
Abstract
We investigated the anti-inflammatory and analgesic effects of quercetin in monosodium urate crystals (MSU)-induced gout arthritis, and the sensitivity of quercetin effects to naloxone, an opioid receptor antagonist. Mice were treated with quercetin, and mechanical hyperalgesia was assessed at 1–24 h after MSU injection. In vivo, leukocyte recruitment, cytokine levels, oxidative stress, NFκB activation, and gp91phox and inflammasome components (NLRP3, ASC, Pro-caspase-1, and Pro-IL-1β) mRNA expression by qPCR were determined in the knee joints at 24 h after MSU injection. Inflammasome activation was determined, in vitro, in lipopolysaccharide-primed macrophages challenged with MSU. Quercetin inhibited MSU-induced mechanical hyperalgesia, leukocyte recruitment, TNFα and IL-1β production, superoxide anion production, inflammasome activation, decrease of antioxidants levels, NFκB activation, and inflammasome components mRNA expression. Naloxone pre-treatment prevented all the inhibitory effects of quercetin over MSU-induced gout arthritis. These results demonstrate that quercetin exerts analgesic and anti-inflammatory effect in the MSU-induced arthritis in a naloxone-sensitive manner.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Gout is an arthropathy characterized by intense inflammation and pain induced by monosodium urate (MSU) crystal deposition into the articular joint and surrounding tissues (Khanna et al. 2012; McGettrick and O’Neill 2013). The interaction of MSU crystals with phagocytes such as macrophages and infiltrating leukocytes induces the secretion of various inflammatory mediators, consequently triggering a typical inflammatory response and increased pain sensitivity, provoking hyperalgesia (Verri et al. 2006; Martin et al. 2009; Zarpelon et al. 2013). It is well known that MSU crystals undergo phagocytosis followed by rupture of the phagolysosome and activation of NLRP3 inflammasome, which culminates in the release of the active form of IL-1β. It has also been reported that patients with gout arthritis show a significant response to IL-1 inhibition, indicating the crucial role of IL-1β in driving intense inflammatory pathology (Lu et al. 2014). Furthermore, IL-1β activates synoviocyte-like macrophages promoting the release of additional pro-inflammatory cytokines such IL-8, IL-17, and TNFα (Martinon et al. 2006; Lu et al. 2014). Inflammatory cytokines such as IL-1β and TNFα activate the transcription factor NF-κB resulting in production of chemotactic molecules including IL-8 to induce neutrophil chemoattraction (Martinon et al. 2006; Neuschäfer-Rube et al. 2013; Dhanasekar et al. 2015).
The current therapeutic approaches against gouty arthritis are given mainly by the treatment of inflammation and the control of hyperuricemia. For instance, allopurinol acts by inhibiting hepatic xanthine oxidase enzyme (XOD) activity and then reducing serum uric acid levels. However, allopurinol is not able to control the acute gout attacks (Dubchak and Falasca 2010), and also can induce several side effects such as fever, skin rashes, allergic reactions, hepatitis, and nephropathy (Umamaheswari et al. 2009). Thus, non-steroidal anti-inflammatory (NSAIDs) drugs (e.g., indomethacin) and alkaloid drugs (e.g., colchicine) are frequently used as the first-line approach therapies for treatment of acute gout inflammation. Nevertheless, these classes of drugs induce severe side effects such as gastrointestinal and renal toxicity, and gastrointestinal bleeding (Cronstein and Terkeltaub 2006). Immunobiological therapies targeting IL-1β are also useful in the treatment of gout arthritis attack, but the high cost reduces its wide clinical use (Dubchak and Falasca 2010). Thus, the research on novel drugs to treat gout arthritis has been a growing focus of interest (Sabina et al. 2011; de Souza et al. 2012; Rukdee et al. 2015; Zhao and Huang 2015; Xu et al. 2016). Therapeutic natural compounds are a growing field, since current therapies have relayed on natural compounds to develop efficacious treatments such as morphine, capsaicin, salicylate, and menthol (Julius 2013).
Flavonoids are multi-target molecules that inhibit inflammation and oxidative stress (Pinho-Ribeiro et al. 2015; Martinez et al. 2015a, b; Sun et al. 2016; Kim and Park 2016; Manchope et al. 2016; Liu et al. 2016). Evidence supports the safety of flavonoids due to low or no toxic effect contrasting to non-steroidal anti-inflammatory drugs (Okamoto 2005; Pinho-Ribeiro et al. 2016). Quercetin, a bioflavonoid presenting low toxicity (Okamoto 2005) and several biological properties such as antioxidant and anti-inflammatory (Valério et al. 2009; Wang et al. 2012; Guazelli et al. 2013; Sun et al. 2015a), has been a focus of varied studies on pain and inflammation (Anjaneyulu and Chopra 2003a; Valério et al. 2009; Souto et al. 2011; Calixto-Campos et al. 2015). Quercetin acts in an opioid-dependent manner inducing analgesia in diabetic- and cancer-induced pain (Anjaneyulu and Chopra 2003a; Calixto-Campos et al. 2015). Quercetin inhibits MSU-induced knee joint edema, histological leukocyte infiltration, IL-1β, TNFα, prostaglandin E2 and nitric oxide production, cyclooxygenase-2 expression, and lipid peroxidation. Furthermore, quercetin treatment reverted MSU-induced reduction of superoxide dismutase, catalase, and glutathione peroxidase activities in rats (Huang et al. 2012). However, it remains to be determined the molecular mechanisms of gout arthritis physiopathology that are targeted by quercetin.
This study aimed to investigate the effect of quercetin treatment in experimental gout arthritis induced by intra-articular injection of MSU assessing its analgesic, anti-inflammatory and antioxidant effects, including the NFκB and NLRP3 inflammasome pathways. Hence, we evaluated the contribution of endogenous opioid in the effect of quercetin.
Materials and methods
Animals
The experiments were performed with male Swiss mice (20–25 g, Universidade Estadual de Londrina, Londrina, PR, Brazil) housed in standard clear plastic cages (5–6 per cage) with free access to food and water. All testing was performed between 9:00 a.m. and 5:00 p.m. in a temperature-controlled room. Animal care and handling procedures were performed with the approval of the Ethics Committee of the Universidade Estadual de Londrina (Process Number 13279.2011.76) and followed the International Association for the Study of Pain guidelines as described by Zimmermann (1983). All efforts were made to minimize the number of animals used and their suffering.
Drugs and reagents
The following materials were obtained from the sources indicated: Quercetin at 95% purity from Acros (Pittsburg, PA); MSU and naloxone were from Sigma-Aldrich (St Louis, MO); Boric acid was from Promega, and uricase/fasturtec was from Sanofi-Synthelabo. ELISA kits to determine murine TNF-α and IL-1β were obtained from eBioscience (San Diego, CA).
MSU crystal preparation
MSU crystals were prepared according to the method described previously (Nishimura et al. 1997). In brief, 800 mg of monosodium urate was dissolved in 155 mL of boiling water containing 5 mL of 1 N NaOH. After the pH was adjusted to 7.2, the solution was gradually cooled by stirring at room temperature. The crystals were collected by centrifugation at 3000g for 2 min at 4 °C. The crystals were evaporated and sterilized by heating at 180 °C for 2 h and stored in a sterile environment until use.
