cyanobacteria is known to be a source of organic matter for the growth of higher plants (Renuka et al. 2018). As an indirect impact, the decaying biomass of microalgae and cyanobacteria in agricultural fields can be decomposed or utilized by various other organisms and grazers, thereby augment the organic carbon levels in the soil. The significant role of cyanobacteria in paddy fields, besides nitrogen fixation was emphasized by Mandal et al. (1999) as they can trigger enzymatic processes in soil and plants, and promote fertility and plant health, as also observed in this study. The overall changes in all the microbiological and plant parameters recorded in the present investigation depict a beneficial relationship between cyanobacterial treatment and the tested spinach genotype, duly supported by previous studies in which soil microbiological activity, such as that of dehydrogenase or those related to nutrient mobilization were greatly enhanced upon the addition of cyanobacterial biomass and their exopolysaccharides (De Caire et al. 2000; Rana et al. 2012; Prasanna et al. 2018). This has often been attributed to the beneficial impact of cyanobacterial inoculation on plant growth and development in terms of increased availability of nitrogen in the soil, microbial biomass carbon accretion, particularly photosynthetic biomass, measured as soil chlorophyll. Bergi and Trivedi (2020) discussed the role of cyanobacteria as biofertilizers in increasing phosphorus content in saline soils, while in our study, significant improvement in nitrogen content in spinach leaves was recorded. This aligns with prior research on the use of cyanobacterial biofertilizers, which as diazotrophs increase nitrogen availability, followed by its mobilization, enhancing plant nitrogen concentration, and promoting plant growth during the growth stage, as reported by Rodgers et al. (1979).
Similarly, IAA-producing cyanobacterial strains caused significant enhancements in both soil and plant chlorophyll content in wheat seedlings (Sood et al. 2011); earlier also reported by PGPR by Cakmakci et al. (2009). Bharti et al. (2021), amended the potting medium with a cyanobacterial strain and found significant increase in nitrogen availability in the soil, resulting in improved flower traits and productivity in Chrysanthemum. Therefore, coating seeds with cyanobacterial formulations or its use as a drench can be instrumental in enhancing nutrient availability in the soil surrounding spinach plants, leading to improved growth and overall productivity of the crop. Chlorophyll molecules play a pivotal role in the light-dependent photosynthetic reactions by capturing light energy and transferring this absorbed energy to the photo system reaction centers in plants and algae (Ghotbi et al. 2019). Spinach, being valued for its culinary and nutritional attributes of its leaves, our effort was to enrich this trait and we observed a positive response to cyanobacterial inoculation. A significant increase in The levels of chlorophyll a, b, carotenoids, and total pigments was recorded in the treated plants as compared to the control, as earlier recorded by Grzesik and Romanovska (2015), in willows treated with Microcystis aeruginosa and Anabaena sp. An increased chlorophyll content can be attributed due to the presence of microelements (iron zinc and magnesium) being more available, as earlier also shown in rice and wheat plants receiving cyanobacterial inoculation (Rana et al. 2012; Shivay et al. 2022).
Cyanobacteria have garnered attention for their potential to enhance soil macro- and micronutrient availability and aid in the efficient mobilization of minerals by plants (Singh et al. 2016; Renuka et al. 2018). Furthermore, the cyanobacterial strains utilized in our study enhanced soil micronutrient parameters, including available iron and available zinc in soil, as also recorded earlier (Rana et al. 2012; Shivay et al. 2022). Manjunath et al. (2016) observed that soil available iron levels at the mid-crop stage recorded a threefold increase compared to the control when An–Az biofilm formulation was used in okra crop. Renuka et al. (2017) evaluated the promise of unicellular and filamentous micro algal-cyanobacterial consortial inoculation on soil composition and grain quality in a wheat crop. A significant increase in micronutrient availability in the soil, particularly zinc and iron, soil organic carbon content, led to improved crop yield, and enhanced grain quality.
