
Abstract
Where biodiversity conservation and environmental preservation are significant concerns, rapid assessment and monitoring of the colonization and spread of non-native species are essential for timely decision-making and response. We developed and evaluated the adequacy of a rapid assessment protocol (RAP) for detecting non-native fish species in 74 Eastern Brazilian lakes. The RAP consists of a single field day employing two surveyors to conduct interviews with local sport fishers, visual surveys of fish, angling with lures, and gillnetting. We compared our results with those from separate, intense, large sampling effort (LSE) field surveys. Despite requiring less than 1/100th of the field effort, the RAP was able to detect the presence of most non-native fish species that were reported in the same lakes by LSE surveys. Information from local sport fishers was invaluable, particularly for detecting species that were in low abundance, but was not available for lakes within a forest preserve area. Non-native introductions commonly lead to rapid and widespread invasion and adverse effects on native biota. Our results strongly suggest that the RAP could function as a cost-effective tool for efficiently assessing the presence of non-native fishes in lakes and monitoring their colonization and spread over large geographic areas.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rapid Assessment Protocols or Programs (RAP methods) are fast sampling methods used to survey biodiversity in potential habitats containing large numbers of endemic species, permitting a quick evaluation of the biodiversity or specific taxonomic or functional groups in specific regions (Lodh & Agarwala, 2016; Madalozzo et al., 2017; Sant et al., 2017). Despite its use in biodiversity surveys, the fast sampling, inherent to RAP methods, may be valuable to quickly identify invasions in some systems, for instance, in those occupied by non-native fish. Although RAP is thought to diminish the time and costs involved in biodiversity surveys, this method can produce inaccurate data because of low sampling effort. Good sampling designs that have a suitable number of representative and independent samples are needed to obtain reasonable parameter estimates (Krebs, 1999). The number of species identified depends on sampling effort (Harrison & Martinez, 1995), and the RAP efficacy may depend on the abilities of the researcher (Metzeling et al., 2003).
Reduced funding availability for biodiversity conservation, mainly in developing countries with a potential for quick degradation of natural resources, makes it essential to identify priorities for biodiversity protection (Douglass et al., 2011; Wu et al., 2014). In this context, RAP techniques may receive an increasing global interest, considering the continuous dispersion of non-native species worldwide (Simberloff et al., 2013) and the scarcity of funds available to deal with the problem.
With increasing tourism and commercial activities, organism invasions have become present in altered and preserved environments across the world (Harwood & Parjulee, 2010). As in Africa (Marr et al., 2017), North America (MacIsaac et al., 2015), and Europe (Nunes et al., 2015), in Brazil, lakes of the River Doce basin were invaded by non-native fish species (Lima et al., 2010) what caused local extinction of many native fish species (Latini & Petrere, 2007; Giacomini et al., 2011). Among these non-native species are the peacock bass Cichla ocellaris Bloch & Schneider, 1801 and the red piranha Pygocentrus nattereri Kner, 1858 (Latini & Petrere, 2004). The lakes are dispersed in a region of more than 100,000 ha and may be used as an exceptional case to test RAP techniques, considering, application worldwide. As in many regions worldwide, the River Doce basin is compounded by the fact that the relevant conservation decision makers have little time to study the system, as they race to prevent more local fish extinctions. Nevertheless, a successful management action against non-native fish invasions requires the identification of which non-native species occurs in which lake. So, RAP techniques may be useful tools for unveiling the real dimension of the problem in the River Doce basin, and, in other regions worldwide.
In this study, we tested the hypothesis that a RAP method (with a low sampling effort) may produce the same results of a large sampling effort (LSE) method surveying non-native fish in lakes.
