
Abstract
This study aimed to evaluate and compare the intertidal macroalgae community from reef structures subject to different urbanization degrees. Samplings were made in 11 beaches from the Pernambuco coast, northeastern Brazil. The sites were classified according to the level of urbanization into three classes of increasing urbanization pressure: non urbanized (NU), in urbanization process (UP), and consolidated urbanization (UC). Macroalgae were identified in situ non-destructively. A total of 53 taxa were identified, 41 of these were macroalgae. Significant differences were observed in the composition of macroalgae according to the urbanization levels, with Palisada perforata, Gelidiella acerosa, and Caulerpa spp. dominating NU and UP sites, whereas Chondracanthus acicularis, Bryopsis sp., and Ulva spp. dominated UC sites. This work shows that urbanization can have a strong effect on the structure of rocky intertidal macroalgal assemblages and highlights some macroalgae species that can be used as bioindicators for assessing the impact of urbanization on coastal shores.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nearly a quarter of the Brazilian population lives on the coastal zone, being the Northeastern region one of the most densely occupied. This region is rich in consolidated substrates (Branner 1904; Laborel 1969; Dominguez et al. 1990; Castro and Pires 2001), supporting a rich benthic biodiversity. The reefs in urban areas are highly susceptible to anthropogenic impacts such as elevated sedimentation rates and nutrient run-off that can modify the structure and diversity of these ecosystems (Worm et al. 1999; Airoldi 2003; Connell et al. 2008; Krause-Jensen et al. 2008; Goatley and Bellwood 2013). These impacts generally lead to the establishment and dominance of opportunistic species (Borowitzka 1972), which may boost low levels of diversity.
Biodiversity of coastal ecosystems close to large urban centers is often affected by several anthropic pressures, like pollution (Johnston and Roberts 2009; Martins et al. 2012), overexploitation, habitat destruction, and climate change (Scherner et al. 2013) which, cumulatively, can lead to the degradation of these productive ecosystems (Shanmugam et al. 2007; Halpern et al. 2012; Turra et al. 2013). For instance, there is great concern about the widespread loss of canopy-forming algae from many rocky coastlines and how it may be related to urbanization (Benedetti-Cecchi et al. 2001; Airoldi and Beck 2007; Connell et al. 2008).
Macroalgae play a fundamental ecological role as primary producers, by providing sheltering, nursery grounds, and substrate for a diverse assemblage of species (Schiel 2006). Many algal species (i.e., canopy-forming algae) are commonly recognized as key for ecosystem engineers as they play an important structuring role in intertidal communities (Davison and Pearson 1996). Like many other organisms, however, macroalgae are exposed to several disturbances of both natural and human origins. Intertidal macroalgal assemblages generally respond rapidly to those disturbances, as they are regularly exposed to potential stressful conditions, such as nutrient limitation, temperature range, desiccation, osmotic pressure variations, and anthropic contaminants (Areces 2001; Scherner et al. 2013).
The effects of anthropogenic disturbances on the phycoflora can result in changes of dominance, specific composition of assemblages, and variations in the distribution patterns. Some macroalgal groups are particularly susceptible to anthropogenic disturbances. For instance, the canopy-forming Cystoseira spp. has been shown to decline and replaced by turf-forming species with increasing anthropogenic pressure along the Mediterranean (i.e., Benedetti-Cecchi et al. 2001). In tropical areas, canopies formed by macroalgae (Sargassum spp.) are also important, and their loss has led to long-term changes in structure and functioning of benthic communities, with consequences reaching far up the food chain (Tait and Schiel 2011). Thus, this study aimed to evaluate and compare the structure of intertidal reef macroalgal assemblages affected by different levels of coastal urbanization.