Earlier economic analyses of spinach production undertaken illustrated that cultivating spinach under green shade net conditions showed promise, resulting in a substantial increase in yields and productivity (Meena et al. 2014). The cyanobacterial treatments resulted in significant changes in plant height, root length, increase in yield, individual leaf weight, and expansion in leaf area compared to the control (T1) plants. These findings align with previous observations (Salamah et al. 2019), in which the use of Nostoc sp. SO-A31 as a nitrogen source resulted in robust vegetative growth and increased biomass production in spinach. Similarly, the use of Chlorella vulgaris and Spirulina platensis in a pot experiment with maize, led to increase in the number of leaves from 6 in the control plants to 8 leaves in the treated plants (Dineshkumar et al. 2019). In a study conducted by Kumar et al. (2013), the application of cyanobacterial strains, A. laxa and Calothrix elenkinii as seed treatment of spice crops led to significant improvement in the plant biometrical parameters and dry weight. Kholsi et al. (2021) examined the combined use of cyanobacteria, especially Calothrix sp. and Anabaena cylindrica, and plant-growth-promoting rhizobacteria (PGPR) and recorded more than 36% increase in plant height in wheat. Similar improvement in plant biometric parameters, including leaf area, and yield were recorded in the present investigation.
Significant enrichment to the quality attributes of the Pusa All green spinach cultivar using three different cyanobacterial treatments was observed. Specifically, an increase in phenol content, leaf proteins, along with boost in beta carotene content, and enhancement in leaf firmness. The most promising treatment RPAN8 was also found to elevate antioxidant activity significantly. Previous studies (Mala et al. 2017; Ertani et al. 2019) have demonstrated the effectiveness of cyanobacterial-based biostimulants, as soil inoculation or foliar application against oxidative stress. This is known to help in minimizing the damage caused by stress originated free radicals, by acting as scavengers, preventing the production of reactive oxygen species. Singh et al. (2011) documented that introducing Oscillatoria acuta and Plectonema boryanum through soil inoculation resulted in a notable enhancement of total phenolic content in rice. Mahmoud et al. (2019) demonstrated that the spinach leaves, when applied with an aqueous extract from Anabaena sphaerica increased the plant protein content.
Cyanobacterial inoculation can improve the micronutrient density of the produce, as shown in in rice, wheat and maize grains/seeds (Rana et al. 2012; Nishanth et al. 2021; Shivay et al. 2022). Kokila et al. (2023) recorded enhanced fruit quality in tomato, with higher titrable acidity, ascorbic acid, carotenoids and lycopene content in the tomato fruits. This is known to be mediated through siderophores or other chelating molecules and their respective transporters. The accumulation of phenolic acids, including caffeic, chlorogenic, ferulic, gallic, and gentisic acids, as well as increased levels of proteins and chlorophylls was shown in rice plants inoculated with various cyanobacterial strains (Singh et al. 2011). These findings support the beneficial effects of cyanobacteria as biofertilizers and effective biostimulatory agents for promoting plant growth and quality of produce, as also shown in spinach, in this investigation.
Phenotypic correlation analyses aligned with the biochemical and microbiological data recorded in spinach crop and brought to light the significant role played by cyanobacteria-based seed coating and drenching in fostering positive connections among a range of plant, soil, yield and quality factors. These results underscore the valuable contributions of diazotrophy, IAA and macro- and micronutrient mobilizing potential of cyanobacteria in stimulating the robust growth of spinach crop. The correlation analyses spotlighted the strong associations existing between soil available nitrogen, yield and quality traits with cyanobacterial inoculants at various stages of growth. Utilization of these cyanobacterial formulations, specifically A. laxa, led to a substantial impact on a multitude of traits linked to plant, soil, yield and quality parameters.
Path coefficient analysis has conventionally been applied in subjects such as plant breeding, varietal selection and yield analyses. However, it has been relatively overlooked in the realm of microbiology, particularly, plant-microbe interactions. By using this analysis, the findings of this investigation were analysed through an innovative perspective, which provided insights regarding the substantial impact of cyanobacterial treatments in enhancing the plant growth, soil characteristics, yield as well as quality attributes in spinach crop. Path coefficient analyses was instrumental in elucidating the vital role of examined parameters, particularly in the context of improving plant health, as evidenced by the important role of soil available nitrogen with yield and quality traits at 2, 3, 5 WAS and harvest stage.