Methods
Study area
This study was conducted in the State of Minas Gerais, the Eastern region (Marliéria, Timóteo, and Dionísio districts). Lakes in the river basin are located between 42°38′–48°28′W and 19°41′–19°30′S, with an altitude between 236 and 515 m AMSL. The climate in the region is tropical and partly humid with a rainy season from November to March and a dry season lasting 4–5 months, usually from May to September (Nimer, 1989). Two adjacent and contrasting areas were studied: (i) a forest reserve in the Parque Estadual do Rio Doce (PERD) (~ 36,000 ha) and (ii) an economically important eucalyptus plantation managed by Companhia Agrícola Florestal Santa Bárbara (CAF) (~ 25,000 ha). In the PERD area, fishing is only permitted for scientific purposes. Sampled lakes included about 30% of the 149 lakes in the region with 32 in the PERD area and 24 in the CAF area (Fig. 1). Two sampling methods were used in all of these lakes: the first was characterized by a low sampling effort used in a short time interval in all lakes (this is called here as the RAP method); the second was the LSE method employed in ten lakes in the CAF area and six lakes in the PERD area. The LSE method was used to compare and test the efficiency of the RAP method.

Picture showing PERD (Parque Estadual do Rio Doce) and CAF (Companhia Agrícola Florestal) areas at River Doce Basin, Brazil. White spots represent lakes, and some of them are numbered corresponding to used in the results of this study (see Table 1)
The RAP method
Different fish species respond differently to distinct fishing methods (Sutherland, 2001). So, to survey non-native fish species, multiple techniques were used to increase the probability of detecting fish and to make efficient use of the available time between February and May 2002, totaling 38 sampling days. The techniques used were interviews with sport fishers (whenever possible), visual detection, fishing with lures, and gillnetting. All techniques were employed at the same time in each lake.
Non-structured interviews with sport fishers were performed with the purpose of obtaining information about the non-native fish present in the lakes. The number of people interviewed differed among lakes: some lakes, being more attractive, apparently had more sport fishers. At least one interview per lake was conducted. Most of the times, however, only one or two interviews per lake were carried out. The reports of non-native species provided by sport fishers were considered only when the fishers could authenticate the claim showing a captured fish. This technique was applied only in lakes in the CAF area because in the PERD area there were no fishermen.
The visual search for non-native fish species was performed only in the littoral zone of each lake, using aquatic macrophytes banks, and in zones with logs and trunks where some of the non-native fish tend to make their nests (e.g., peacock bass and oscar Astronotus ocellatus (Agassiz, 1831)). In all lakes, the same two observers searched for 30 min (observing from above water surface) for non-native species which were marked as present if they were detected by at least one of the two observers.
Fishing with lures was also performed in all lakes. The lures used were the top-water Yo-Zuri Suspending Crystal Minnows, 51/4″, 3/4 oz. with blue/silver color and the subsurface Stamina Peacock Minnow, 7″, 1¼ oz. with silver/olive color. Fish were sampled by casting lures for 30 min at each of several locations around the shoreline. Non-native species were recorded after each catch.
Gillnets were deployed, using nets each measuring 10 m × 1.6 m with six different mesh sizes (15, 20, 30, 40, 50 , and 60 mm between adjacent knots). These gillnets were joined in a set and remained in the lakes for 30 min. Whenever possible, these were used in shoreline zones with different habitats (e.g., aquatic macrophyte banks, swamps, beaches) for a more representative sampling. Gillnetting had a fishing effort of only 30 m.h for each sampled lake (obtained by multiplication of net length by number of nets by time in hours; effort = 10 × 6 × 0.5 = 30 m.h). If a catch of at least one specimen of non-native species was recorded, this species was considered as present in the lake.
Lakes were characterized as being free of non-natives only if no non-native species were detected with any of the multiple RAP techniques. The presence of one non-native fish specimen from at least one of these four techniques led us to classify the lake as an invaded lake.
The LSE method
The LSE (large sampling effort) method consisted of bimonthly sampling in ten lakes in the CAF area, from July 2002 until May 2003, with six samples in each lake to increase the chance of sampling non-native species due to possible seasonal variation in some populations. In this method, three sets of gill nets were used consisting of six different mesh sizes, each set (15, 20, 30, 40, 50, and 60 mm between adjacent knots) that measured 10 m × 1.6 m (except for the 60 mm gill net, which had a 20 m length).
