
Abstract
The positive relationship between species regional distribution and local abundance is one of the most ubiquitous patterns in ecology. Among the hypotheses proposed to explain the relationship, the niche breadth and the niche position (or habitat availability) hypothesis are the most investigated. An unappreciated issue, but that is likely to be important for the understanding the relationship is the nature of variables used to estimate niche measures. Here, we analyzed the form of this relationship in lotic chironomid genera and tested whether niche measures estimated from local and landscape variables explain the observed pattern. Analyses were based in 47 forested streams within Southeastern Brazil. From our data set, we randomly partitioned the data in two non-overlapping sets to estimate taxa distribution and abundance (Distribution Data; n = 23 sites) and to generate niche measures (Niche Data; n = 24). We repeated that process 1,000 times, and for each one, we generated niche breadth and position measures using in-stream and landscape variables and estimated abundance and distribution for each taxa. With these, we estimated the relationship between both abundance and distribution and niche measures using ordinary least-squares regressions. We found no relationship between niche position estimated from local variables and local abundance nor regional distribution. There was a negative relationship between niche position estimated from landscape and local abundance, and regional distribution. We found a positive relationship between niche breadth (local and landscape) and both local abundance and regional distribution. When the relationship was significant, both niche position and niche breadth explained less than a half of total variation in abundance and distribution. This suggests that not only niche-based processes, but also other mechanisms may be responsible for the abundance–distribution relationship in lotic chironomids. A novel finding of this study was that although there was much unexplained variability around the relationships, niche breadth was a better predictor of abundance and distribution than niche position. We suggest that future studies should investigate if spatial processes, like dispersal, together with environmental processes affect interspecific abundance–distribution relationships.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
One of the most extensively investigated large-scale patterns in ecology is the relationship between species local abundance and their regional distribution (Williams, 1960; Brown, 1984; Gaston & Blackburn, 2000). In most cases, the relationship is positive, i.e., locally abundant species tend to be widespread (Gaston et al., 1997). This pattern is stronger for marine than for terrestrial systems, whereas the weakest relationships are found in freshwater systems (Blackburn et al., 2006). However, it tends to hold despite variations in the way data are gathered and analyzed (Gaston, 1994) and the taxonomic group considered (e.g., Gaston et al., 1998; Heino, 2005; Harcourt et al., 2005; Soininen & Heino, 2005; Heino & Virtanen, 2006).
Various hypotheses have been proposed to explain this relationship. These are mainly related to niche-based mechanisms, metapopulation dynamics, range position, and sampling artifacts related to phylogenetic and spatial non-independence (see Gaston et al., 1997). The niche breadth hypothesis (Brown, 1984) and the niche position (or habitat availability) hypothesis (Hanski et al., 1993; Venier & Fahrig, 1996) are among the most investigated with the latter, receiving good support (Gaston & Blackburn, 2000). In short, the former predicts that species occupying broader range of habitats and exploiting diverse environmental conditions and resources (i.e., wide niche breath) would be able to occupy more places. The latter hypothesis predicts that species utilizing common and widespread resources (or habitats) in a given region would be widespread and abundant. Some studies have demonstrated positive correlations between niche breadth and distribution, although few have demonstrated positive correlation between niche breadth and abundance (Gaston et al., 1997).
The niche of a species can be defined as the environmental conditions that allow this species to satisfy its minimum requirements, so that birth rate of a local population is equal to or greater than its death rate (Hutchinson, 1957; Chase & Leibold, 2003). For freshwaters, environmental characteristics of the water body (e.g., water flow and chemistry) are believed to be closely linked to the characteristics of the surrounding landscape (Frissell et al., 1986; Wiens, 2002). According to Poff’s (1997) landscape filter concept, environmental factors act to determine the occurrence and abundance of species at different spatial scales. That is, to be part of a local community, species in the regional pool must have appropriate characteristics to “pass” through the nested filters (Poff, 1997). Therefore, one could hypothesize that niche properties measured at the local and landscape levels would provide similar explanations for the abundance–distribution relationship.