Material and methods
Study area
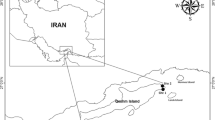
The Pernambuco coast comprises 187 km long (7°32′52″ S 34°50′27″ W and 8°54′57″ S, 35°09′76″ W), and it is located in the Tropical Southwest Atlantic biogeographic province and Brazil’s Northeastern ecoregion (Fig. 1) (Spalding et al. 2007). The local climate is classified as humid tropical (As′) (Köppen 1948), with average annual temperatures ranging between 25 °C (minimum) and 30 °C (maximum) (CPRH 2003). The region has a semidiurnal tidal regime, with a predominant E-SE swell, and an average superficial sea temperature between 27 and 29 °C (Manso et al. 2006). The Pernambuco coastal area represents only 4% of the state territory; however, it hosts 46% of the Pernambuco population, figuring the second most densely populated city of the Brazilian coast (IBGE 2010). The Pernambuco coastal reefs are basically rocky bodies composed mostly by quartz sand grains, cemented by calcium carbonate. These reef formations are abundant in the northeastern Brazilian coast (Branner 1904; Laborel 1969; Dominguez et al. 1990).

Sampling sites along with metropolitan and south sectors of Pernambuco’s coast
Sampling design
Non-destructive sampling was performed in 11 sampling sites, all reefs located on the metropolitan and south sectors of Pernambuco coast (8°5′18.14″S, 34°52′44.31″ W and 8°54′16.59″S, 35° 8′16.26″W) (ITEP 2012). The 11 reefs were classified according to the human occupation levels and population density based on urbanization criteria (Projeto Orla 2002). This classification was achieved using a combination of field visits and consulting demographic data (IBGE 2010), local technical reports (SNIS 2011), and processing remote sensing images (Google Earth 2013). Demographic surveys were made in each sector until 1 km away of the sampling site in both directions parallel to the shore, and locations were accordingly classified into three levels of urbanization: non urbanized coast (NU), coast in urbanization process (UP), and consolidated urbanization coast (UC).
The “Line Point Transect” method (Ambrose 2002) was used to quantify the relative abundance of species in the macroalgae community. In this non-destructive method, the observer records in situ the species present in previously defined points at the lowest level (naked eye). Macroalgae were later assigned to a morpho-functional group (MFG) (adapted from Steneck and Dethier 1994; Benedetti-Cecchi 2001; Balata et al. 2011), to each sessile invertebrate a feeding guild (FG) was assigned (Paine 1980). A total of seven transects of 10 m each, perpendicular to the coastline, were done in each reef, starting at the lowest shore levels of the intertidal region. In each transect, we recorded the benthic sessile organism present every 10 cm.
To assess temporal generality of the results, each reef was sampled twice (November 2013 until February 2014 and between May 2014 and August 2014), to avoid pseudoreplication analyses of seasonal changes that were not the target of the present study.
Data analysis
Prior to analyses, data were transformed into relative abundance Ra(i) = Ni/Ta, where Ra(i) is the taxa frequency, N(i) is the number of times the taxa showed up on the sample, and Ta is the amount of points occupied by macroalgae on the transect. The tested hypothesis was: macroalgae dominance patterns differ and vary according to the level of urbanization.
For species taxonomic variables, a two-way permutational multivariate analysis of variance (PERMANOVA) with the factors: “Urbanization” (fixed, 3 levels) and “Time” (random, 2 levels) was performed with 9999 and Monte Carlo permutation. The morpho-functional group/feeding guild (MFG/FG) was compared using ANOVA one-way with the factor urbanization (3 levels). Analyses for species taxonomic variables were performed on fourth-root transformed data by using the Bray-Curtis similarity coefficient. The permutational analysis of multivariate dispersions (PERMADISP) was used to test the homogeneity of urbanization and time factors. The similarity percentage analysis (SIMPER) was used to highlight the contribution of each MFG/FG for differences within levels of urban factor (Clarke and Gorley 2006). An unconstrained ordination (principal coordinates, PCO) was used to graphically represent the results of multivariate data. All analyses were performed using the software PRIMER-E v6 (Clarke and Warwick 2005) PERMANOVA+ (Anderson et al. 2008).