Experimental protocols
In the first set of experiments, mice were treated with quercetin (10–100 mg/kg, 20% Tween 80, s.c., 30 min) or vehicle (20% tween 80 in saline) before the MSU injection (100 μg/10 μL per knee joint) to determine the dose–response of quercetin (Fig. 1a). The intensity of mechanical hyperalgesia was measured 1–24 h after MSU injection by the electronic pressure-meter test. To investigate the participation of endogenous opioids in the analgesic and anti-inflammatory effects of quercetin, mice were pre-treated with naloxone (0.3–3 mg/kg, i.p., 1 h) before quercetin treatment (100 mg/kg, s.c.) or vehicle (saline). The inflammatory parameters were evaluated at 24 h after injection of the MSU crystals. After 24 h, the articular cavities were washed with saline/EDTA solution for leukocytes counts and differential cell counts determination. The inflammatory cell infiltrate (histopathological analysis), cytokines, NFκB activation, inflammasome components mRNA expression (NLRP3, ASC, Pro-caspase-1, and Pro-IL-1β), antioxidant capacity (GSH, FRAP, and ABTS assay), superoxide anion production (NBT reduction), gp91phox, Nrf2, and HO-1 mRNA expression by qPCR were determined in the articular tissue. In vitro (Fig. 1b), bone marrow-derived macrophages (BMDMs) were cultured in 96-well plates. The activation of BMDMs first signal was induced by LPS (500 ng/mL) stimulation during 3 h. The participation of endogenous opioid in quercetin effect was assessed using naloxone (0.2–20 µM) treatment of BMDMs 1 h before quercetin treatment. In addition, quercetin (0.3–30 µM) and morphine (0.2–20 µM) treatments were performed 30 min before stimulation with MSU crystals (450 µg/mL—second signal) and incubated during 5 h. ELISA was used to measure the IL-1β levels in the culture supernatant.

Schematic experimental protocols. In vivo a: Mice were anesthetised (10 mg/kg ketamine:xylazine injected intraperitoneally [i.p.]) and were injected with MSU crystals (100 μg/10 μL) into the tibio-femoral knee joint. In the first set of experiments, mice were treated with quercetin (10–100 mg/kg, s.c. [Subcutaneous], 30 min) or vehicle (20% tween 80 in saline) before MSU injection. The intensity of mechanical hyperalgesia was measured 1–24 h after MSU injection by the electronic pressure-meter test. To determine the participation of endogenous opioid on quercetin analgesic and anti-inflammatory effect, mice were pre-treated with naloxone (0.3–3 mg/kg, i.p., 1 h) before quercetin treatment (100 mg/kg, s.c.) or vehicle. The inflammation parameters were evaluated at 24 h after injection of the MSU crystals. After 24 h, the articular cavities were washed with saline/EDTA solution for leukocytes counts and differential determination. Periarticular tissues were removed from the joints for evaluation of synovitis in articular tissue, cytokines, NFκB activation, inflammasome components mRNA expression (NLRP3, ASC, Pro-caspase-1 and Pro-IL-1β), evaluation of antioxidant capacity (GSH, FRAP, and ABTS assay), superoxide anion production (NBT reduction), Gp91phox, Nrf2 and HO-1 by qPCR. In vitro b: The macrophages BMDMs were performed in 96-well plate, LPS (500 ng/mL)-primed BMDM was incubated during 3 h, after that naloxone (0.2–20 µM) incubated with BDMNs cells 1 h before quercetin treatment, in addition, quercetin (0.3–30 µM) or morphine (0.2–20 µM) treatment performed during 30 min before stimulation with MSU crystals (450 µg/mL) and incubated during 5 h. IL-1β levels in the culture supernatant were measured
Electronic pressure-meter test (mechanical hyperalgesia)
Evaluation of mechanical hyperalgesia was performed as described previously (Guerrero et al. 2006) using an electronic pressure-meter (INSIGHT Instruments). The flexion withdrawal threshold was used to in behavioral responses associated with pain. Results are expressed in grams force (g).
Leukocyte total and differential counts
To determine the leukocyte recruitment to the articular cavities, joint cavities were washed three times with 3.3 μL of PBS containing 1 mM EDTA. The total numbers of leukocytes were determined in Neubauer chamber diluted in Turk’s solution. Differential cell counts were determined in cytocentrifuge Rosenfeld stained slices (Cytospin 4; Shandon, Pittsburg, PA). Total and differential cell counts were performed with a light microscope and the results were expressed as the number (mean ± SEM) of total of leukocytes or neutrophils ×103 per cavity (Verri et al. 2010).
Histopathological analysis
Mice were sacrificed 24 h after MSU injection. The articular joint of mice was removed, fixed with 10% paraformaldehyde in PBS, and then decalcified for 10 days with EDTA and embedded in paraffin for histological analysis. The paraffin sections were stained with hematoxylin and eosin for the conventional morphological evaluation. Dimension used for the analysis was 616 × 662 pixels for analysis (Field) and magnification 400× and the arrow indicate representative infiltrate inflammatory cells counted.
Cytokine production
At 24 h after the i.a. injection of MSU, animals were terminally anaesthetized, and the samples of knee joints were removed and homogenized in 500 μL of the appropriate buffer containing protease inhibitors. The TNF-α and IL-1β levels were determined as described previously (Verri et al. 2010) by enzyme-linked immunosorbent assay (ELISA) kits following the manufacturer’s instructions (eBioscience). The results were expressed as picograms (pg) of each cytokine per 100 mg of tissue weight. As a negative control, the concentrations of these cytokines were determined in mice injected with saline.
NF-κB activation
The articular joint samples were collected and homogenized in ice-cold lysis buffer (Cell Signaling). The homogenates were centrifuged (14,000 rpm × 10 min × 4 °C), with the supernatants used to assess the levels of phosphorylated and total NF-κB p65 subunit by ELISA using PathScan® kits (Cell Signaling) according to the manufacturer’s directions. The results represent the sample ratio (total p65/phospho-p65) measured at 450 nm (Multiskan GO Thermo Scientific).
Quantitative polymerase chain reaction (qPCR)
The knee joint samples were collected at 24 h after the i.a. injection of MSU, animals were terminally anaesthetized and homogenized in 500 µL of Trizol reagent and centrifuged (12,000 rcf × 15 min × 4 °C), and total RNA was measured with a spectrophotometer and the wavelength absorption ratio (260/280 nm) was between 1.8 and 2.0 for all preparations and extracted using the SV Total RNA Isolation System (Promega) (Verri et al. 2008). All reactions were performed in triplicate using the following cycling conditions: 50 °C for 2 min, 95 °C for 2 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 30 s. qPCR was performed in a LightCycler Nano Instrument (Roche, Mississauga, ON, USA) sequence detection system using the Platinum SYBR Green qPCR SuperMix UDG (Invitrogen, USA). The mRNA level of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. The primers used were Gapdh forward: CAT ACC AGG AAA TGA GCT TG, reverse: ATG ACA TCA AGA AGG TGG TG; Nrf2, forward: TCA CAC GAG ATG AGC TTA GGG CAA, reverse: TAC AGT TCT GGG CGG CGA CTT TAT; Gp91phox, forward: AGC TAT GAG GTG GTG ATG TTA GTG G, reverse: CAC AAT ATT TGT ACC AGA CAG ACT TGA G; Nlrp3, forward: AGC TAT GAG GTG GTG ATG TTA GTG G, reverse: CAC AAT ATT TGT ACC AGA CAG ACT TGA G; HO-1, forward: CCC AAA ACT GGC CTG TAA AA, reverse: CGT GGT CAG TCA ACA TGG AT; Pro-caspase-1: forward: TGG TCT TGT GAC TTG GAG GA, reverse: TGG CTT CTT ATT GGC ACG AT; Pro-IL-1β, forward: GAA ATG CCA CCT TTT GAC AGT G, reverse: TGG ATG CTC TCA TCA GGA CAG; ASC, forward: ATG GGG CGG GCA CGA GAT G, reverse: GCT CTG CTC CAG GTC CAT CAC. The SYBR green PCR Master Mix was used according to the manufacturer’s instructions.