Most studies on the relationship between distribution and abundance were developed in temperate regions and focused on terrestrial ecosystems (see Blackburn et al., 2006). An investigation on tropical freshwater organisms would certainly contribute to reduce system (and, indirectly, taxonomic) biases and to unravel the role of the mechanisms proposed to account for the relationship. Aquatic chironomids (Chironomidae: Diptera) are a useful group for exploring the abundance–distribution relationship due to their ecological importance as well as to applied aspects. Besides representing one of the most species-rich and abundant group in most aquatic environment, chironomids also present a range of life history that differs markedly, for example, in lifespan, locomotion, feeding habits, and physiological tolerance to oxygen deficit (Pinder, 1986).
It has been demonstrated that some assemblage patterns hold for different taxonomic resolutions on stream macroinvertebrates (e.g., species, genus, and family levels: Marchant et al., 1995; Lenat & Resh, 2001; Melo, 2005). Similar to species distributions, the distribution of higher taxa seems to be related to environmental and spatial variables as well (Murphy & Davy-Bowker, 2005). The reliability of the higher-taxa approach to detect general ecological patterns depends on how species within higher taxa respond to environmental gradients. If their responses are correlated, ecological patterns (e.g., abundance–distribution relationship) can be detected independently of the taxonomic resolution. Having this in mind, one could expect that the abundance–distribution relationship would also occur at higher taxonomic levels other than species, which would be desirable bearing in mind the limited knowledge available for Neotropical fauna. This seems especially suitable for chironomids, in which there are a high number of synonyms and unclear descriptions (Spies & Sæther, 2004) that create problems associated with differing species validity (Ferrington, 2008).
In this study, we investigated the relationship between local abundance and regional distribution of lotic chironomids. First, we analyzed the form of the relationship in an attempt to find out if a positive relationship holds at the genus level. Based on a “higher-taxon approach,” we hypothesized that there would be a positive relationship between distribution and local abundance in lotic chironomid genera. If so, we asked if the relationship could be explained by niche characteristics of taxa estimated using local and landscape environmental variables. Following Poff’s (1997) landscape filter concept, we hypothesized that niche breadth (Brown, 1984) and niche position (Hanski et al., 1993) estimated using both local in-stream and landscape variables would explain the positive relationship. We expected that both local abundance and regional distribution of chironomid genera would be positively related to their niche breadth and negatively related to their niche position. Moreover, these relationships should be significant when defining the niche with local and landscape features. However, given that the finest grained environmental filter through which a species go by is the local one (Poff, 1997), we predicted that the amount of variation explained in the relationships should be higher when using niche breadth and position estimated from the local scale.
Materials and methods
Studied sites and measured variables
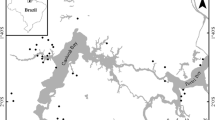
We used data on chironomid larvae distribution extracted from the “Macroinvertebrates Database” compiled by the research group of the “Laboratório de Entomologia Aquática—Universidade Federal de São Carlos.” This data were collected during the dry seasons of 2001, 2005, and 2006 using Surber sampler (0.1 m2 area and 250 mm mesh size) in 47 forested streams of southeastern Brazil (20–25°S, 44–53°W; Fig. 1). Our research group has been continually visiting all sampling areas since 2001 and did not notice any drastic change in land use during this period.

The geographical location of the studied streams in the São Paulo state, Brazil. Areas in gray within the limits of São Paulo state represent forested areas
Sites were typical of Brazilian Atlantic Forest (sensu Oliveira-Filho & Fontes, 2000) headwater streams with water depths <50 cm, tree canopy coverage exceeding 70% of the channel, and absence of macrophytes. The riparian vegetation along all streams was well preserved. Six sampling units (three samples from pool and three from riffle sites) of chironomid larvae were taken randomly from a 100-m reach within each stream using a Surber sampler. For each stream, sampling units were pooled prior to statistical analysis. We mounted specimens on slides and identified them to genus level.
Environmental data
Local environmental measurements were taken at each site to characterize habitat conditions. Conductivity, pH, and dissolved oxygen were measured in situ using a Horiba U-10 or a Yellow Springs-556 water checker equipped with multiple probes. Percentage of canopy cover and predominant substrates were estimated visually. Substrates were classified as the proportion of the stream bottom covered by boulder and cobble (>256 mm), gravel (2–255 mm), sand (0.125–2 mm), and mud (<0.125 mm).