This study is noteworthy in that it resulted in a nutrimental intervention, through seed coating + drench using cyanobacterial formulations to reduce chemical inputs for a crop, particularly urea (N fertilizer; only one basal dose of urea, as against two in control, given in cyanobacterial treatments). This is often a costly input, which is environment-polluting and in excess, often a wasteful expenditure, due to its volatilization. Such a research mediation in spinach cultivation with better quality of produce, which is mostly consumed fresh, showcases microbial inputs as a more efficient and cost-effective approach. Further research is focused on evaluation across diverse agro-climatic conditions and varieties, to assess the viability and efficacy, thereby environmental proofing the technology and its widespread use in agriculture.
Data availability
All datasets generated or analysed during the current study are given as Tables and Figures as well as Supplementary Tables and Figures.
References
Apak R, Güçlü K, Özyürek M, Karademir SE (2004) Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J Agric Food Chem 52:7970–7981
Arnon DI (1949) Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol 24:1–15
Association of Official Analytical Chemists [AOAC] (2000) Coffee and tea, Official methods of analysis, 17th edn. AOAC, Gaithersburg
Bergi J, Trivedi R (2020) Bioremediation of saline soil by cyanobacteria. In Microb Bioremed Biodegrad, p 447–465
Bhardwaj A, Prasanna R, Bavana N, Kokila V, Nivedha RM, Gaur Rudra S, Singh AK, Shivay YS (2024) Foliar application of cyanobacterial formulations stimulates plant growth and fruit quality in tomato under protected cultivation. N Z J Crop Hort Sci. https://doi.org/10.1080/01140671.2023.2294729
Bharti A, Prasanna R, Kumar G, Nain L, Rana A, Ramakrishnan B, Shivay YS (2021) Cyanobacterium-primed Chrysanthemum nursery improves performance of the plant and soil quality. Biol Fertil Soils 57:89–105
Cakmakci R, Erat M, Oral B, Erdogan Ü, Șahin F (2009) Enzyme activities and growth promotion of spinach by indole-3-acetic acid-producing rhizobacteria. J Hort Sci Biotechnol 84:375–380
Carillo P, Ciarmiello LF, Woodrow P, Corrado G, Chiaiese P, Rouphael Y (2020) Enhancing sustainability by improving plant salt tolerance through macro-and micro-algal biostimulants. Biology 9:253
Casida LE Jr, Klein DA, Santoro T (1964) Soil dehydrogenase activity. Soil Sci 98:371–376
De Caire GZ, De Cano MS, Palma RM, De Mule CZ (2000) Changes in soil enzyme activities following additions of cyanobacterial biomass and exopolysaccharide. Soil Biol Biochem 32:1985–1987
Dineshkumar R, Subramanian J, Gopalsamy J, Jayasingam P, Arumugam A, Kannadasan S, Sampathkumar P (2019) The impact of using microalgae as biofertilizer in maize (Zea mays L.). Waste Biomass Valoriz 10:1101–1110
DuBois M, Gilles KA, Hamilton JK, Rebers PT, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356
Ertani A, Nardi S, Francioso O, Sanchez-Cortes S, Foggia MD, Schiavon M (2019) Effects of two protein hydrolysates obtained from chickpea (Cicer arietinum L.) and Spirulina platensis on Zea mays (L.) plants. Front Plant Sci 10:954
Fallik E, Alkalai-Tuvia S, Parselan Y, Aharon Z, Elmann A, Offir Y, Matan E, Yehezkel H, Ratner K, Zur N, Shahak Y (2008) Can colored shade nets maintain sweet pepper quality during storage and marketing? ISHS Acta Horticulturae 830:37–44
Filip Z (2002) International approach to assessing soil quality by ecologically-related biological parameters. Agric Ecosyst Environ 88:169–174
Garlapati D, Chandrasekaran M, Devanesan A, Mathimani T, Pugazhendhi A (2019) Role of cyanobacteria in agricultural and industrial sectors: an outlook on economically important byproducts. Appl Microbiol Biotechnol 103:4709–4721