Reduced glutathione (GSH) measurement
At 24 h after the i.a. injection of MSU, animals were terminally anaesthetized and the samples of knee joints were removed and maintained at −80 °C for at least 48 h. The samples were homogenized with 200 µL of 0.02 M EDTA. The homogenate was mixed with 25 μL of 50% trichloroacetic acid and was homogenized three times during 15 min. The mixture was centrifuged (15 min × 1500g × 4 °C). The supernatant was added to 200 μL of 0.2 M TRIS buffer, pH 8.2, and 10 μL of 0.01 M DTNB. After 5 min, the absorbance was measured at 412 nm against a reagent blank with no supernatant. A standard curve was performed with standard GSH. The results are expressed as GSH per milligram of tissue (Sedlak and Lindsay 1968; Borghi et al. 2013; Staurengo-Ferrari et al. 2014).
Nitrobluetetrazolium (NBT) test
The quantitation of O2 − production in tissue homogenate (10 mg/mL in 1.15% KCl) was performed using the nitroblue tetrazolium test (NBT). Briefly, 50 µL of homogenate tissue was incubated with 100 µL of NBT (1 mg/mL) in 96-well plates at 37 °C for 1 h. The supernatant was then removed and the reduced formazan solubilized by adding 120 µL of 2 M KOH and 120 µL of DMSO. The NBT reduction was measured using a Multiscan spectrophotometer at 600 nm. The results were expressed as optical density per milligram of tissue.
Ferric-reducing ability potential (FRAP) and free-radical scavenging ability (ABTS) assays
The samples of knee joints were collected, immediately homogenized with 500 μL of 1.15% KCl, and centrifuged (10 min × 200g × 4 °C). The ability of the sample to resist oxidative damage was determined as ferric-reducing ability using the FRAP assay and as free-radical scavenging ability using the ABTS assay (Borghi et al. 2013). For the FRAP assay, 50 μL of supernatant was mixed with 150 μL of deionized water and 1.5 mL of freshly prepared FRAP reagent. The reaction mixture was incubated at 37 °C for 30 min, and the absorbance was measured at 595 nm. For the ABTS assay, the ABTS solution was diluted with phosphate buffer saline at pH 7.4 to an absorbance of 0.80 at 730 nm. Then, 1.0 mL of diluted ABTS solution was mixed with 20 μL of supernatant. After 6 min, the absorbance was measured at 730 nm. The results were equated against a Trolox standard curve (1.5–30 μmol/L, final concentrations). The results are expressed as Trolox equivalents per milligram of tissue weight in both assays.
Preparation of bone marrow-derived macrophages (BMDMs) and inflammasome activation assay
Femora and tibiae of mice C57BL/6 mice (8 weeks old) were aspirated with RPMI1640 media. Bone marrow cells were cultured in RPMI 1640 medium containing 10% FBS and 15% L929 cell conditioned medium. BMDM were harvested at day 7 and plated at the density of 1.5 × 105 cells/well in 96-well plate. BMDM were stimulated with 500 ng/mL E. coli LPS (Santa Cruz Biotechnology) and 3 h later treated with 450 µg/mL of MSU to stimulate NLRP3 inflammasome activation as described previously (Martinon et al. 2006). BMDM were treated with quercetin (0.3; 3 or 30 µM), or morphine (0.2; 2 or 20 µM), 30 min before MSU stimulation. Naloxone (0.2; 2 or 20 µM) was added 1 h prior to quercetin treatment (Khabbazi et al. 2015). Supernatants were collected 5 h after MSU stimulation and IL-1β concentration quantitated by ELISA.
Data analyses
Data were analyzed using GraphPad Prism statistical software (GraphPad Software, Inc., USA-500.288, version 5.0). Results are presented as mean ± SEM of measurements made on six mice per group per experiment and are representative of two independent experiments. Two-way ANOVA was used to compare the groups and doses at all times when the parameters were measured at different times after the stimulus injection. The analyzed factors were treatments, time, and time versus treatment interaction. One-way ANOVA followed by Tukey’s test was performed for each time-point. P < 0.05 was considered significant.
Results
Quercetin inhibits MSU-induced mechanical hyperalgesia and leukocyte recruitment in gout arthritis
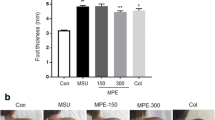
Quercetin decreased the parameters of mechanical hyperalgesia and leukocyte recruitment; however, the dose of 10 mg/kg of quercetin affect MSU-induced mechanical hyperalgesia only at 1 h after MSU injection. On the other hand, the dose of 30 mg/kg inhibited at 1, 5, 7, and 24 h, and the dose of 100 mg/kg inhibited all points evaluated of mechanical hyperalgesia (Fig. 2a). Only dose of 100 mg/kg of quercetin inhibited the MSU-induced recruitment of total of leukocytes and neutrophils to the knee joint (Fig. 2b, c). Based on the results of Fig. 2, the dose of 100 mg/kg of quercetin was selected for next experiments (Fig. 2a, b).

Quercetin dose-dependently reduces monosodium urate crystal (MSU)-induced mechanical hyperalgesia and leukocytes migration. Treatment with quercetin (10, 30 and 100 mg/kg, s.c.) or vehicle (Tween 80 plus saline) was performed 30 min before stimulus with MSU (100 µg/10 µL, intra-articular [i.a.]). The mechanical hyperalgesia (a) was evaluated 1–24 h after stimulus injection with an electronic pressure-meter test. The total of leukocytes (b) and neutrophils (c) counts was determined using Newbauer chamber and Rosenfelt stained slices 24 h after MSU injection. Results are presented as mean ± SEM of six mice per group per experiment, and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared with MSU + vehicle group and **P < 0.05 compared with 10 mg/kg of Quercetin. ANOVA followed by Tukey’s test
Quercetin inhibits MSU-induced mechanical hyperalgesia and leukocyte recruitment in a naloxone-sensitive manner
The dose of 0.3 mg/kg of naloxone was ineffective in inhibiting the antinociceptive effect of quercetin in mechanical hyperalgesia test. On the other hand, the doses of 1 and 3 mg/kg of naloxone prevented since the 1st h the antinociceptive effect of quercetin (Fig. 3a) in MSU-induced mechanical hyperalgesia. The pre-treatment with naloxone also reversed the quercetin inhibition of MSU-induced recruitment of total leukocytes (Fig. 3b) and neutrophils (Fig. 3c) only at the dose of 3 mg/kg. Thus, the quercetin inhibition of MSU-induced mechanical hyperalgesia and leukocyte recruitment depends on activation of opioid receptors.

Quercetin inhibited MSU-induced mechanical hyperalgesia and leukocyte recruitment in an opioid-dependent manner. Treatment with naloxone (0.3–3 mg/kg/saline, i.p.) was performed 1 h before quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL, i.a.). The mechanical hyperalgesia (a) was evaluated 1–24 h after stimulus injection with an electronic pressure-meter test. The total of leukocytes (b) and neutrophils (c) counts was determined using Newbauer chamber and Rosenfelt stained slices 24 h after MSU injection. Results are presented as mean ± SEM of six mice per group per experiment, and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared to MSU, **P < 0.05 compared to quercetin group. ANOVA followed by Tukey’s test
Quercetin reduces the synovitis in articular joint-induced by gouty arthritis in a naloxone-sensitive manner
Quercetin inhibits MSU-induced synovitis in articular joint, decreasing the inflammatory infiltrate cells (Fig. 4a–f). In addition, quercetin effect was reversed by pre-treatment with naloxone at dose 3 mg/kg in a sensitive manner (Fig. 4f). The treatment with naloxone did not present effect per se.