In order to compute the landscape metrics, we delineated a circular buffer area (500 m radius, ca. 78.5 ha) around a point located in the center of the stream channel of each of the 47 sampling sites (see Umetsu et al., 2008; for application of landscape metrics based on buffers of varying width). Some of our landscape metrics were derived from a land cover map at a scale of 1:50,000 from the Forestry Institute of São Paulo (Metzger et al., 2008). Macro-regional climatic variables, derived from coarse-scale maps, were also included in this set of variables. Landscape metrics derived from the land cover map were: cover area of forest; the total edge contrast index (TECI), and the edge density (LED). The macro-regional variables were: the enhanced vegetation index (EVI), the rainfall, and the solar radiation, all of which are indicative of primary production and biomass accumulation. We included four variables based on EVI measures: the EVI for the autumn (EVI AR) and for the winter (EVI AM), and both the range (EVI WR) and mean (EVI WM) between the two seasonal variables. We also included a measure of elevation in our set of landscape metrics. This metric was calculated using the mean value of the altitude across a 500-m radius circle around each sampling site provided by the Shuttle Radar Topography Mission (SRTM/NASA; http://www2.jpl.nasa.gov/srtm). Cover area of forest, total edge contrast index, and edge density were calculated using FRAGSTATS 3.3 (McGarigal et al., 2002), whereas the enhanced vegetation index and elevation were calculated using the Zonal statistics tool in ArcGis 9.
Niche measures and data analysis
From our database, we randomly selected 23 sites to estimate the taxa distribution and abundance and 24 to generate niche measures. Hereafter, these data sets are referred as “Distribution Data” and “Niche Data”. The reason for using two independent data sets is to overcome the circularity that occurs when one estimates species’ distribution and niche from the same data. This statistical bias is mainly associated with the problem of dissociating sample size effects from real differences between the niche breadth and position of common and rare species (Gaston, 1994). However, only that does not guarantee that our estimations are truly unbiased. For example, let us suppose that, by chance, the “Niche Data” included the most similar streams regarding environmental conditions. This would enhance the chance of reducing niche measures variability among different taxa and, thus, produce a biased model. For this reason, we randomly generated 1,000 Distribution and Niche data sets, and for each one we applied the steps explained below.
For each genus within the Distribution data sets, we calculated the regional distribution by summing the number of sites occupied by that genus and genus local abundance as the number of larvae of a given genus found in each stream. The ecological niche of a taxon can be represented by using its mean position and breadth along various environmental axes (Schoener, 1989). We applied the Outlying Mean Index (OMI) analysis (Dolédec et al., 2000; Thuiller et al., 2005, Broennimann et al., 2006 for other applications of this method) to generate measures of niche position and niche breadth for each taxon using the local and landscape variables measured within the Niche Data. The OMI (niche position hereafter) is a measure of the distance between the mean habitat conditions used by a taxon (centroid) and the mean habitat condition of the entire sampling area (origin of the niche hyperspace). The niche position of the taxon is assessed through their niche deviation from a reference. This reference represents a theoretical ubiquitous taxon that tolerates the most common habitat conditions. Genera that display high values of niche position have marginal niches (its environmental requirements are far from the mean conditions of the study area). Thus, we tested the niche availability hypothesis (Hanski et al., 1993; Venier & Fahrig, 1996) using OMI niche position. The OMI analysis also provides a measure of tolerance or niche breadth. Genera with high values of niche breadth are generalists, occurring across large portions of environmental gradients (wide habitat niche breadth).
Most former studies on the abundance–distribution relationship regarded abundance as the response variable. However, we cannot be sure whether the predominant direction of causality (if present) runs from local abundance to regional distribution or from the opposite (for a discussion, see Gaston & Blackburn, 2000). Thus, we estimated the relationships between both local abundance and regional distribution (response variables) with the explanatory variables niche breadth and niche position (both defined using local and landscape variables) using ordinary least-squares regression. All these variables were log-transformed before analysis.
We did each regression mentioned above 1,000 times—using the randomly partitioned data sets (Distribution and Niche data sets)—producing a distribution of coefficients of determination (r 2), P values, and regression coefficients, which we used to access the average explanatory power of each model. We only considered those significant models with less than 50 non-significant regressions (α = 5%). All analyses were performed in the R-language environment (R Development Core Team, 2006) using the package “ade4” (Dray & Dufour, 2007) for generating niche measures.