The gill nets were placed at shorelines for 3 h, beginning between 5 and 6 p.m. (including daylight, twilight, and night periods). The location of the first net of each set was chosen by lot in first sampling and kept in the same coordinates during the other samples. Other nets were placed in a sequence of this distance by 8–10 m. The LSE method produced a total effort of 3,780 m.h (obtained by multiplying the number of samples by the number of sets by net length by the number of nets by time in hours; Effort10m gillnets = 6 × 3 × 10 × 5 × 3 = 2,700 m.h + Effort20m gillnets = 6 × 3 × 20 × 1 × 3 = 1,080 m.h so Total Effort = 3,780 m.h), 126 times larger than the effort applied by the RAP method.
Latini & Petrere’s (2004) study of non-native species effects on native community used a massive fishing effort in six lakes in the PERD area (Gambá, Gambazinho, Águas Claras, Azul, Bonita, and Lagoinha lakes). In that study, fishing was performed between May and August 2000, and the fishing effort was standardized using 6 gill nets with 13 different mesh sizes (15, 20, 30, 40, 50, 60, 70, 80, 90, 100, and 110 mm between adjacent knots) with 10 m × 1.6 m gill nets. Gill nets were employed in areas both with and without aquatic macrophytes for 16 h, producing an effort of 12,480 m.h, 416 times larger than the effort expended by the RAP method. All sampled fish in the RAP and the LSE methods were fixed in 10% formalin and identified in the laboratory with the appropriate keys (Géry, 1977; Nelson, 1994; Britski et al., 1999).
Statistical analysis
To test the RAP method efficacy in detecting non-native fish species, the presence of each non-native species obtained by the RAP was compared with its abundance using the LSE method. Such comparison permitted the analysis of detection power for each non-native species. To perform this analysis, a logistic regression model (Hosmer & Lemeshow, 1989) was used as a detector of the relationship between the categorical binary variable obtained from the RAP (presence [1 value] or absence [0 value] of non-native fish) and the quantitative continuous variable obtained from the LSE method (abundance in the number of non-native species). This model may be expressed as follows: \(Y = \frac{1}{{1 + e^{ - (\alpha + \beta X)} }}\), where Y is the probability of non-native fish presence, α is analogous to the intercept on linear regression, and β indicates the coefficient of explanatory variable X. The 5% significance level was adopted in these analyses.
Results
RAP results
The RAP was conducted in 24 lakes in the CAF area and 32 lakes in the PERD area. According to the RAP, in the CAF area, non-native species were observed in 13 (54.2%) lakes. In the PERD area, just three (9.4%) presented only native fish in their fish assemblage, while 29 had at least one non-native fish species, according to the RAP. Seven non-native species were recorded in sampled lakes in total: the peacock bass; the red piranha; the oscar; the singing catfish Hoplosternum littorale (Hancock, 1828); the tambaqui Colossoma macropomum (Curvier, 1818); the African catfish Clarias gariepinus (Burchell, 1822); and the Nile tilapia Oreochromis niloticus niloticus (Linnaeus, 1758).
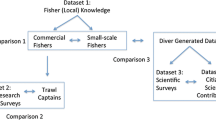
The multiple sampling techniques detected different non-native fish in the two lake areas (Fig. 2) despite their being spatially adjacent and located within the same sub-basin. In the CAF area, where fishing is permitted, the efficiency of sampling techniques differed markedly, and sport fishers’ interviews were particularly effective (Table 1).

Number of lakes with non-native fish recorded by RAP using multiple techniques in the lakes of PERD (Parque Estadual do Rio Doce) and CAF (Companhia Agrícola Florestal) areas at River Doce Basin, Brazil
In 11 of the CAF lakes, the RAP recorded no non-native species, while in the other 13 at least one species was recorded (Table 1). Sport fishers’ interviews recorded non-native fish species in eight out of the 13 lakes where these species occur (based on the total result and the LSE method). In the eight lakes where interviews detected non-native species, only the interview technique rendered positive results, while the other three techniques were inefficient. For the other five lakes where non-native species were recorded, the visual technique was conclusive in two lakes, while gillnetting detected non-native species in the remaining three lakes (Table 2). Fishing with lures did not detect any non-native species in RAP. In the CAF lakes, detection of non-native fish in each lake was obtained by one technique and no detection by other techniques.