Quercetin inhibits inflammatory infiltrate cells on synovial tissue in an opioid-dependent manner. Treatment with naloxone (3 mg/kg, i.p.) was performed 1 h before quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL, i.a.). The joint samples were collected at 24 h after MSU injection. Control (a), MSU (b), MSU and quercetin (c), MSU and quercetin and naloxone (d), and MSU and naloxone (e) and inflammatory infiltrate cells analysis/fields (f). All sections were stained with Hematoxylin and Eosin (magnification a–f, ×400) and the figure is representative of all experiment. Dimension used was 616 × 662 pixels for analysis (Field). Scale bars 50 µm. The arrow indicates representative infiltrate inflammatory cells counted. Results are presented as mean ± SEM of six mice per group per experiment, and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared to MSU group and **P < 0.05 compared with Quercetin group. ANOVA followed by Tukey’s t test
Quercetin inhibits MSU-induced TNF-α and IL-1β production in a naloxone-sensitive manner
Quercetin inhibited MSU-induced production of TNF-α (Fig. 5a) and IL-1β (Fig. 5b). In turn, naloxone pre-treatment prevented quercetin inhibition of MSU-induced TNF-α and IL-1β production in the knee joints at 24 h (Fig. 5a, b). The treatment with naloxone did not present effect per se.

Quercetin inhibited MSU-induced TNF-α and IL-1β production in joint in an opioid-dependent manner. Treatment with naloxone (3 mg/kg, i.p.) was performed 1 h before quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL, i.a.). The joint samples were collected at 24 h after MSU injection, and TNF-α (a) and IL-1β (b) levels were determined by ELISA method. Results are presented as mean ± SEM of six mice per group per experiment, and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared to MSU group and **P < 0.05 compared with Quercetin group. ANOVA followed by Tukey’s t test
Quercetin inhibits MSU-induced NFκB activation in a naloxone-sensitive manner
Quercetin inhibited MSU-induced NFκB activation as observed by an increase of total NFκB/phosphoNFκB ratio. In turn, naloxone treatment prevented quercetin inhibition of MSU-induced NFκB activation (Fig. 6). The treatment with naloxone did not present effect per se.

Pre-treatment with naloxone reversed quercetin inhibition of MSU-induced NFκB activation. Treatment with naloxone (3 mg/kg, i.p.) was performed 1 h before quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL, i.a.). The joint samples were collected at 24 h after MSU injection, and total NFκB/phospho NFB ratio (activation) was determined by ELISA method. Results are presented as mean ± SEM of six mice per group per experiment and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared to MSU group and **P < 0.05 compared with Quercetin group. ANOVA followed by Tukey’s t test
Quercetin inhibits MSU-induced NLRP3, ASC, Pro-caspase-1, and Pro-IL-1β mRNA expression in a naloxone-sensitive manner
Quercetin inhibited MSU-induced mRNA expression of the inflammasome components NLRP3, ASC, pro-caspase-1, and pro-IL-1β (Fig. 7a–d) in the knee joints at 24 h post-stimulus injection as determined by qPCR. On the other hand, naloxone treatment prevented quercetin inhibition of MSU-induced mRNA expression of inflammasome components.

Naloxone treatment reversed quercetin inhibition of MSU-induced NLRP3 inflammasome components mRNA expression. Treatment with naloxone (3 mg/kg/saline, i.p.) was performed 1 h before quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL). The NLRP3 (a), ASC (b), Pro-caspase-1 (c), and Pro-IL-1β (d) mRNA expression were evaluated 24 h after MSU injection. Results are presented as mean ± SEM of six mice per group per experiment and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared to MSU group and **P < 0.05 compared with Quercetin group. ANOVA followed by Tukey’s t test
Quercetin inhibits MSU-induced decrease of antioxidant capacity, superoxide anion production, and gp91phox mRNA expression in a naloxone-sensitive manner
Quercetin inhibited MSU-induced reduction of antioxidant defenses as observed by maintenance of GSH levels (Fig. 8a), ferric-reducing ability potential (FRAP assay—Fig. 8b), and free-radical scavenging ability (ABTS assay—Fig. 8c). Quercetin also inhibited MSU-induced NBT reduction (superoxide production—Fig. 8d) assessment and gp91phox mRNA expression (Fig. 8e) in the knee joints. In turn, quercetin effects were inhibited by naloxone treatment (Fig. 8a–e). The treatment with naloxone did not present effect per se.

Quercetin inhibited MSU-induced the decrease of antioxidant capacity, and superoxide production and gp91phox mRNA expression in an opioid-dependent manner. Treatment with naloxone (3 mg/kg/saline, i.p.) was performed 1 h before quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL). The GSH (a), FRAP (b), ABTS (c), NBT (d), and gp91phox mRNA expression (e) were evaluated 24 h after MSU injection. Results are presented as mean ± SEM of six mice per group per experiment, and are representative of two separated experiments. *P < 0.05 compared to saline group; # P < 0.05 compared to MSU group and **P < 0.05 compared with Quercetin group. ANOVA followed by Tukey’s t test
Quercetin induces Nrf2 and HO-1 mRNA expression in MSU-induced arthritis in a naloxone-sensitive manner
MSU stimulus did not alter Nrf2 (Fig. 9a) or HO-1 (Fig. 9b) mRNA expression in the knee joints. On the other hand, quercetin induced significant increase of Nrf2 (Fig. 9a) or HO-1 (Fig. 9b) mRNA expression, which was inhibited by naloxone treatment.

Quercetin inhibited MSU-induced Nrf2 and HO-1 mRNA expression in a naloxone-sensitive manner. Treatment with naloxone (3 mg/kg/saline, i.p.) was performed 1 h before Quercetin (100 mg/kg, s.c.) or vehicle (Tween 80 plus saline), and after additional 30 min, mice received MSU injection (100 µg/10 µL). The Nrf2 (a) and HO-1 (b) mRNA expression were evaluated 24 h after MSU injection. Results are presented as mean ± SEM of six mice per group per experiment, and are representative of two separated experiments. *P < 0.05 compared to saline group, # P < 0.05 compared to MSU group, **P < 0.05 compared with Quercetin group. ANOVA followed by Tukey’s t test
Quercetin inhibits MSU-induced IL-1β levels in BMDMs supernatant in gouty arthritis in a naloxone-sensitive manner in vitro
LPS priming (first signal) followed by MSU stimulation (second signal) induced an increase of IL-1β levels in BMDM, which was inhibited by quercetin in a dose-dependent manner (Fig. 10a). Quercetin at the concentration of 30 µM prevented inflammasome activation by MSU crystals decreasing IL-1β levels in BMDMs supernatant. In a similar way, morphine treatment also inhibited NLRP3 activating when stimulated with MSU crystals at the concentration of 20 µM (Fig. 10c). In turn, NLPR3 inflammasome stimulation in the presence of naloxone at concentration (20 µM) reversed the quercetin effect (Fig. 10b).

Quercetin inhibits MSU-induced IL-1β release by macrophages in a naloxone-sensitive manner. LPS (500 ng/ml)-primed BMDM were treated with quercetin 30 min before stimulation with MSU crystals (450 µg/mL) during 5 h. IL-1β concentration in the culture supernatant was measured by ELISA. a Concentration response of quercetin, b concentration response of naloxone added 1 h before quercetin treatment, and c concentration response of morphine. Results are mean ± SEM are representative of or more independent experiments. *P < 0.05 compared to saline group, # P < 0.05 compared to MSU group, **P < 0.05 compared with quercetin group. ANOVA followed by Tukey’s t test
Discussion
Monosodium urate (MSU) crystals activate phagocytes in the joints leading to the development of inflammatory response due to activation of NF-κB and NLRP3 inflammasome. These signaling pathways promote, for instance, the production of IL-1β resulting in the influx of neutrophils into the affected articular space. In fact, neutrophil recruitment is a major feature of acute gout arthritis provoking synovial membrane damage and release of lysosomal enzymes, elastase, oxygen-derived free radicals, chemotactic factors, and pro-inflammatory cytokines, which lead to enhanced inflammatory response and pain (Desaulniers et al. 2001; Cronstein and Terkeltaub 2006; Huang et al. 2012). The present data show that MSU-induced gout arthritis promotes mechanical hyperalgesia, leukocyte recruitment, TNF-α and IL-1β cytokine production, as well as NFκB activation, up regulation of NLRP3, ASC, Pro-caspase-1, and pro-IL-1β mRNA expression, and oxidative stress. All these MSU-induced physiopathological alterations in the joints were inhibited by the flavonoid quercetin in a naloxone-sensitive manner.