Results
The variables used to classify the scale of environmental stress in the sampling sites are in according to the urbanization degrees and they are listed on Table 1. The beaches Pina, Boa Viagem, and Piedade were classified as UC urbanization degree, due to its high demographic density, with over than 500 inhabitants km−2, over 85% of occupied residences, and in Piedade, only 6.9% of the sewage are collected. Enseada dos Corais, Suape, Serrambi, Tamandaré (Campas), and São José da Coroa Grande beaches were classified as UP for its urbanization degree. Those beaches had demographic densities varying about 20 to 500 inhabitants km−2 and with 29.1 to 76.8% of occupied residences. Finally, the beaches classified as NU urbanization degree were Paiva, Toquinho, and Tamandaré (Mamucabas), with the lowest demographic densities, varying from 5 to 250 inhabitants km−2 (Table 1).
A total of 100 transects were obtained across the 11 sampled reefs, where 51 taxa were identified including 24 Rhodophyta, 11 Chlorophyta, and 6 Ochrophyta, and sessile animals as Porifera, Annelida, Mollusca (2), Cnidaria (3), Echinodermata, and Arthropoda. The macroalgae and their respective MFG/FG identified according to the degree of urbanization are shown in Table 2.
Regarding the urbanization degree, identified macroalgae are distributed in the three phyla in this way: 19 Rhodophyta, 7 Chlorophyta, and 3 Ochrophyta at the NU environment; 22 Rhodophyta, 10 Chlorophyta, and 6 Ochrophyta at the UP environment; and 16 Rhodophyta, 4 Chlorophyta, and 3 Ochrophyta at the UC environment (Table 2). Their relative frequencies were: 63% Rhodophyta, 21% Chlorophyta, and 16% Ochrophyta at NU environment; 62% Rhodophyta, 22% Chlorophyta, and 16% Ochrophyta at UP environment; and 64% Rhodophyta, 32% Chlorophyta, and 4% Ochrophyta at UC environment (Fig. 2).

Relative abundance of macroalgae phyla. a Macroalgae phyla relative abundance at different urbanization degrees: non urbanized (NU), urbanization process (UP), and consolidated urbanization (UC). b Morpho-functional groups and feeding guilds relative abundance at different urbanization degrees: non urbanized (NU), urbanization process (UP), and consolidated urbanization (UC)
The red macroalgae Palisada perforata and Gelidiella acerosa were abundant at all studied sites, but mostly in UP and UC sites, they decreased in abundance with increasing of urbanization levels whereas other macroalgae, such as green Ulva spp., Bryopsis sp., and the red Chondracanthus acicularis became the dominant taxa with increasing of urbanization levels (UC sites). Concerning the MFG, the Foliose, Siphonous, Canopy, and Cnidaria groups presented responses in abundance when compared at different levels of urbanization (Fig. 3, Table 3). The Foliose and Siphonous groups were more abundant at UC sites, gradually decreasing with the environment quality. Canopy and Cnidaria groups had higher abundance at sites with lower urbanization degree.

Abundance of morpho-functional groups and feeding guilds in response to urbanization degree: non urbanized (NU), urbanization process (UP), and consolidated urbanization (UC). Results are presented as mean ± standard error (n = 100)
Macroalgae community structure significantly varied among levels of urbanization (F = 12.89; p = 0.003), which were consistent in time (time × urbanization F = 1.08; p = 0.37). There were significant differences in the structure of macroalgal assemblages between UP and UC (t = 3.52 and p = 0.014) and between NU and UC (t = 4.49 and p = 0.004), but not between NU and UP (t = 1.78 and p = 0.12) (Fig. 4, Table 4). The dispersion of urbanization levels was different (F = 33.366; p = 0.001). UC level was different than UP (t = 8.05; pperm = 0.001) and NU (t = 7.12; pperm = 0.001), while NU and UP levels did not differ, and UC level had the lowest dispersion between the urbanization levels (Table 5).