In the PERD lakes where fishing is not allowed, just three lakes from the RAP analysis did not present non-native fish (Table 1). In these lakes, the techniques showed more similar efficiency; in 38% of lakes with at least one non-native species, at least two techniques provided the detection. Of the 29 lakes with non-native fish, gillnetting detected their presence in 22. Fishing with lures detected non-native species in 14 out of the 29 lakes, along with visual detection produced results in nine. The gillnetting technique had exclusive records in 11 lakes, while fishing with lures had in five lakes, and visual detection had in two lakes. For the PERD lakes, several techniques recorded the same non-native species (Table 2).
RAP techniques efficiency for different non-native species
In lakes of the CAF area, the recorded non-native species were red piranha (six lakes), peacock bass (seven lakes), Nile tilapia (three lakes), oscar (two lakes), and tambaqui (one lake). The more dispersed species were peacock bass, and red piranha with an occurrence detected of only one specimen in 58.9 and 53.6% of the sampled lakes, respectively. African catfish and singing catfish occurred in 14.3% of the lakes. Oscar occurred in 10.7%, Nile tilapia in 5.3%, and tambaqui in 1.7% of the sampled lakes. Together, all non-native species were found in 76.8% of the sampled lakes.
The interview technique, used only in the CAF area, recorded 100% of red piranha, African catfish, oscar, and tambaqui occurrences, while it recorded 86% for peacock bass, 50% for singing catfish, and 33% for Nile tilapia. At the PERD area, peacock bass occurred in 26 lakes, and red piranha in 24, with both species co-occurring in 21 lakes. The other three non-native species recorded were less frequently found; the oscar was present in four lakes, and the singing catfish and the African catfish in two lakes. The red piranha was detected in 19 out of 24 lakes by gillnetting. In two of these 19 lakes, the detection of the red piranha was made by lures, while in another one this species was visually detected. The peacock bass was detected by gillnetting in 13 out of the 26 lakes, being that ten detections were made exclusively by gillnets. The visual technique was the only one to detect the peacock bass in six different lakes, while fishing with lures only could detect it in five lakes. All occurrences of the oscar, the African catfish, and the singing catfish were obtained with gillnetting.
RAP efficiency
An initial inspection of the CAF data indicates great similarity between non-native species detection by RAP and by LSE. Considering all non-native species, in only one case was there an omission of a species by a method in one of the invaded lakes. The RAP did not fail to detect the red piranha, the peacock bass, and the Nile tilapia, even when they occurred in low abundance (Fig. 3), but did miss the singing catfish in three lakes and the oscar in one lake. On the other hand, the RAP detected the African catfish in one lake, whereas LSE did not.

Abundance (number) of non-native fish sampled in lakes from PERD and CAF areas at River Doce Basin, Brazil. Different symbols represent median values for different species. Upper and lower bars represent 75 and 25% abundance values, while upper and lower sticks represent maximum and minimum abundance values for each species
An initial inspection of the PERD lakes indicates identical detection results to those obtained by Latini & Petrere (2004) for all non-native species, and this was the case for the basic detection of each non-native species alone and the detection of the number of non-native species. The RAP efficiency in the detection of the red piranha, the peacock bass, and the African catfish is very high, permitting detection in situations of low abundance (Table 3; Fig. 4A, B, C). However, the RAP did not perform as efficiently for the oscar and the singing catfish. Although RAP could detect the oscar in low abundance, the number of lakes containing this species is small (Table 3; Fig. 4D). The RAP detected the singing catfish in low abundance but failed to detect its occurrence in some lakes where this species is found in high abundance (Table 3; Fig. 4E).