Quercetin presents analgesic effect, decreasing the pain threshold and leukocyte recruitment in varied pain models such as Ehrlich tumor-induced cancer pain (Calixto-Campos et al. 2015), streptozotocin-induced diabetic neuropathic pain (Anjaneyulu and Chopra 2003a), superoxide anion-induced pain (Maioli et al. 2015), lipopolysaccharide (LPS)-induced hyperalgesia (Anjaneyulu and Chopra 2003b), and carrageenan-induced hyperalgesia (Valério et al. 2009) and leukocyte recruitment (Souto et al. 2011). Therefore, the analgesic effect of quercetin is consistent among varied models. However, it was not known, to our knowledge, whether quercetin would have analgesic effect in the gout arthritis model. Quercetin treatment inhibited MSU-induced mechanical hyperalgesia at all time-points concomitantly with a reduction of leukocyte recruitment in a dose-dependent manner. Despite the description that recruited neutrophils contribute to inflammatory hyperalgesia (Cunha et al. 2008), the inhibition of hyperalgesia and neutrophil recruitment lined up with the dose of quercetin of 100 mg/kg, while the dose of 30 mg/kg inhibited only the MSU-induced hyperalgesia, which suggests that the inhibition of neutrophil recruitment and hyperalgesia are not fully interdependent. There is evidence that quercetin inhibits MSU-induced edema and leukocyte recruitment by inhibiting cytokine production and oxidative stress (Huang et al. 2012).
The previous studies identified that the analgesic effect of quercetin depends on endogenous opioids and, therefore, is reversed by the opioid receptor antagonist naloxone (Anjaneyulu and Chopra 2003a; Calixto-Campos et al. 2015; Maioli et al. 2015). However, it is unknown if this is the mechanism by which quercetin inhibits MSU-induced inflammation. In the present study, naloxone reversed, in a dose-dependent manner, the quercetin reduction of MSU-induced mechanical hyperalgesia and leukocyte recruitment. Opioids can act by three different mechanisms. First, opioids inhibit neuronal adenilate cyclase resulting in the reduction of PGE2-induced activation of adenilate cyclase and hyperalgesia; second, opioids act on peripheral nociceptor neurons activating the phosphatidylinositol 3-kinase (PI3K)g/protein kinase B (AKT)/neuronal nitric oxide synthase (nNOS)/nitric oxide (NO)/cyclic guanosine monophosphate-dependent protein kinase (PKG)/ATP-sensitive potassium channel (KATP) signaling inducing hyperpolarization of nociceptors (Cunha et al. 2010); and third, opioids inhibit cytokine production and the consequent hyperalgesia and neutrophil recruitment (Wang et al. 2005; Clark et al. 2007; Martin et al. 2010).
In experimental conditions related to opioid tolerance, opioids can enhance pain through increased TLR4 signaling and cytokine production (Ellis et al. 2016; Liang et al. 2016). On the other hand, in regular clinical conditions, opioids induce analgesia and inhibit inflammation. For instance, the κ-opioid receptor agonist U50,488H inhibited myocardial ischemia and reperfusion injury-induced TLR4 expression, NFκB activation, myeloperoxidase (MPO) activity, and TNF-α production (Cai et al. 2014). The μ opioid receptor agonist morphine reduces TLR4 expression by macrophages. In turn, the μ opioid receptor antagonist naltrexone increases macrophage TLR4 expression, which indicates endogenous role of μ opioid receptors in regulating TLR4 expression (Franchi et al. 2012). Thus, opioid receptor activation has been related to consistent anti-inflammatory effects.
The anti-inflammatory and analgesic effects of quercetin depend on inhibiting NFκB activation (Chen et al. 2005; Vicentini et al. 2011; Borghi et al. 2016), which corroborates the quercetin inhibition of MSU-induced cytokine production in rats and mice (Huang et al. 2012). Nevertheless, our data expand these findings demonstrating that quercetin inhibition of NFκB activation is naloxone-sensitive and is an underlying mechanism by which quercetin reduced MSU-induced inflammation and pain. As a result of NFκB inhibition, quercetin inhibited MSU-induced NLRP3, ASC, pro-caspase-1, and pro-IL-1β mRNA expression.
In BMDM primed with LPS, the challenge with MSU induces IL-1β release in the supernatant, which was inhibited by quercetin in a naloxone-sensitive manner. The in vitro system used in the present study allows to determine whether after signal 1 activation with LPS, a test drug can inhibit signal 2 that results in IL-1β maturation and release in the supernatant of cell culture (Martinon et al. 2006). The present data show that quercetin inhibits both signal 1 and 2, which explains the major anti-inflammatory and analgesic effect of quercetin in MSU-induced arthritis. Quercetin also inhibited NLRP3 inflammasome components expression and decreased IL-1β production in models of hepatic inflammation (Zhang et al. 2015), carrageenan-induced paw inflammation (Valério et al. 2009), renal injury (Hu et al. 2012), and spinal cord injury (Zhou et al. 2010; Jiang et al. 2016), although these studies did not investigate whether quercetin would be able to inhibit signal 2 as performed in the present work. In agreement with the present data, it has recently been demonstrated that quercetin inhibits ATP, nigericin, alum (NLRP3 activators), and double strand DNA (AIM2 activator)-induced inflammasome activation by diminishing ASC oligomerization (Domiciano et al. 2017).
Reactive oxygen species (ROS) contribute to acute and chronic inflammation (Rasool and Varalakshmi 2006). MSU-induced arthritis promoted oxidative stress observed as decrease of GSH levels, FRAP and ABTS activities, and increase of NBT reduction (superoxide anion production) and gp91phox mRNA expression. Quercetin inhibited MSU-induced oxidative stress in a naloxone-sensitive manner. Quercetin has inherent antioxidant chemical groups that can explain its antioxidant effects in vitro and in vivo (Verri et al. 2012). This flavonoid inhibits ROS such as hydrogen peroxide (H2O2) (Boumaza et al. 2016) and decreases gp91phox gene expression (Sun et al. 2015b; Borghi et al. 2016) in models of inflammation, and also attenuates lipid peroxidation and increases antioxidant enzymes in gout arthritis models in rats (Huang et al. 2012). Importantly, quercetin acting on oxidative stress sensitive signaling pathways induces Nrf2 and HO-1 gene expression (Yao et al. 2007; Liu et al. 2015) enhancing the expression of multiple antioxidant enzymes such as SOD and GSH, thus decreasing oxidative stress (Jung and Kwak 2010; Bryan et al. 2013), which contributes to the resolution of oxidative stress damages. The sensitivity to naloxone treatment suggests that the antioxidant effect of quercetin is not solely dependent on its antioxidant chemical groups in the MSU-induced arthritis. In fact, quercetin treatment induced an increase of Nrf2 and its downstream target HO-1 mRNA expression. Nrf2 also has an effect in the increase of GSH levels. Moreover, naloxone inhibited quercetin induction of Nrf2 and HO-1 mRNA expression. Thus, in addition to the quercetin antioxidant effects dependent on its chemical groups, triggering an endogenous opioid system might account to the antioxidant effect of quercetin in reducing the MSU-induced inflammasome activation and arthritis.