Results
We identified 41 chironomid genera occurring in all sampled streams. The most widely distributed taxa were Endotribelos (41 streams), the Tanytarsus/Caladomyia complex (40), Polypedilum (38), Parametriocnemus (35), Rheotanytarsus (33), and Larsia (31). Likewise, the most abundant taxa were the Tanytarsus/Caladomyia complex, Polypedilum, Parametriocnemus, Rheotanytarsus, Endotribelos, and Larsia. As we expected, there was a positive relationship between genus distribution and local abundance. This relationship was significant (P ≤ 0.05) in all 1,000 randomizations, with the models accounting for most of the variability in regional distribution (mean r 2 = 0.88, Fig. 2).

Distribution of coefficients of determination (r 2) of the regressions between regional distribution and local abundance. The arrow indicates the mean value of r 2 (0.88) across all 1,000 data subsets. All regression models between regional distribution and local abundance were significant at P ≤ 0.05
Niche position, as defined by local environmental variables, was not significantly related neither with local abundance nor with regional distribution in 52.4 and 62.4% of all data sets, respectively (Fig. 3a, c). On the other hand, when estimated from landscape variables, there was a significant negative relationship between niche position and local abundance in 99.8% of all data sets (Fig. 3b) and regional distribution in 99.7% of all data sets (Fig. 3d). We found a significant positive relationship between niche breadth (estimated using local and landscape variables) and both local abundance and regional distribution in more than 95% of all data sets (Fig. 4). In general, niche position and niche breadth explained less than a half of total variations in both local abundance and regional distribution (Figs. 3, 4). Therefore, in summary, niche breadth models significantly explained variation in abundance and distribution at both scales, whereas niche position models were significant only when we used landscape variables. Furthermore, in cases where they were significant, niche position and niche breadth models explained similar amounts of total variation in the response variables (Figs. 3, 4).

Distribution of r 2 values generated based on 1,000 regressions between niche position and local abundance (a, b) and regional distribution (c, d). Arrows indicate the mean value of r 2 across all 1,000 data subsets (a r 2 = 0.13*, b r 2 = 0.39, c r 2 = 0.16*, d r 2 = 0.40). * Non-significant models (a 47.6% of regressions with P > 0.05, c 37.6% of regressions with P > 0.05)

Distribution of r 2 values generated based on 1,000 regressions between niche breadth and local abundance (a, b) and regional distribution (c, d). Arrows indicate the mean value of r 2 across all 1,000 data subsets (a r 2 = 0.40, b r 2 = 0.39, c r 2 = 0.42, d r 2 = 0.36). More than 95% of regressions were significant at P ≤ 0.05
Discussion
Results obtained here confirm our expectation that the positive abundance–distribution relationship already observed in many taxa (see Gaston & Blackburn, 2000) also holds true for lotic chironomid genera. This result adds new evidence to the conclusion that higher taxonomic levels can be used to detect local assemblage patterns (e.g., Marchant et al., 1995; Melo, 2005) as well as macroecological relationships (e.g., Harcourt et al., 2005). Unfortunately, we do not have data on species level, thus it was not possible to analyze how much information was lost, if any, when using genus level identification. Nevertheless, the use of a higher taxonomic level aids in minimizing the potential statistical problem of phylogenetic dependency (which is a type of pseudo-replication) making the analyses more conservative. This is especially advantageous here since phylogenetic non-independence is one of the proposed artifactual mechanisms that can generate a positive abundance–distribution relationship (Gaston & Blackburn, 2000).
In general, we found that most of the variation in abundance and distribution was not explained by niche measures. These results are partly similar to previous studies on freshwater organisms. Tales et al. (2004) found support for the niche position hypothesis (Hanski et al., 1993; Venier & Fahrig, 1996) as a mechanism to explain the abundance–distribution relationship. However, they found that niche breadth was not a good explanation for variation in abundance and distribution of riverine fish. On the other hand, Heino (2005) found significant relationships between both niche position and niche breadth and abundance–distribution in stream insects. A general characteristic of these studies and ours is that there is much unexplained variability around these relationships. Our results differ from those reported by Heino (2005) in the sense that in his study niche position explained more variation in both abundance and distribution than niche breadth. Here, niche position estimated using local environmental variables did not explain variation in either local abundance or regional distribution. Therefore, the observed positive relationship between abundance and distribution is not a consequence of taxa niche position, regarding local environmental variables. This result is surprising given that most studies on the abundance–distribution relationship have found stronger support for the niche position hypothesis (see review in Gaston et al., 1997). This lack of support for the niche breadth hypothesis reported by several previous studies is due to, in part, difficulties in generating adequate niche breadth measurements (Gaston, 1994). Given the multidimensional nature of the niche (Hutchinson, 1957), important variables describing niche breadth might be missing from analyses. Also, an artifactual relationship is expected to arise because of the estimation of niche position is always unbalanced (Gaston et al., 1997). That is, rare and less widespread species contribute with a great number of zeros to the species matrix; thus, it is expected that those species will attain high values of niche position, i.e., marginal niches. The use of an independent data to estimate niche breadth through a resampling procedure like ours does not completely solve this particular problem but minimize it.