Results of logistic regression for RAP efficiency analysis in non-native detection at River Doce Basin, Brazil. The X values represent the abundance of non-native species obtained from LSE method and Y values represent detection of non-native fish by RAP method. Bars represent standard deviation measure of variation. Different figures refer to red piranha (A), peacock bass (B), African catfish (C), oscar (D), and singing catfish detection (E). Only A, B, and C represent significant results
Discussion
The RAP is a reliable method to detect non-native fish species in the studied lakes as it presented the same accuracy as a larger sampling effort. Seven non-native species were detected, with the red piranha and the peacock bass as the most frequent. The efficiency of the RAP comes from the multiple techniques employed, as some techniques are more efficient for different species. At the PERD lakes, where information from sport fishers was not available, the other sampling techniques were more effective than they were in the CAF lakes. Gill netting was the most efficient method in the PERD lakes, but the efficiency of this technique was lower in the CAF lakes. Interestingly the abundance of non-native fish species was lower in the CAF lakes (Fig. 3), resulting in lower probability of capture by this technique than in the PERD lakes. The lower abundances of non-native species in the CAF lakes appear to be the result of two factors: more recent introductions and presence of fishing activities that suppress growth rates in the non-native populations.
At the CAF lakes, sport fisher interviews were the most efficient technique for detecting non-native fishes. Without this technique, the RAP would not have detected the occurrence of non-native fish in eight out of the 13 lakes containing them. This efficiency in non-native species’ catches is likely due to fishers being talented in the use of fishing methods and spending a lot of time fishing, along with several non-native species being of commercial interest.
The superior efficiency of gillnetting in the PERD lakes and of the sport fishers’ interviews in the CAF lakes does not diminish the importance of other techniques. Visual detection and lures were unique in the identification of non-native species in some lakes. For example, peacock bass was exclusively identified by visual techniques in the Baixa Verde lake in the CAF area and by lures in the Juquita lake in the PERD area, corroborating the importance of multiple techniques in the RAP methodology.
Sport fishers are cited as possible dispersion agents of non-native fish in the River Doce lakes (Lima et al., 2010; Latini et al., 2016). The results obtained from interviews indicate that they are also one of the important tools for analyzing the problem. This indicates that the RAP methods for fish surveys have to make use of fishers’ interviews as part of the sampling technique. Fish stock studies use information from fishers (e.g., Petrere et al., 2004; Golden et al., 2014), and biological invasion scientists may consider doing the same.
Larger sampling effort methods allowed to identify non-native species of lower abundance in the CAF and the PERD lakes. However, the RAP also detected non-native species in low-abundance situations. The detection of oscar and singing catfish was less efficient by RAP, suggesting it was dependent on species abundance. Besides that, general RAP’s success suggests that it has potential for use in monitoring non-native fish species invasions in large areas, such as the River Doce lakes area, what may produce the best results to minimize the economic costs associated with non-native species invasions (Heger & Trepl, 2003; Zanden et al., 2011).
Particularly in Eastern Brazil tropical lakes, the RAP is as efficient as the LSE method with the advantage of having lower costs. However, the possibility of comparison between the two methods as done in this study is not common in other areas. Here, specific efforts of two methods were calculated, and high quality in RAP methodology was observed. Moreover, if a problem requires RAP in a specific site, it is likely that, with the agility required by the problem, the researcher will not have time to verify what is the best methodology. An additional problem is that some of the techniques used here will not be possible on other sites: for example, if the study is done in a protected area, there should be no fishers; if the water is turbid, visual observation might not be successful, or if the ponds are too shallow, gillnetting may be impractical. However, other new tools can be useful to specific situations: electrofishing, benthic gillnets, multi-mesh gillnets, or different times for sampling or sampling periods can increase the quality of a RAP in other situations. In other words, the significant finding of this work is that with knowledge about regional characteristics, it is possible to make a RAP with a set of valuable sampling methods and thus obtain optimum solutions for local questions worldwide.