Principal coordinates (PCO) plot. (circle) Consolidated urbanization, (down-pointing triangle) urbanization process, and (square) non urbanized. Solid symbols represent time I and blank symbols represent time II. The vectors correspond to a multiple correlation > 0.6 of macroalgae taxa: Ulv: Ulva spp., Chon: Chondracanthus acicularis, C.rac: Caulerpa recemosa and Sar: Sargassum sp.; and morpho-functional group and feeding guilds: SI: siphonous; FO: foliose; CA: canopy; CR: Cnidaria
The PCO and SIMPER analyses (Table 6) revealed that in UC, there was generally a greater dominance of filamentous (i.e., Bryopsis sp.), turf-forming (i.e., Chondracanthus sp.), and foliose (i.e. Ulva spp.) algae. In contrast, the canopy alga Sargassum sp. was completely absent from UC, but dominated assemblages in UP and NU situations.
In the PCO plot is possible to identify two groups. Group 1 is formed by all samples from UC sites, and group 2 is composed of samples from NU and UP sites (Fig. 4).
Discussion
Reefs classified as UC were located among one of the largest metropolitan regions of Brazil (Recife’s Metropolitan Region). Despite sewage treatments, only 36% of the sewage is currently collected (SNIS 2011) leaving a substantial proportion of sewage still running free into the coastal region. UP reefs receive seasonal population booms (activity during summer), and due to their greater distance to larger urban centers, the sewage collecting system is poor. The few places classified as NU were essentially remote locations with little levels of visitation, even during summer.
Morpho-functional groups of phytobenthos and feeding guilds of zoobenthos responded, in parts, to the cover variation of the substrate according to the urbanization degree. The Cnidaria group, composed by Zoanthus, Palythoa, and Protopalythoa, were more frequent at places of low urbanization degree, even though they were still present, at lower frequency, at urbanized places (Portugal et al. 2016). When found in places with low impact, Zoanthus spp. directly compete with macroalgae such as Canopy (Sargassum, in tropical environments), at lower regions of the intertidal zone (Santos et al. 2015).
The studied places supported macroalgal assemblages typical from the tropical phycogeographic region (Horta et al. 2001), although there were variations on the phyla proportions when places were analyzed for their urbanization degree.
The studied reef formation presented high abundance values of macroalgae such as Palisada perforata, Gelidiella acerosa, Gracilaria sp., and Caulerpa racemosa, which are dominant algae of the reef plateau from tropical regions (Pereira et al. 2002, 2008; Villaça et al. 2010; Fonseca et al. 2012).
On the low mesolitoral region reefs, we found species (Sargassum sp., Padina sp., and Hypnea musciformis) that dominate places with great influence from littoral, in other words, places on the reef that are constantly washed by waves, allowing organisms to be wet (Vasconcelos et al. 2013). At the same way as the Fucales, Cystoseira are key species at temperate and subtropical places, responding differently to the environment impacts (Mangialajo et al. 2008), Sargassum species perform this same structuring role at shallow infralittoral environments from tropical places. They are resilient to a series of natural impacts (Loffler and Hoey 2018), but anthropic pressure could decrease their population (Coelho et al. 2000).
We also identified another group of macroalgae composed by Ceramiaceae species, Ulva spp. and Gelidium sp., which cannot stand great hydrodynamics, or cannot compete to colonize the reef edge surroundings, often placing themselves next to plateau and top of the reef, tolerating the desiccation stress.
The abundance of P. perforata on reef formations is directly connected to high-temperature environments (above 28 °C) and air exposure (Tsai et al. 2005), typical of a mesolittoral environment. However, this macroalgae relative abundance responded negatively to the UC urbanization level on the sampled locations. In a reef environment dominated by Gracilaria coronopifolia J. Agardh and P. perforata (previously reported as Laurencia papillosa) (Tsai et al. 2005), the first one takes advantage when there is a eutrophication process in the area, leading to diminish the occurrence of P. perforata, showing that this alga decreases its dominance on impacted environments, based on diverse descriptors (Cabrera et al. 2005).