Here, we adopted a simple analytical procedure never used before—as far as we know—to estimate the average expected relationships between organism’s abundance and distribution, and their niche. A resampling procedure like ours is a useful tool to minimize the problem of dependence explained above and also to avoid the biased choice of two independent data sets. It is important to notice that we found considerable variability in the coefficients of determination (r 2). For example, we showed that it is possible to find, depending on the particular data subset, models with low explanatory power but also models explaining almost 80% of the variance in the data (Fig. 4d). Also, we showed that sometimes almost a half of the possible models can be non-significant (Fig. 3a). In that sense, we think that future studies seeking for such relationships would benefit from an approach similar to the one we used here because it is a step toward leaving particular explanations (related, for example, to a single data set) to a more general view of the processes that might being operating in nature.
Besides explaining only less than a half of the total variation in local abundance and regional distribution, these relationships seem to be affected by the type of variable used to generate niche measures. Local environmental features of a stream can be partially determined by factors acting at the scale of the surrounding landscape (Hynes, 1970). The OMI analyses we used here provided a description of the variability of habitats used by each taxa in the in-stream and surrounding environmental space. Thus, considering the expected association between in-stream and catchment environmental factors, one would expect that niche measures provided similar responses at both local and landscape levels. This was not the case here. For example, we found that niche position did not explain variation in any response variables when it was estimated from local environmental factors, but was related to both abundance and distribution when estimated from landscape variables. Note, however, that this is not a matter of landscape variables being necessarily more suitable to estimate niche measures. Niche breadth estimated from both local and landscape variables explained similar amounts of variation in local abundance and regional distribution. A number of studies have focused on how local and landscape characteristics of a stream can act to influence the distribution and abundance of macroinvertebrates (Corkum, 1992). Some of these studies have suggested that both scales act in structuring local communities, whereas others have found that local stream variables play a major role (see Richards et al., 1997; Death & Joy, 2004). Here, we demonstrated that landscape-based models were significant independently of the response and predictor variables, whereas local-based models were significant only when niche breadth was the predictor variable. Thus, at this moment, we can only agree with the view that stream communities are structured by processes operating at different spatial scales (Vinson & Hawkins, 1998), without ruling out which scales and factors are the most influential.
Given the high amount of unexplained variation in our regression models and assuming that we have measured appropriate environmental variables to represent niche, we postulate that niche-based processes may not be the main causes for the abundance–distribution relationship in lotic chironomids. In addition to niche position and breadth, some other mechanisms have been proposed as possible determinants of the positive relationship between abundance and distribution of species (Gaston et al., 1997). These other mechanisms are generally classified as statistical, range position, and population dynamic explanations. Since they are not mutually exclusive, it is likely that the abundance–distribution relationship could be a result of multiple processes (Gaston et al., 2000). Statistical explanations are mainly related to sampling artifacts, e.g., non-detection of uncommon taxa that are actually present at a given site, and phylogenetic non-independence, i.e., related taxa does not constitute independent data points in analysis. Previous studies have refuted these two statistical explanations by using large data sets of well-known groups and by controlling for phylogenetic non-independence (see Murray et al., 1998). Although we have not controlled for phylogeny here (there is no available phylogeny for Chironomidae), the most abundant and distributed genera belong to different tribes and subfamilies, so we believe this was not a major cause for the observed relationship. Range position explanations state that because, in general, species abundance are higher at the centers of its geographical range, species whose range limits are located within the study region would have lower abundances (Gaston & Blackburn, 2000). This mechanism was also rejected by a number of studies (see Gaston et al., 1997). As our study was based on genus, the likelihood of this mechanism is diminished here, because the sampled genera have geographical ranges that extrapolate our study area. Finally, population dynamic explanations stem from metapopulation models, mainly the rescue effect hypothesis (Hanski, 1991). In this, dispersal between patches decreases the probability of local population extinction and increases the proportion of patches occupied by a given species. Experimental evaluations of this hypothesis have been conducted but lead to equivocal conclusions. Gonzalez et al. (1998) experimentally interrupted dispersal between moss fragments resulting in a decline in both abundance and distribution, and the loss of the positive abundance–distribution relationship. When dispersal was re-established, there was an increase in both abundance and distribution and the positive relationship between them appeared again. On the other hand, Warren & Gaston (1997) found positive relationships in all treatments of an experiment with protists, even in those where patches were isolated by dispersal limitation. Unfortunately, we do not have information on metapopulation dynamics for the analyzed data. Therefore, a direct investigation of this mechanism is not possible at present.