There are studies showing the results of fishing at different times of day (e.g., Kocovsky et al., 2010), comparing the efficiency of a methodology in different habitat types (e.g., Deceliere-Verges et al., 2009) and more frequently comparing methods in the same site (e.g., Olin et al., 2009). However, apparently, there are no studies comparing the efficiency of multiple sampling approaches, and this study could help to supply information about the effort required for distinct sample situations, such as habitat heterogeneity, spatial patterning, or population abundance, which can improve efficiency using RAPs to the non-native fish survey.
References
Britski, H. A., K. Z. S. Silimon & B. S. Lopes, 1999. Peixes do Pantanal – Manual de identificação. Embrapa CPAP, Corumbá.
Deceliere-Verges, C., C. Argillier, C. Lanoiselee, J. De Bortoli & J. Guillard, 2009. Stability and precision of the fish metrics obtained using CEN multi-mesh gillnets in natural and artificial lakes in France. Fisheries Research 99: 17–25.
Douglass, L. L., H. P. Possingham, J. Carwardine, C. J. Klein, S. H. Roxburgh, J. Russell-Smith & K. A. Wilson, 2011. The effect of carbon credits on savanna land management and priorities for biodiversity conservation. PLoS ONE 6: 1–11. https://doi.org/10.1371/journal.pone.0023843.
Géry, J., 1977. Characoids of the World. T F H Publication Inc., Ltd., Neptune City.
Giacomini, H. C., D. P. Lima-Jr, A. O. Latini & H. M. V. Espírito-Santo, 2011. Spatio-temporal segregation and size distribution of fish assemblages as related to non-native species occurrence in the middle rio Doce Valley, MG, Brazil. Neotropical Ichthyology 9: 135–146.
Golden, A. S., W. Naisilsisili, I. Ligairi & J. A. Drew, 2014. Combining natural history collections with fisher knowledge for community-based conservation in Fiji. PLoS ONE 9: e98036.
Harrison, J. A. & P. Martinez, 1995. Measurement and mapping of avian diversity in southern Africa: implications for conservation planning. The International Journal of Avian Science 137: 410–417.
Harwood, J. D. & M. N. Parjulee, 2010. Global impact of biological invasions: transformation in pest management approaches. Biological Invasions 12: 2855–2856.
Heger, T. & L. Trepl, 2003. Predicting biological invasions. Biological Invasions 5: 313–321.
Hosmer, D. W. & S. Lemeshow, 1989. Applied logistic regression. Wiley, New York.
Kocovsky, P. M., M. A. Stapanjan & C. T. Knight, 2010. Night sampling improves indices used for management of yellow perch in Lake Erie. Fisheries Management and Ecology 17: 10–18.
Krebs, C. J., 1999. Ecological Methodology. Benjamin/Cummings, Menlo Park.
Latini, A. O. & M. Petrere Jr., 2004. Reduction of a native fish fauna by alien species: an example from Brazilian freshwater tropical lakes. Fisheries Management and Ecology 11: 71–79.
Latini, A. O. & M. Petrere Jr., 2007. Which factors determine non-indigenous fish dispersal? A study of the red piranha in tropical Brazilian lakes. In Gherardi, F. (ed.), Biological invaders in inland waters: profiles, distribution and threats. Springer, Dordrecht: 415–422.
Latini, A. O., L. T. Oporto, D. P. Lima-Júnior, D. C. Resende & R. O. Latini, 2016. Peixes. In Latini, A. O., D. C. Resende, V. B. Pombo & L. Coradin (eds), Espécies exóticas invasoras de águas Continentais no Brasil. MMA, Brasília: 295–581.
Lima, F. P., A. O. Latini & P. De Marco Jr, 2010. How are the lakes? Environmental perception by fishermen and alien fish dispersal in Brazilian tropical lakes. Interciencia 35: 84–91.
Lodh, R. & B. K. Agarwala, 2016. Rapid assessment of diversity and conservation of butterflies in Rowa Wildlife Sanctuary: an Indo-Burmese hotspot – Tripura, N.E., India. Tropical Ecology 57: 231–242.