Geries LS, Elsadany AY (2021) Maximizing growth and productivity of onion (Allium cepa L.) by Spirulina platensis extract and nitrogen-fixing endophyte Pseudomonas stutzeri. Arch Microbiol 203:169–181
Ghotbi-Ravandi AA, Shariati M, Shobbar ZS, Shahbazi M (2019) Expression pattern and physiological roles of plastid terminal oxidase (PTOX) in wild and cultivated barley genotypes under drought stress. Environ Exp Bot 162:313–320
Gopalan C (1992) New dimensions of ‘old problem’. In: Nutrition in Development Transition in South-East Asia. World Health Organisation, Regional Office for South-East Asia, New Delhi. pp 34–48
Anjana, Umar S, Iqbal M, Abrol YP (2007) Are nitrate concentrations in leafy vegetables within safe limits?. Curr Sci 92:355–360
Gopinath PP, Parsad R, Joseph B, Adarsh VS (2020) GRAPES: General R shiny Based Analysis Platform Empowered by Statistics https://www.kaugrapes.com/home
Gordon SA, Weber RP (1951) Colorimetric estimation of indoleacetic acid. Plant Physiol 26:192–195
Grzesik M, Romanowska-Duda Z (2015) Ability of cyanobacteria and green algae to improve metabolic activity and development of willow plants. Pol J Environ Stud 24:1003–1012
Gupta V, Ratha SK, Sood A, Chaudhary V, Prasanna R (2013) New insights into the biodiversity and applications of cyanobacteria (blue-green algae)—Prospects and challenges. Algal Res 2:79–97
Hanaka A, Plak A, Zagórski P, Ozimek E, Rysiak A, Majewska M, Jaroszuk-Ściseł J (2019) Relationships between the properties of Spitsbergen soil, number and biodiversity of rhizosphere microorganisms, and heavy metal concentration in selected plant species. Plant Soil 436:49–69
Herbert D, Phipps PJ, Strange RE (1971) Chemical analysis of microbial cells. Meth Microbiol 5:209–344
Higa T, Wididana GN (1991) The concept and theories of effective microorganisms. In: Proceedings of the first international conference on Kyusei nature farming. US Department of Agriculture, Washington, DC pp 118–124
Kholssi R, Marks EA, Miñón J, Maté AP, Sacristán G, Montero O, Debdoubi A, Rad C (2021) A consortium of cyanobacteria and plant growth promoting rhizobacteria for wheat growth improvement in a hydroponic system. S Afr J Bot 142:247–258
Kittas C, Rigakis N, Katsoulas N, Bartzanas T (2008) Influence of shading screens on microclimate, growth and productivity of tomato. ISHS Acta Horticulturae 807:97–102
Kokila V, Prasanna R, Kumar A, Nishanth S, Singh B, Rudra SG, Pal P, Pal M, Shivay YS, Singh AK (2023) Elevated CO2 along with inoculation of cyanobacterial biofilm or its partners differentially modulates C-N metabolism and quality of tomato beneficially. Heliyon 9:e20470
Kumar M, Prasanna R, Bidyarani N, Babu S, Mishra BK, Kumar A, Adak A, Jauhari S, Yadav K, Singh R, Saxena AK (2013) Evaluating the plant growth promoting ability of thermotolerant bacteria and cyanobacteria and their interactions with seed spice crops. Sci Hortic 164:94–101
Lichtenthaler HK (1987) [34] Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth Enzymol 148:350–382
Mager DM (2010) Carbohydrates in cyanobacterial soil crusts as a source of carbon in the southwest Kalahari. Botswana. Soil Biol Biochem 42:313–318
Mahmoud SA, Abd El-Aty AM, Kandil H, Siam HS (2019) Influence of different algal species application on growth of spinach plant (Spinacia oleracea L.) and their role in phytoremediation of heavy metals from polluted soil. Plant Arch 19:2275–2281
Mala R, Celsia AR, Mahalakshmi R, Rajeswari S (2017) Agronomic biofortification of Amaranthus dubius with macro nutrients and vitamin A. IOP Conf Ser: Mater Sci Eng 225:012214
Mandal B, Vlek PL, Mandal LN (1999) Beneficial effects of blue-green algae and Azolla, excluding supplying nitrogen, on wetland rice fields: a review. Biol Fertil Soils 28:329–342