Recent advances in community ecology point that niche assembly and dispersal assembly models are not mutually exclusive in explaining the same community patterns (Chave et al., 2002, Mouquet & Loreau, 2002). For instance, Cottenie (2005) suggested that when both environment and spatial processes are acting, local communities are structured by a combination of species sorting and mass effects dynamics. Species sorting dynamics emphasizes the importance of species niche and environmental heterogeneity in determining community structure and also assumes moderate dispersal rates (Chave et al., 2002). A mass effect describes a sink-source process of dispersal in heterogeneous environments high enough to change population abundances (Holt, 1993). There is a parallel here between these and the proposed mechanisms for the abundance–distribution relationship (niche-based hypothesis and rescue effect hypothesis). Thus, we believe that future studies should investigate if spatial processes, like dispersal, together with environmental processes affect the abundance–distribution relationship in the context of community structure. This perspective has recently been combined in a unified framework for explaining how compositions of local communities vary in space. The metacommunity framework (Leibold et al., 2004) offers the theoretical background and the analytical tools to integrate local and regional processes in a more inclusive concept for understanding community dynamics.
In summary, the main picture emerging here is that niche-based processes are not the unique cause for variation in the abundance and distribution of lotic chironomids. A novel finding of this study was that we found more support for the niche breadth hypothesis (Brown, 1984) than for the niche position hypothesis (Hanski et al., 1993). We also demonstrated the pertinence of using higher taxa data to analyze the relationship between distribution and abundance. Finally, we showed how a simple resampling procedure can be a useful tool to minimize the lack of independence in estimating niche, abundance, and distribution of taxa from the same data set.
References
Blackburn, T. M., P. Cassey & K. J. Gaston, 2006. Variations on a theme: sources of heterogeneity in the form of the interspecific relationship between abundance and distribution. Journal of Animal Ecology 75: 1426–1439.
Broennimann, O., W. Thuiller, G. Hughes, G. F. Midgley, J. M. R. Alkemade & A. Guisan, 2006. Do geographic distribution, niche property and life form explain plants’ vulnerability to global change? Global Change Biology 12: 1079–1093.
Brown, J. H., 1984. On the relationship between abundance and distribution of species. American Naturalist 124: 255–279.
Chase, J. M. & M. A. Leibold, 2003. Ecological Niches – Linking Classical and Contemporary Approaches. The University of Chicago Press, Chicago, IL.
Chave, J., H. C. Muller-Landau & S. A. Levin, 2002. Comparing classical community models: theoretical consequences for patterns of diversity. The American Naturalist 159: 1–23.
Corkum, L. D., 1992. Spatial distribution patterns of macroinvertebrates along rivers within and among biomes. Hydrobiologia 239: 101–114.
Cottenie, K., 2005. Integrating environmental and spatial processes in ecological community dynamics. Ecology Letters 8: 1175–1182.
Death, R. G. & M. K. Joy, 2004. Invertebrate community structure in streams of the Manawatu-Wanganui region, New Zealand: the roles of catchment versus reach scale influences. Freshwater Biology 49: 982–997.
Dolédec, S., D. Chessel & C. Gimaret-Carpentier, 2000. Niche separation in community analysis: a new method. Ecology 81: 2914–2927.