MacIsaac, H. J., B. Beric, S. A. Bailey, N. E. Mandrak & A. Ricciardi, 2015. Are the Great Lakes at risk of new fish invasions from trans-Atlantic shipping? Journal of Great Lakes Research 41: 1172–1175.
Madalozzo, B., T. G. Santos, M. B. Santos, C. Both & S. Cechin, 2017. Biodiversity assessment: selecting sampling techniques to access anuran diversity in grassland ecosystems. Wildlife Research 44: 78–91.
Marr, S. M., B. R. Ellender, D. J. Woodford, M. E. Alexander, R. J. Wasserman, P. Z. Ivey, T. Zengeya & O. L. F. Weyl, 2017. Evaluating invasion risk for freshwater fishes in South Africa. Bothalia – African Biodiversity & Conservation 47: 1–10.
Metzeling, L., B. Chessman, R. Hardwick & V. Wong, 2003. Rapid assessment of rivers using macroinvertebrates: the role of experience, and comparisons with quantitative methods. Hydrobiologia 510: 39–52.
Nelson, J. S., 1994. Fishes of the World. Wiley, New York.
Nimer, E., 1989. Climatologia do Brasil. IBGE, Rio de Janeiro.
Nunes, A. L., E. Tricarico, V. E. Panov, A. Cardoso & S. Katsanevakis, 2015. Pathways and gateways of freshwater invasions in Europe. Aquatic Invasions 4: 359–370.
Olin, M., I. Malinen & J. Ruuhijarvi, 2009. Gillnet catch in estimating the density and structure of fish community-Comparison of gillnet and trawl samples in a eutrophic lake. Fisheries Research 96: 88–94.
Petrere Jr., M., R. B. Barthem, E. A. Córdoba & B. C. Gómez, 2004. Review of the large catfish fisheries in the upper Amazon and the stock depletion of piraíba (Brachyplatystoma filamentosum Lichtenstein). Reviews in Fish Biology and Fisheries 14: 403–414.
Sant, N., E. Chappuis, C. Rodríguez-Prieto, R. Montserrat & E. Ballesteros, 2017. Cost-benefit of three different methods for studying Mediterranean rocky benthic assemblages. Scientia Marina 81: 129–138.
Simberloff, D., M. Jean-Louis, P. Genovesi, V. Maris, D. A. Wardle, J. Aronson, F. Courchamp, B. Galil, E. García-Berthou, et al., 2013. Impacts of biological invasions: what’s what and the way forward. Trends in Ecology and Evolution 28: 58–66.
Sutherland, W. J., 2001. The Conservation Handbook, Research, Management and Policy. Blackwell Science, Cornwall.
Wu, R., Y. Long, G. P. Malanson, P. A. Garber, S. Zhang, D. Li, et al., 2014. Optimized spatial priorities for biodiversity conservation in China: a systematic conservation planning perspective. PLoS ONE 9: e103783.
Zanden, M. J. V., G. J. A. Hansen, S. N. Higgins & M. S. Kornis, 2011. Invasive species early detection and eradication: a response to Horns (2011). Journal of Great Lakes Research 37: 595–596.
Acknowledgements
This study was supported by the Fundo Nacional do Meio Ambiente (FNMA—Grant 80/2001), Ministry of Environment, and by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). The Instituto Estadual de Florestas de Minas Gerais (IEF-MG) and the Companhia Agrícola Florestal Santa Bárbara (CAF—Arcelor) provided logistic support. Thanks to Adelaine La Guardia for improvement in the English quality of this final version. The authors are grateful to the two anonymous reviewers who improved the clarity and quality of the paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: John E. Havel, Sidinei M. Thomaz, Lee B. Kats, Katya E. Kovalenko & Luciano N. Santos / Aquatic Invasive Species II
Rights and permissions
About this article

Cite this article
Latini, A.O., Petrere Júnior, M. Efficiency of rapid field methods for detecting non-native fish in Eastern Brazilian lakes. Hydrobiologia 817, 85–96 (2018). https://doi.org/10.1007/s10750-018-3624-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-018-3624-x