The opioid receptors have been reported to present crucial role in oxidative stress in several models by acting on inhibition of H2O2 in vitro (Sazonova et al. 2016), inducing gastric protection (Ribeiro et al. 2016) and reducing NSAID-induced oxidative stress (Yin et al. 2015). Opioid receptor agonists have potent antioxidant effects by inhibiting the generation of ROS such as inducible nitric oxide synthase (iNOS) by modulation of sigma-1 receptor (σ1R) in lipopolysaccharide (LPS)-stimulated BV2 microglia (Wu et al. 2015). In line with the current findings and contribution of the opioid system to quercetin effects, quercetin inhibits interstitial cells of Cajal (ICCs) pacemaker activities via opioid receptors in cultured murine ICCs and decreases oxidative stress (Gim et al. 2015). Other flavonoids have been reported to reduce oxidative stress by acting on opioid receptors such as kappa and delta-opioid receptors (Meotti 2005; Donato et al. 2014; Rosales et al. 2014).
The δ-opioid receptor mediated the increase in Nrf2 translocation promoting a cytoprotection against hypoxic injury in vitro, reducing the oxidative molecules and oxidative damage of cells (Cao et al. 2015). Activation of other opioid receptors such as σ1R protects against cellular oxidative stress and activates antioxidant responsive elements (ARE), and increases the levels of the antioxidant protein peroxiredoxin 6 (Prdx6), endoplasmic reticulum (ER) chaperone BiP (GRP78), NAD(P)H quinone oxidoreductase 1 (NQO1), and superoxide dismutase 1 (SOD1) mRNA expression in COS cells (Pal et al. 2012). Thusly, it is well established that antioxidant response element (ARE) activation triggers the transcription of genes involved in antioxidant defense by a mechanism involving the Nrf2 and KEAP1 pathway (Muthusamy et al. 2012; Gan and Johnson 2014). In models of retinal disease, the σ1R increased Nrf2 mRNA expression and decrease oxidative stress in Müller glia cells, thereby demonstrating a major contribution of σ1R as a mediator of the key antioxidant pathway Nrf2/Keap1 (Wang et al. 2015) and endoplasmic reticulum (ER) stress genes (BiP/GRP78, Atf6, Atf4, and Ire1α) (Ha et al. 2014).
It is noteworthy to mention that in the Ehrlich tumor-induced pain model in mice, quercetin inhibited tumor-induced hyperalgesia, paw skin myeloperoxidase activity (an indirect marker of neutrophil and macrophage recruitment), oxidative stress, and cytokine production. However, naloxone treatment reversed only the analgesic effect of quercetin (Calixto-Campos et al. 2015). A likely explanation for this lack of effect of naloxone contrasting with the present data is that quercetin treatment was performed during 8 days, whilst naloxone treatment was performed only at the 8th day post-tumor inoculation (Calixto-Campos et al. 2015), and in the present study, both quercetin and naloxone were administrated once. Furthermore, the dose of naloxone used in the Ehrlich tumor model was threefold lower than in the present study. These differences in treatment protocols (number of treatments and doses) in addition to the differences in the physiopathology of the experimental models (tumor-induced pain versus innate immune response-induced pain) and time-course of disease might explain the contrasting results.
In conclusion, the present data demonstrate that quercetin inhibits MSU-induced gout arthritis diminishing mechanical hyperalgesia and leukocyte recruitment. These anti-inflammatory and analgesic effects of quercetin were accompanied by inhibition of NFκB activation, which is a likely explanation for the reduction of TNF-α and IL-1β production as well as inflammasome components and gp91phox mRNA expression. Furthermore, quercetin inhibited MSU-induced IL-1β release by LPS-primed macrophages, which suggests that this flavonoid inhibits NRLP3 inflammasome activation. Quercetin also induced Nrf2 and HO-1 mRNA expression that contributed to the antioxidant effect. The analgesic and anti-inflammatory effects of quercetin were sensitive to naloxone treatment, suggesting that quercetin induces endogenous opioid-dependent mechanisms regulating NFκB and Nrf2 transcription factors-dependent pathways as well as inflammasome activation. In fact, morphine was also capable of inhibiting MSU-induced IL-1β release by LPS-primed macrophages. Therefore, quercetin deserves further investigation as a potential treatment for acute gout arthritis.
References
Anjaneyulu M, Chopra K (2003a) Quercetin, a bioflavonoid, attenuates thermal hyperalgesia in a mouse model of diabetic neuropathic pain. Prog Neuropsychopharmacol Biol Psychiatry 27:1001–1005
Anjaneyulu M, Chopra K (2003b) Reversal of lipopolysaccharide-induced thermal and behavioural hyperalgesia by quercetin. Drug Dev Res 58:248–252
Borghi SM, Carvalho TT, Staurengo-Ferrari L et al (2013) Vitexin inhibits inflammatory pain in mice by targeting TRPV1, oxidative stress, and cytokines. J Nat Prod 76:1141–1149
Borghi SM, Pinho-Ribeiro FA, Fattori V et al (2016) Quercetin inhibits peripheral and spinal cord nociceptive mechanisms to reduce intense acute swimming-induced muscle pain in mice. PLoS One 11:e0162267
Boumaza S, Belkebir A, Neggazi S et al (2016) Therapeutic role of resveratrol and quercetin on aortic fibroblasts of psammomys obesus after oxidative stress by hydrogen peroxide. Am J Ther 0:1–15
Bryan HK, Olayanju A, Goldring CE, Park BK (2013) The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem Pharmacol 85:705–717
Cai W, Zhang Y, Li J, Wang J (2014) κ-opioid receptor agonist U50, 488H attenuates myocardial ischemia-reperfusionvia modulating Toll-like receptor 4/nuclear factor-κB signaling in rat. Zhonghua Xin Xue Guan Bing Za Zhi 42:866–872
Calixto-Campos C, Corrêa MP, Carvalho TT et al (2015) Quercetin reduces Ehrlich tumor-induced cancer pain in mice. Anal Cell Pathol 2015:1–18
Cao S, Chao D, Zhou H et al (2015) A novel mechanism for cytoprotection against hypoxic injury: δ-opioid receptor-mediated increase in Nrf2 translocation. Br J Pharmacol 172:1869–1881
Chen J-C, Ho F-M, Chao Pei-Dawn Lee et al (2005) Inhibition of iNOS gene expression by quercetin is mediated by the inhibition of IkappaB kinase, nuclear factor-kappa B and STAT1, and depends on heme oxygenase-1 induction in mouse BV-2 microglia. Eur J Pharmacol 521:9–20
Clark JD, Shi X, Li X et al (2007) Morphine reduces local cytokine expression and neutrophil infiltration after incision. Mol Pain 3:28
Cronstein BN, Terkeltaub R (2006) The inflammatory process of gout and its treatment. Arthritis Res Ther 8(Suppl 1):S3
Cunha TM, Verri WA, Schivo IR et al (2008) Crucial role of neutrophils in the development of mechanical inflammatory hypernociception. J Leukoc Biol 83:824–832
Cunha TM, Roman-Campos D, Lotufo CM et al (2010) Morphine peripheral analgesia depends on activation of the PI3K/AKT/nNOS/NO/KATP signaling pathway. Proc Natl Acad Sci 107:4442–4447
de Souza MR, de Paula CA, Pereira de Resende ML et al (2012) Pharmacological basis for use of Lychnophora trichocarpha in gouty arthritis: anti-hyperuricemic and anti-inflammatory effects of its extract, fraction and constituents. J Ethnopharmacol 142:845–850