Dray, S. & A. B. Dufour, 2007. The ade4 package: implementing the duality diagram for ecologists. Journal of Statistical Software 22: 1–20.
Ferrington Jr., L. C., 2008. Global diversity of non-biting chironomids (Chironomidae; Insecta-Diptera) in freshwater. Hydrobiologia 595: 447–455.
Frissell, C. A., W. J. Wiss, C. E. Warren & M. D. Huxley, 1986. A hierarchical framework for stream classification: viewing streams in watershed context. Environmental Management 10: 199–214.
Gaston, K. J., 1994. Rarity. Chapman & Hall, London.
Gaston, K. J. & T. M. Blackburn, 2000. Pattern and Process in Macroecology. Blackwell, London.
Gaston, K. J., T. M. Blackburn & J. H. Lawton, 1997. Interspecific abundance-range size relationships: an appraisal of mechanisms. Journal of Animal Ecology 66: 579–601.
Gaston, K. J., T. M. Blackburn, R. D. Gregory & J. J. D. Greenwood, 1998. The anatomy of the interspecific abundance-range size relationship for the British avifauna: I. Spatial patterns. Ecology Letters 1: 38–46.
Gaston, K. J., T. M. Blackburn, J. J. D. Greenwood, R. D. Gregory, R. M. Quinn & J. H. Lawton, 2000. Abundance–occupancy relationships. Journal of Applied Ecology 37: 39–59.
Gonzalez, A., J. H. Lawton, F. S. Gilbert, T. M. Blackburn & I. Evans-Freke, 1998. Metapopulation dynamics maintain the positive species abundance–distribution relationship. Science 281: 2045–2047.
Hanski, I., 1991. Single-species metapopulation dynamics: concepts, models and observations. Biological Journal of the Linnean Society 42: 17–38.
Hanski, I., J. Kouki & A. Halkka, 1993. Three explanations of the positive relationship between distribution and abundance of species. In Ricklefs, R. E. & D. Schluter (eds), Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press, Chicago: 108–116.
Harcourt, A. H., S. A. Coppeto & S. A. Park, 2005. The distribution–abundance (density) relationship: its form and causes in a tropical mammal order, primates. Journal of Biogeography 32: 565–579.
Heino, J., 2005. Positive relationship between regional distribution and local abundance in stream insects: a consequence of niche breadth or niche position? Ecography 28: 345–354.
Heino, J. & R. Virtanen, 2006. Relationships between distribution and abundance vary with spatial scale and ecological group in stream bryophytes. Freshwater Biology 51: 1879–1889.
Holt, R. D., 1993. Ecology at the mesoscale: the influence of regional processes on local communities. In Ricklefs, R. E. & D. Schluter (eds), Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press, Chicago: 77–88.
Hutchinson, G. E., 1957. Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology 22: 145–159.
Hynes, H. B. N., 1970. The Ecology of Running Waters. University of Toronto Press, Toronto.
Leibold, M. A., M. Holyoak, N. Mouquet, P. Amarasekare, J. M. Chase, M. F. Hoopes, R. D. Holt, J. B. Shurin, R. Law & D. Tilman, 2004. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters 7: 601–613.
Lenat, D. R. & V. H. Resh, 2001. Taxonomy and stream ecology: the benefits of genus and species identifications. Journal of the North American Benthological Society 20: 287–298.
Marchant, R., L. A. Barmuta & B. C. Chessman, 1995. Influence of sample quantification and taxonomic resolution on the ordination of macroinvertebrate communities from running waters in Victoria, Australia. Marine and Freshwater Research 46: 501–506.
McGarigal, K., S. A. Cushman, M. C. Neel & E. Ene, 2002. Fragstats: Spatial Pattern Analysis Program for Categorical Maps. Computer Software Program produced by the authors at the University of Massachusetts, Amherst [available at the following web site: www.umass.edu/landeco/research/fragstats/fragstats.html].
Melo, A., 2005. Effects of taxonomic and numeric resolution on the ability to detect ecological patterns at a local scale using stream macroinvertebrates. Archiv fur Hydrobiologie 164: 309–323.