The dominant taxa P. perforata, Sargassum sp., and Amansia sp. on the beaches with NU and UP urbanization degrees were replaced by Ulva spp., Bryopsis spp., and C. acicularis on the UC beaches. Macroalgae from Ulva and Bryopsis genus have been well documented and classified as opportunistic algae that take advantage in eutrophicated environments (Ortega 2000; Areces 2001; Juanes et al. 2008) and that are greatly related to urbanized environments (Martins et al. 2012; Scherner et al. 2013; Orlandi et al. 2014). Chondracanthus acicularis was classified as opportunistic algae (Orfanidis et al. 2003), with fast growth rate and short life cycles, present in areas with organic contamination, indicating environmental impact (Portugal et al. 2016). Gracilaria sp. was also present in high urbanization degree environments. Those macroalgae genera are capable to resist changes in salinity, temperature oscillations, high nutrient concentration, and even landfilling by suspension particles (Lapointe 1989; Santelices and Doty 1989; Peckol and Rivers 1996; Moreira et al. 2006).
Impacts associated to UC sites lead to the domination of opportunistic species, which causes huge ecological consequences, since the disappearance of these structural and engineering species involves the loss of entire communities, the regression of other species, and the homogenization of the underwater landscape (Thibaut et al. 2005).
Using a fast and easy data achievement method (Line Point Transect) and a relative abundance descriptor, it was possible to verify changes on macroalgae dominance in the coastal reef environments classified into different urbanization degrees. The present study results also appoint to importance not only of the taxa occurrence, but mostly of the indicator dominant taxa relative abundance. The reef formation leads the community to a natural heterogeneity, but in the presence of an urbanization gradient, it tends to shift and simplify the community in the reefs observed.
Associated to our findings, future monitoring studies that consider temporal/seasonal scales could elucidate the community dynamics against urbanization impacts, and hence take measures to preserve tropical coastal environments. Also, it becomes necessary to find a way to quali-quantitatively evaluate the evolution of those environments due to increasing coast area urbanization, through environmental quality indexes using key elements from ecosystems.
References
Airoldi L (2003) The effects of sedimentation on rocky coast assemblages. Oceanogr Mar Biol 41:161–236
Airoldi L, Beck WM (2007) Loss, status and trends for coastal marine habitats of Europe. Oceanogr Mar Biol 45:345–405
Ambrose RF (2002) Transects, quadrats, and other sampling units. In: Murray SN, Ambrose RF, Dethier MN (eds) Methods for performing monitoring, impact, and ecological studies on rocky shores. Coastal Research Center, Marine Science Institute, University of California, Santa Barbara, pp 98–122
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for Primer. Primer-E, Plymouth
Areces A (2001) La ficoflora intermareal como bioindicadora de calidad ambiental. Estudio de caso: El litoral habanero. In: Alveal K, Antezana T (eds) Sustentabilidad de la biodiversidade. Um problema actual, bases científico-técnicas, teorizaciones y perspectivas. Trama Impresores S.A, Concepción, pp 569–589
Balata D, Piazzi L, Rindi F (2011) Testing a new classification of morphological functional groups of marine macroalgae for the detection of responses to stress. Mar Biol 158:2459–2469