Manjunath M, Kanchan A, Ranjan K, Venkatachalam S, Prasanna R, Ramakrishnan B, Hossain F, Nain L, Shivay YS, Rai AB, Singh B (2016) Beneficial cyanobacteria and eubacteria synergistically enhance bioavailability of soil nutrients and yield of okra. Heliyon 2:e00066
Meena RK, Vashisth A, Singh R, Singh B, Manjaih KM (2014) Study on change in microenvironment under different colour shade nets and its impact on yield of spinach (Spinacia oleracea L.). J Agrometeorol 16:104–111
Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31:426–428
Miller JG, Buchanan CJ, Eastwood MA, Fry SC (1995) The solubilisation and hydrolysis of spinach cell wall polysaccharides in gastric and pancreatic fluids. J Sci Food Agric 68:389–394
Nayak S, Prasanna R, Pabby A, Dominic TK, Singh PK (2004) Effect of urea, blue green algae and Azolla on nitrogen fixation and chlorophyll accumulation in soil under rice. Biol Fertil Soils 40:67–72
Nishanth S, Prasanna R, Hossain F, Muthusamy V, Shivay YS, Nain L (2021) Interactions of microbial inoculants with soil and plant attributes for enhancing Fe and Zn biofortification in maize genotypes. Rhizosphere 19:100421
Nunan N, Morgan MA, Herlihy M (1998) Ultraviolet absorbance (280 nm) of compounds released from soil during chloroform fumigation as an estimate of the microbial biomass. Soil Biol Biochem 30:1599–1603
Olsen SR (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. US Department of Agriculture Circular 939 p 19
Prakash D, Pal M (1991) Nutritional and antinutritional composition of vegetable and grain amaranth leaves. J Sci Food Agric 57:573–583
Prasad R (2006) Zinc in soils and in plant, human & animal nutrition. Indian J Fertil 2:103-119
Prasad R, Shivay YS, Kumar D, Sharma SN (2006) Learning by doing exercises in soil fertility (A Practical Manual for Soil Fertility). Division of Agronomy, Indian Agricultural Research Institute, New Delhi, p 68
Prasanna R, Renuka N, Nain L, Ramakrishnan B (2021) Natural and constructed cyanobacteria-based consortia for enhancing crop growth and soil fertility. In: Seneviratne G, Zavahir JS (eds) Microbes for sustainability: Role of microbial communities. Springer, Cham, pp 333–362
Prasanna R, Saxena G, Singh B, Ranjan K, Buddhadeo R, Velmourougane K, Ramakrishnan B, Nain L, Singh MC, Hasan M, Shivay YS (2018) Mode of application influences the biofertilizing efficacy of cyanobacterial biofilm formulations in chrysanthemum varieties under protected cultivation. Open Agric 3:478–489
Prasanna R, Sood A, Ratha SK, Singh P (2014) Cyanobacteria as a “green option” for sustainable agriculture. In: Sharma NK, Stal LJ, Rai AK (eds). Cyanobacteria: An Economic Perspective. John Wiley and Sons, Chichester, pp 145–166
Raghuramulu N, Nair KM, Kalyanasundaram S (eds) (1983) A manual of laboratory techniques. National Institute of Nutrition, Hyderabad
Rana A, Joshi M, Prasanna R, Shivay YS, Nain L (2012) Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur J Soil Biol 50:118–126
Ranganna S (1986) Handbook of analysis and quality control for fruit and vegetable products. Tata McGraw-Hill Education, New Delhi
Renuka N, Guldhe A, Prasanna R, Singh P, Bux F (2018) Microalgae as multi-functional options in modern agriculture: current trends, prospects and challenges. Biotechnol Adv 36:1255–1273
Renuka N, Prasanna R, Sood A, Bansal R, Bidyarani N, Singh R, Shivay YS, Nain L, Ahluwalia AS (2017) Wastewater grown microalgal biomass as inoculants for improving micronutrient availability in wheat. Rhizosphere 3:150–159
Rodgers GA, Bergman B, Henriksson E, Udris M (1979) Utilisation of blue-green algae as biofertilisers. Plant Soil 52:99–107
Salamah A, Fadilah N, Khoiriyah I, Hendrayanti D (2019) Application of N2-Fixing cyanobacteria Nostoc sp. SO-A31 to hydroponically grown water spinach (Ipomoea aquatic L.). AGRIVITA J Ag Sci 41:325–334