Desaulniers P, Fernandes M, Gilbert C et al (2001) Crystal-induced neutrophil activation. VII. Involvement of Syk in the responses to monosodium urate crystals. J Leukoc Biol 70:659–668
Dhanasekar C, Kalaiselvan S, Rasool M (2015) Morin, a bioflavonoid suppresses monosodium urate crystal-induced inflammatory immune response in RAW 264.7 macrophages through the inhibition of inflammatory mediators, intracellular ROS levels and NF-κB activation. PLoS One 10:e0145093
Domiciano TP, Wakita D, Jones HD et al (2017) Quercetin inhibits inflammasome activation by interfering with ASC oligomerization and prevents interleukin-1 mediated mouse vasculitis. Sci Rep 7:41539
Donato F, de Gomes MG, Goes ATR et al (2014) Hesperidin exerts antidepressant-like effects in acute and chronic treatments in mice: possible role of l-arginine-NO-cGMP pathway and BDNF levels. Brain Res Bull 104:19–26
Dubchak N, Falasca GF (2010) New and improved strategies for the treatment of gout. Int J Nephrol Renovasc Dis 3:145–166
Ellis A, Grace PM, Wieseler J et al (2016) Morphine amplifies mechanical allodynia via TLR4 in a rat model of spinal cord injury. Brain Behav Immun 58:348–356
Franchi S, Moretti S, Castelli M et al (2012) Mu opioid receptor activation modulates Toll like receptor 4 in murine macrophages. Brain Behav Immun 26:480–488
Gan L, Johnson JA (2014) Oxidative damage and the Nrf2-ARE pathway in neurodegenerative diseases. Biochim Biophys Acta 1842:1208–1218
Gim H, Nam JH, Lee S et al (2015) Quercetin inhibits pacemaker potentials via nitric oxide/cGMP-dependent activation and TRPM7/ANO1 channels in cultured interstitial cells of cajal from mouse small intestine. Cell Physiol Biochem 35:2422–2436
Guazelli CFS, Fattori V, Colombo BB et al (2013) Quercetin-loaded microcapsules ameliorate experimental colitis in mice by anti-inflammatory and antioxidant mechanisms. J Nat Prod 76:200–208
Guerrero ATGG, Verri WA, Cunha TM et al (2006) Hypernociception elicited by tibio-tarsal joint flexion in mice: a novel experimental arthritis model for pharmacological screening. Pharmacol Biochem Behav 84:244–251
Ha Y, Shanmugam AK, Markand S et al (2014) Sigma receptor 1 modulates ER stress and Bcl2 in murine retina. Cell Tissue Res 356:15–27
Hu Q-H, Zhang X, Pan Y et al (2012) Allopurinol, quercetin and rutin ameliorate renal NLRP3 inflammasome activation and lipid accumulation in fructose-fed rats. Biochem Pharmacol 84:113–125
Huang J, Zhu M, Tao Y et al (2012) Therapeutic properties of quercetin on monosodium urate crystal-induced inflammation in rat. J Pharm Pharmacol 64:1119–1127
Jiang W, Huang Y, Han N et al (2016) Quercetin suppresses NLRP3 inflammasome activation and attenuates histopathology in a rat model of spinal cord injury. Spinal Cord 54:592–596
Julius D (2013) TRP channels and pain. Annu Rev Cell Dev Biol 29:355–384
Jung K-A, Kwak M-K (2010) The Nrf2 system as a potential target for the development of indirect antioxidants. Molecules 15:7266–7291
Khabbazi S, Goumon Y, Parat M-O (2015) Morphine modulates interleukin-4- or breast cancer cell-induced pro-metastatic activation of macrophages. Sci Rep 5:11389
Khanna D, Fitzgerald JD, Khanna PP et al (2012) 2012 American College of Rheumatology guidelines for management of gout. Part 1: Systematic nonpharmacologic and pharmacologic therapeutic approaches to hyperuricemia. Arthritis Care Res (Hoboken) 64:1431–1446
Kim Y-J, Park W (2016) Anti-inflammatory effect of quercetin on RAW 264.7 mouse macrophages induced with polyinosinic-polycytidylic acid. Molecules 21:450
Liang Y, Chu H, Jiang Y, Yuan L (2016) Morphine enhances IL-1β release through toll-like receptor 4-mediated endocytic pathway in microglia. Purinergic Signal 12:637–645
Liu C-M, Ma J-Q, Xie W-R et al (2015) Quercetin protects mouse liver against nickel-induced DNA methylation and inflammation associated with the Nrf2/HO-1 and p38/STAT1/NF-κB pathway. Food Chem Toxicol 82:19–26
Liu D, Cao G, Han L et al (2016) Flavonoids from Radix Tetrastigmae inhibit TLR4/MD-2 mediated JNK and NF-κB pathway with anti-inflammatory properties. Cytokine 84:29–36
Lu F, Liu L, Yu D et al (2014) Therapeutic effect of Rhizoma Dioscoreae Nipponicae on gouty arthritis based on the SDF-1/CXCR 4 and p38 MAPK pathway: an in vivo and in vitro study. Phyther Res 28:280–288
Maioli NAA, Zarpelon ACC, Mizokami SSS et al (2015) The superoxide anion donor, potassium superoxide, induces pain and inflammation in mice through production of reactive oxygen species and cyclooxygenase-2. Braz J Med Biol Res 48:321–331
Manchope MF, Calixto-Campos C, Coelho-Silva L et al (2016) Naringenin inhibits superoxide anion-induced inflammatory pain: role of oxidative stress, cytokines, Nrf-2 and the NO–cGMP–PKG–KATPChannel signaling pathway. PLoS One 11:e0153015
Martin WJ, Walton M, Harper J (2009) Resident macrophages initiating and driving inflammation in a monosodium urate monohydrate crystal-induced murine peritoneal model of acute gout. Arthritis Rheum 60:281–289
Martin JL, Koodie L, Krishnan AG et al (2010) Chronic morphine administration delays wound healing by inhibiting immune cell recruitment to the wound site. Am J Pathol 176:786–799
Martinez RM, Pinho-Ribeiro FA, Steffen VS et al (2015a) Hesperidin methyl chalcone inhibits oxidative stress and inflammation in a mouse model of ultraviolet B irradiation-induced skin damage. J Photochem Photobiol B Biol 148:145–153
Martinez RM, Pinho-Ribeiro FA, Steffen VS et al (2015b) Naringenin inhibits UVB irradiation-induced inflammation and oxidative stress in the skin of hairless mice. J Nat Prod 78:1647–1655
Martinon F, Pétrilli V, Mayor A et al (2006) Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 440:237–241
McGettrick AF, O’Neill LAJ (2013) How metabolism generates signals during innate immunity and inflammation. J Biol Chem 288:22893–22898
Meotti FC (2005) Analysis of the antinociceptive effect of the flavonoid myricitrin: evidence for a role of the l-arginine-nitric oxide and protein kinase C pathways. J Pharmacol Exp Ther 316:789–796
Muthusamy VR, Kannan S, Sadhaasivam K et al (2012) Acute exercise stress activates Nrf2/ARE signaling and promotes antioxidant mechanisms in the myocardium. Free Radic Biol Med 52:366–376
Neuschäfer-Rube F, Pathe-Neuschäfer-Rube A, Hippenstiel S et al (2013) NF-κB-dependent IL-8 induction by prostaglandin E2 receptors EP1 and EP4. Br J Pharmacol 168:704–717
Nishimura A, Akahoshi T, Takahashi M et al (1997) Attenuation of monosodium urate crystal-induced arthritis in rabbits by a neutralizing antibody against interleukin-8. J Leukoc Biol 62:444–449
Okamoto T (2005) Safety of quercetin for clinical application (review). Int J Mol Med 16:275–278
Pal A, Fontanilla D, Gopalakrishnan A et al (2012) The sigma-1 receptor protects against cellular oxidative stress and activates antioxidant response elements. Eur J Pharmacol 682:12–20
Pinho-Ribeiro FA, Hohmann MSNN, Borghi SM et al (2015) Protective effects of the flavonoid hesperidin methyl chalcone in inflammation and pain in mice: role of TRPV1, oxidative stress, cytokines and NF-κB. Chem Biol Interact 228:88–99