Benedetti-Cecchi L (2001) Variability in abundance of algae and invertebrates at different spatial scales on rocky sea shores. Mar Ecol Prog Ser 215:79–92
Benedetti-Cecchi L, Pannacciulli F, Bulleri F, Moschella PS, Airoldi L, Relini G, Cinelli F (2001) Predicting the consequences of anthropogenic disturbance: large-scale effects of loss of canopy algae on rocky shores. Mar Ecol Prog Ser 214:137–150
Borowitzka MA (1972) Intertidal algal species diversity and the effects of pollution. Aust J Mar Freshwat Res 25:73–84
Branner JC (1904) The stone reefs of Brazil, their geological and geographical relations, with a chapter on the coral reefs, vol 54. Bulletin of the Museum of Comparative Zoology, Cambridge, pp 1–285
Cabrera R, Moreira A, Primelles J, Suárez AM (2005) Variación de la biomasa de Chondrophycus papillosus (C. Agardh) Garbary et Harper (Ceramiales: Rhodophyta) y su epifitismo en la bahía de Nuevitas. Cuba Rev Invest Mar 26:15–20
Castro CB, Pires DO (2001) Brazilian coral reefs: what we know and what is still missing. Bull Mar Sci 69(2):357–371
Clarke KR, Gorley RN (2006) PRIMER v6: user manual/tutorial (Plymouth routines in multivariate ecological research). Primer-E Ltd, Plymouth
Clarke KR, Warwick RM (2005) Primer-6 computer program. Natural Environment Research Council, Plymouth
Coelho SM, Rijstenbil JW, Brown MT (2000) Impacts of anthropogenic stresses on the early development stages of seaweed. J Aquat Ecosyst Stress Recov 7:317–333
Connell SD, Russell BD, Turner DJ, Shepherd SA, Kildea T, Miller D, Airoldi L, Cheshire A (2008) Recovering a lost baseline: missing kelp forests from a metropolitan coast. Mar Ecol Prog Ser 360:63–72
CPRH (2003) Agência estadual de meio ambiente e recursos hídricos. Diagnóstico Sócioambiental do Litoral Sul de Pernambuco Avaiable at: http://wwwcprhpegovbr. Acessed 26 June 2015
Davison IR, Pearson GA (1996) Stress tolerance in intertidal seaweeds. J Phycol 32:197–211
Dominguez JML, Bittencourt ACSP, Leão ZMAN, Azevedo AEG (1990) Geologia do quaternário costeiro do Estado de Pernambuco. Rev Bras Geociênc 20:208–215
Fonseca AC, Villaça R, Knoppers B (2012) Reef flat community structure of Atol das Rocas, Northeast Brazil and Southwest Atlantic. J Mar Biol 2012:1–10
Goatley CH, Bellwood DR (2013) Ecological consequences of sediment on high-energy coral reefs. PLoS One 8(10):e77737
Google Earth® (2013) Google Inc.
Halpern BS, Longo C, Hardy D, McLeod KL, Samhouri JF, Katona SK et al (2012) An index to assess the health and benefits of the global ocean. Nature 488:615–620
Horta PA, Amancio E, Coimbra CS, Oliveira EC (2001) Considerações sobre a distribuição e origem da flora de macroalgas marinhas brasileiras. Hoehnea 28:243–265
IBGE (2010) Instituto Brasileiro de Geografia e Estatística Censo Demográfico. http://www.censo2010.ibge.gov.br. Acessed 26 June 2015
ITEP (2012) Instituto de Tecnologia de Pernambuco. Relatório de impacto ambiental. Recuperação da orla marítima. Municípios de Jaboatão dos Guararapes, Recife, Olinda e Paulista (Pernambuco). Instituto de Tecnologia de Pernambuco, Recife, pp 1–98
Johnston EL, Roberts DA (2009) Contaminants reduce the richness and evenness of marine communities: a review and meta-analysis. Environ Pollut 157:1745–1752
Juanes JA, Guinda X, Puente A, Revilla JA (2008) Macroalgae, a suitable indicator of the ecological status of coastal rocky communities in the NE Atlantic. Ecol Indic 8:351–359