Shapiro BM, Stadtman ER (1970) [130] Glutamine synthetase (Escherichia coli). Meth Enzymol 17:910–922
Shivay YS, Prasanna R, Mandi S, Kanchan A, Simranjit K, Nayak S, Baral K, Sirohi MP, Nain L (2022) Cyanobacterial inoculation enhances nutrient use efficiency and grain quality of basmati rice in the system of rice intensification. ACS Agric Sci Technol 2:742–753
Singh DP, Prabha R, Yandigeri MS, Arora DK (2011) Cyanobacteria-mediated phenylpropanoids and phytohormones in rice (Oryza sativa) enhance plant growth and stress tolerance. Antonie van Leeuwenhoek 100:557–568
Singh JS, Kumar A, Rai AN, Singh DP (2016) Cyanobacteria: a precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front Microbiol 7:529
Singh RK, Chaudhary BD (1977) Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, New Delhi
Singleton VL, Orthofer R, Lamuela-Raventós RM (1999) Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth Enzymol 299:152–178
Snell FD, Snell CT (1961) Colorimetric methods of analysis. D. van Nostrand, New York
Sood A, Singh PK, Kumar A, Singh R, Prasanna R (2011) Growth and biochemical characterization of associations between cyanobionts and wheat seedlings in co-culturing experiments. Biologia 66:104–110
Stanier RY, Kunisawa R, Mandel MC, Cohen-Bazire G (1971) Purification and properties of unicellular blue-green algae (order Chroococcales). Bact Rev 35:171–205
Subbiah BV, Asija GL (1956) A rapid procedure for the estimation of available nitrogen in soils. Curr Sci 25:259–260
Sun Q, Zhang N, Wang J, Zhang H, Li D, Shi J, Li R, Weeda S, Zhao B, Ren S, Guo YD (2015) Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J Exp Bot 66:657–668
Vázquez E, García-Risco MR, Jaime L, Reglero G, Fornari T (2013) Simultaneous extraction of rosemary and spinach leaves and its effect on the antioxidant activity of products. J Supercrit Fluids 82:138–145
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Wright SF, Upadhyaya A (1996) Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci 161:575–586
Wu MX, Wedding RT (1985) Diurnal regulation of phosphoenolpyruvate carboxylase from Crassula. Plant Physiol 77:667–675
Acknowledgements
The Division of Microbiology, ICAR-IARI, is gratefully acknowledged for providing the necessary facilities. Shri Gulab Singh is thanked for support in the nutrient analyses, and the facilities of the Division of Agronomy are gratefully acknowledged towards facilitating the nutrient analyses of soil samples. The staff of CPCT, particularly Shri Parveshji, deserve our gratitude for their support in the maintenance of spinach crop.
Funding
RMN and NRB thank the Graduate School, ICAR-Indian Agricultural Institute, New Delhi for the fellowship towards PhD and MSc programs, respectively. All the authors are grateful for the funds made available through the Division of Microbiology, ICAR-Indian Agricultural Institute, New Delhi and Indian Council of Agricultural Research (ICAR) Network Project on Microorganisms “Application of Microorganisms in Agricultural and Allied Sectors” (AMAAS).
Author information
Authors and Affiliations
Contributions
RMN: Investigation, Data curation, Writing- Original draft preparation, Software use and visualization; RP: Conceptualization, Methodology, Writing- Reviewing and Editing; AB, NB: Investigation, Data curation, SGR, SKL, SB: Resources, Methodology, Data curation; AKS: Investigation, Resources; YSS: Methodology, Data curation. All the authors approved the final version submitted
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nivedha, R.M., Prasanna, R., Bhardwaj, A. et al. Cyanobacteria-based nutrimental strategy to enhance yield and quality of spinach. J Appl Phycol (2024). https://doi.org/10.1007/s10811-024-03292-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10811-024-03292-0