Pinho-Ribeiro FA, Zarpelon AC, Fattori V et al (2016) Naringenin reduces inflammatory pain in mice. Neuropharmacology 105:508–519
Rasool M, Varalakshmi P (2006) Suppressive effect of Withania somnifera root powder on experimental gouty arthritis: an in vivo and in vitro study. Chem Biol Interact 164:174–180
Ribeiro KA, Chaves HV, Filho SMP et al (2016) Αlpha-2 adrenergic and opioids receptors participation in mice gastroprotection of Abelmoschus esculentus lectin. Curr Pharm Des 22:4736–4742
Rosales MAB, Silva KC, Duarte DA et al (2014) Endocytosis of tight junctions caveolin nitrosylation dependent is improved by cocoa via opioid receptor on RPE cells in diabetic conditions. Investig Opthalmol Vis Sci 55:6090
Rukdee N, Rojsanga P, Phechkrajang CM (2015) Development and validation of LC-MS/MS method for quantitative determination of adenosine, guanosine, xanthine and uric acid in widely consumed vegetables in Thailand. Nat Prod Commun 10:1435–1437
Sabina EP, Nagar S, Rasool M (2011) A role of piperine on monosodium urate crystal-induced inflammation—an experimental model of gouty arthritis. Inflammation 34:184–192
Sazonova EN, Samarina EY, Lebed’ko OA et al (2016) Cytoprotective effect of peptide sedatin, an agonist of μ/δ-opioid receptors, on primary culture of pulmonary fibroblasts of albino rats under conditions of oxidative stress. Bull Exp Biol Med 161:41–44
Sedlak J, Lindsay RH (1968) Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal Biochem 25:192–205
Souto FO, Zarpelon AC, Staurengo-Ferrari L et al (2011) Quercetin reduces neutrophil recruitment induced by CXCL8, LTB 4, and fMLP: inhibition of actin polymerization. J Nat Prod 74:113–118
Staurengo-Ferrari L, Mizokami SS, Fattori V et al (2014) The ruthenium nitric oxide donor, [Ru(HEDTA)NO], inhibits acute nociception in mice by modulating oxidative stress, cytokine production and activating the cGMP/PKG/ATP-sensitive potassium channel signaling pathway. Naunyn Schmiedebergs Arch Pharmacol 387:1053–1068
Sun GY, Chen Z, Jasmer KJ et al (2015a) Quercetin attenuates inflammatory responses in BV-2 microglial cells: role of MAPKs on the Nrf2 pathway and induction of heme oxygenase-1. PLoS One 10:e0141509
Sun X, Yamasaki M, Katsube T, Shiwaku K (2015b) Effects of quercetin derivatives from mulberry leaves: improved gene expression related hepatic lipid and glucose metabolism in short-term high-fat fed mice. Nutr Res Pract 9:137
Sun G-W, Qiu Z-D, Wang W-N et al (2016) Flavonoids extraction from propolis attenuates pathological cardiac hypertrophy through PI3K/AKT signaling pathway. Evid Based Complement Altern Med 2016:1–11
Umamaheswari M, Asokkumar K, Sivashanmugam AT et al (2009) In vitro xanthine oxidase inhibitory activity of the fractions of Erythrina stricta Roxb. J Ethnopharmacol 124:646–648
Valério DA, Georgetti SR, Magro DA et al (2009) Quercetin reduces inflammatory pain: inhibition of oxidative stress and cytokine production. J Nat Prod 72:1975–1979
Verri WA, Cunha TM, Parada CA et al (2006) Hypernociceptive role of cytokines and chemokines: targets for analgesic drug development? Pharmacol Ther 112:116–138
Verri WA, Guerrero ATG, Fukada SY et al (2008) IL-33 mediates antigen-induced cutaneous and articular hypernociception in mice. Proc Natl Acad Sci USA 105:2723–2728
Verri WA, Souto FO, Vieira SM et al (2010) IL-33 induces neutrophil migration in rheumatoid arthritis and is a target of anti-TNF therapy. Ann Rheum Dis 69:1697–1703
Verri WA, Vicentini FTMC, Baracat MM et al (2012) Flavonoids as anti-inflammatory and analgesic drugs: mechanisms of action and perspectives in the development of pharmaceutical forms. In: Rahman A (ed) Studies in natural products chemistry. Elsevier, Amsterdam, pp 297–330
Vicentini FTMC, He T, Shao Y et al (2011) Quercetin inhibits UV irradiation-induced inflammatory cytokine production in primary human keratinocytes by suppressing NFkB pathway. J Dermatol Sci 61:162–168
Wang J, Barke RA, Charboneau R, Roy S (2005) Morphine impairs host innate immune response and increases susceptibility to Streptococcus pneumoniae lung infection. J Immunol 174:426–434
Wang C, Pan Y, Zhang Q-Y et al (2012) Quercetin and allopurinol ameliorate kidney injury in STZ-treated rats with regulation of renal NLRP3 inflammasome activation and lipid accumulation. PLoS One 7:e38285
Wang J, Shanmugam A, Markand S et al (2015) Sigma 1 receptor regulates the oxidative stress response in primary retinal Müller glial cells via NRF2 signaling and system xc−, the Na+-independent glutamate–cystine exchanger. Free Radic Biol Med 86:25–36
Wu Z, Li L, Zheng L-T et al (2015) Allosteric modulation of sigma-1 receptors by SKF83959 inhibits microglia-mediated inflammation. J Neurochem 134:904–914
Xu L, Liu S, Guan M, Xue Y (2016) Comparison of prednisolone, etoricoxib, and indomethacin in treatment of acute gouty arthritis: an open-label, randomized, controlled trial. Med Sci Monit 22:810–817
Yao P, Nussler A, Liu L et al (2007) Quercetin protects human hepatocytes from ethanol-derived oxidative stress by inducing heme oxygenase-1 via the MAPK/Nrf2 pathways. J Hepatol 47:253–261
Yin H, Cai H-Z, Wang S-K et al (2015) Wheat peptides reduce oxidative stress and inhibit NO production through modulating μ-opioid receptor in a rat NSAID-induced stomach damage model. Chin J Nat Med 13:22–29
Zarpelon AC, Cunha TMT, Alves-Filho JC et al (2013) IL-33/ST2 signalling contributes to carrageenin-induced innate inflammation and inflammatory pain: role of cytokines, endothelin-1 and prostaglandin E2. Br J Pharmacol 169:90–101
Zhang X, Zhang J-H, Chen X-Y et al (2015) Reactive oxygen species-induced TXNIP drives fructose-mediated hepatic inflammation and lipid accumulation through NLRP3 inflammasome activation. Antioxid Redox Signal 22:848–870
Zhao J, Huang Y (2015) Salivary uric acid as a noninvasive biomarker for monitoring the efficacy of urate-lowering therapy in a patient with chronic gouty arthropathy. Clin Chim Acta 450:115–120
Zhou R, Tardivel A, Thorens B et al (2010) Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat Immunol 11:136–140
Zimmermann M (1983) Ethical guidelines for investigations of experimental pain in conscious animals. Pain 16:109–110
Acknowledgements
This work was supported by Brazilian grants from Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Decit/SCTIE/MS through CNPq with the support of Fundação Araucária and SESA-PR, and Parana State Government. Larissa Staurengo-Ferrari received a post-doctoral fellowship from CNPq (Process: 161279/2015) and Talita P. Domiciano received a post-doctoral fellowship from CAPES/Fundação Araucária (Process: 096/2014).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Ruiz-Miyazawa, K.W., Staurengo-Ferrari, L., Mizokami, S.S. et al. Quercetin inhibits gout arthritis in mice: induction of an opioid-dependent regulation of inflammasome. Inflammopharmacol 25, 555–570 (2017). https://doi.org/10.1007/s10787-017-0356-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10787-017-0356-x