Köppen W (1948) Climatologia: con un estudio de los climas de la tierra. Fondo de Cultura Econômica, Mexico, pp 1–479
Krause-Jensen D, Sagert S, Schubert H, Bostrom C (2008) Empirical relationships linking distribution and abundance of marine vegetation to eutrophication. Ecol Indic 8:515–529
Laborel JL (1969) Madreporaires et hidrocoralliaires recifaux dês cotes brésiliennes. Systematíque, ecologie, repartition verticale et geographic. Ann Inst Oceanogr Paris 47:171–226
Lapointe BE (1989) Macroalgal production and nutrient relations in oligotrophic areas of Florida bay. Bull Mar Sci 44:312–323
Loffler Z, Hoey AS (2018) Canopy-forming macroalgal beds (Sargassum) on coral reefs are resilient to physical disturbance. J Ecol 106:1156–1164
Mangialajo L, Chiantore M, Cattaneo-Vietti R (2008) Loss of fucoid algae along a gradient of urbanisation, and structure of benthic assemblages. Mar Ecol Prog Ser 358:63–74
Manso VAV, Coutinho PN, Guerra NC, Soares CFA (2006) Pernambuco. In: Muehe D (ed) Erosão e Progradação no Litoral Brasileiro. Ministério do Meio Ambiente, Brasília, pp 179–196
Martins CD, Arantes N, Faveri C, Batista MB, Oliveira EC, Pagliosa PR, Fonseca AL, Nunes JMC, Chow F, Pereira SB, Horta PA (2012) The impact of coastal urbanization on the structure of phytobenthic communities in southern Brazil. Mar Pollut Bull 64:772–778
Moreira AR, Armenteros M, Gómez M, Leon AR, Cabrera R, Castellanos ME, Munoz A, Suarez AM (2006) Variation of macroalgae biomass in Cienfuegos bay, Cuba. Rev Investig Mar 27:3–12
Orfanidis S, Panayotidis P, Stamatis N (2003) An insight to the ecological evaluation index (EEI). Ecol Indic 3:27–33
Orlandi L, Bentivogli F, Carlino P, Calizza E, Rossi D, Costantini ML, Rossi L (2014) δ15 N variation in Ulva lactuca as a proxy for anthropogenic nitrogen inputs in coastal areas of gulf of Gaeta (Mediterranean Sea). Mar Pollut Bull 84:76–82
Ortega JLG (2000) Algas. In: Espino GL, Pulido J, Pérez JLC (eds) Organismos indicadores de la calidad del água y de la contaminación (Bioindicadores). Playa y Valdés, Mexico, pp 109–193
Paine RT (1980) Food webs: linkage, interaction strength and community infrastructure. J Anim Ecol 49:667–685
Peckol P, Rivers JS (1996) Contribution by macroalgal mats to primary production of a shallow embayment under high and low nitrogen-loading rates. Estuar Coast Shelf Sci 43:311–325
Pereira SMB, Carvalho MFO, Angeiras JA, Pedrosa MEB, Oliveira NMB, et al (2002) Algas marinhas bentônicas do estado de Pernambuco. In: Tabarelli M, Silva JMC (ed) Diagnóstico da biodiversidade de Pernambuco. Secretaria de Ciência, Tecnologia e Meio Ambiente, Massangana, Recife, 1, p 97–124
Pereira SMB, Oliveira-Carvalho MF, Burgos DC, Araújo EL (2008) Caracterização estrutural das macroalgas de ambiente recifal da praia de Enseada dos Corais. In: Congresso Brasileiro de Ficologia, XI, Itajaí-SC. Anais de trabalho completos. Série Livros do Museu Nacional. Rio de Janeiro, p 231–242
Portugal AB, Carvalho FL, de Macedo Carneiro PB, Rossi S, de Oliveira Soares M (2016) Increased anthropogenic pressure decreases species richness in tropical intertidal reefs. Mar Environ Res 120:44–54
Projeto Orla (2002) Projeto Orla: fundamentos para gestão integrada. Brasília: MMA/SQA; Brasília: MP/SPU, p 78
Santelices B, Doty MS (1989) A review of Gracilaria farming. Aquaculture 78:95–133