Metzger, J. P., M. C. Ribeiro, G. Ciocheti & L. R. Tambosi, 2008. Uso de índices de paisagem para a definição de ações de conservação e restauração da biodiversidade do Estado de São Paulo. In Rodrigues, R. R., C. A. Joly, M. C. W. Brito, A. Paese, J. P. Metzger, L. Casatti, M. A. Nalon, N. Menezes, N. M. Ivanauskas, V. Bolzani & V. L. R. Bononi (eds), Diretrizes para Conservação e Restauração da Biodiversidade no Estado de São Paulo. Secretaria do Meio Ambiente & FAPESP, São Paulo: 120–127.
Mouquet, N. & M. Loreau, 2002. Coexistence in metacommunities: the regional similarity hypothesis. The American Naturalist 159: 420–426.
Murphy, J. F. & J. Davy-Bowker, 2005. Spatial structure in lotic macroinvertebrate communities in England and Wales: relationships with physicochemical and anthropogenic stress variables. Hydrobiologia 534: 151–164.
Murray, B. R., C. R. Fonseca & M. Westoby, 1998. The macroecology of Australian frogs. Journal of Animal Ecology 67: 567–579.
Oliveira-Filho, A. T. & M. A. L. Fontes, 2000. Patterns of floristic differentiation among Atlantic forests in south-eastern Brazil, and the influence of climate. Biotropica 32: 793–810.
Pinder, L. C. V., 1986. Biology of freshwater chironomidae. Annual Review of Entomology 31: 1–23.
Poff, N. L., 1997. Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. Journal of the North American Benthological Society 16: 391–409.
R Development Core Team, 2006. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. http://www.R-roject.org.
Richards, C., R. J. Haro, L. B. Johnson & G. E. Host, 1997. Catchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshwater Biology 37: 219–230.
Schoener, T. W., 1989. The ecological niche. In Cherret, J. (ed.), Ecological Concepts: The Contribution of Ecology to an Understanding of the Natural World. Blackwell Scientific, Oxford: 790–813.
Soininen, J. & J. Heino, 2005. Relationships between local population persistence, local abundance and regional occupancy of species: patterns in diatoms of boreal streams. Journal of Biogeography 32: 1971–1978.
Spies, M. & O. A. Sæther, 2004. Notes and recommendations on taxonomy and nomenclature of Chironomidae (Diptera). Zootaxa 752: 1–90.
Tales, E., P. Keith & T. Oberdorff, 2004. Density-range size relationship in French riverine fishes. Oecologia 138: 360–370.
Thuiller, W., S. Lavorel & M. B. Araujo, 2005. Niche properties and geographical extent as predictors of species sensitivity to climate change. Global Ecology and Biogeography 14: 347–357.
Umetsu, F., J. P. Metzger & R. Pardini, 2008. Importance of estimating matrix quality for modeling species distribution in complex tropical landscapes: a test with Atlantic forest small mammals. Ecography 31: 359–370.
Venier, L. A. & L. Fahrig, 1996. Habitat availability causes the species abundance–distribution relationship. Oikos 76: 564–570.
Vinson, M. R. & C. P. Hawkins, 1998. Biodiversity of stream insects: variation at local, basin, and regional spatial scales. Annual Review of Entomology 43: 271–293.
Warren, P. H. & K. J. Gaston, 1997. Interspecific abundance–occupancy relationships: a test of mechanisms using microcosms. Journal of Animal Ecology 66: 730–742.
Wiens, J. A., 2002. Riverine landscapes: taking landscape ecology into the water. Freshwater Biology 47: 501–515.
Williams, C. B., 1960. The range and pattern of insect abundance. American Naturalist 94: 137–151.
Acknowledgments
We thank Joaquín Hortal and Professor José Alexandre Diniz-Filho for their valuable comments during the elaboration of this study. Adriano S. Melo, Karl Cottenie, and an anonymous referee provided important criticism in an early version of this manuscript and we also thank them. Financial support was provided by the Brazilian Council for Scientific and Technological Development (CNPq) to T. Siqueira and L. M. Bini, and by The State of São Paulo Research Foundation (BIOTA/FAPESP) to F. O. Roque and S. Trivinho-Strixino.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: David Dudgeon
Rights and permissions
About this article
Cite this article
Siqueira, T., Bini, L.M., Cianciaruso, M.V. et al. The role of niche measures in explaining the abundance–distribution relationship in tropical lotic chironomids. Hydrobiologia 636, 163–172 (2009). https://doi.org/10.1007/s10750-009-9945-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-009-9945-z