Santos GS, Lira SMA, Schwamborn R (2015) Análise das comunidades macrobentônicos sésseis, com ênfase na interação entre o cnidário Zoanthus sociatus (Ellis, 1768) e macroalgas, no topo de um recife de águas rasas do nordeste do Brasil. Trop Oceanogr 43:1–8
Scherner F, Horta PA, Oliveira EC, Simonassi JC, Hall-Spencer JM, Chow F, Nunes JM, Prereira SM (2013) Coastal urbanization leads to remarkable seaweed species loss and community shifts along the SW Atlantic. Mar Pollut Bull 76:106–115
Schiel DR (2006) Rivets or bolts? When single species count in the function of temperate rocky reef communities. J Exp Mar Biol Ecol 338:233–252
Shanmugam P, Neelamani S, Ahn YH, Philip L, Hong GH (2007) Assessment of the levels of coastal marine pollution of Chennai city, southern India. Water Resour Manag 21:1187–1206
SNIS (2011) Sistema Nacional de Informações sobre Saneamento: Diagnóstico dos serviços de água e esgotos. Ministério das cidades SNSA, Brasília, p 432
Spalding MD, Fox HE, Allen GR, Davidson N, Ferdaña ZA, Finlayson M, Halpern BS, Jorge MA, Lombana A, Lourie SA, Martin KD, McManus E, Molnar J, Recchia CA, Robertson J (2007) Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience 57:573–583
Steneck RS, Dethier MN (1994) A functional group approach to the structure of algal dominated communities. Oikos 69:476–498
Tait LW, Schiel DR (2011) Legacy effects of canopy disturbance on ecosystem functioning in macroalgal assemblages. PLoS One 6:e26986
Thibaut T, Pinedo S, Torras X, Ballesteros E (2005) Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albéres coast (France, North-Western Mediterranean). Mar Pollut Bull 50:1472–1489
Tsai C, Chang J, Sheu F, Shyu Y, Yu A, Wong S, Dai C, Lee T (2005) Seasonal growth dynamics of Laurencia papillosa and Gracilaria coronopifolia and from a highly eutrophic reef in southern Taiwan: temperature limitation and nutrient availability. J Exp Mar Biol Ecol 315:49–69
Turra A, Croquer A, Carranza A, Mansilla A, Areces AJ, Werlinger C et al (2013) Global environmental changes: setting priorities for Latin American coastal habitats. Glob Chang Biol 19:1965–1969
Vasconcelos ERTPP, Reis TNV, Guimarães-Barros NC, Bernardi J, Areces-Mallea AJ, Cocentino ALM, Fujii MT (2013) Padrão espacial da comunidade de macroalgas de mesolitoral em ambiente recifal do nordeste brasileiro. Trop Oceanogr 41:84–92
Villaça R, Fonseca AC, Jensen VK, Knoppers B (2010) Species composition and distribution of macroalgae on Atol das Rocas, Brazil, SW Atlantic. Bot Mar 53:113–122
Worm B, Lotze HK, Boström C, Engkvist R, Labanauskas V, Sommer U (1999) Marine diversity shift linked to interactions among grazers, nutrients and propagule banks. Mar Ecol Prog Ser 185:309–314
Acknowledgments
The first author thanks the Fundação de Amparo à Ciência e Tecnologia de Pernambuco (FACEPE) for the PhD Scholarship and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the Science without Frontier Scholarship. MTF thanks CNPq for the Productivity Fellowship (Proc. 303915/2013-7). The authors thank Souza-Filho JS, Neumann-Leitão S, Feitosa FA, Rosa-Filho JS for helpful advice and comments.
Funding
This work was supported by research grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq-Proc. 484647/2012-1).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
de Vasconcelos, E.R.T.P.P., Vasconcelos, J.B., Reis, T.N.d. et al. Macroalgal responses to coastal urbanization: relative abundance of indicator species. J Appl Phycol 31, 893–903 (2019). https://doi.org/10.1007/s10811-018-1639-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-018-1639-3