
Abstract
In the last few years, the renewed interest for emmer wheat (Triticum dicoccon Schrank) in Italy has stimulated breeding programs for this crop releasing improved genotypes obtained not only by selection from landraces, but even by crosses with durum wheat (Triticum durum Desf.) varieties. The purpose of this work has been to uncover the genetic make-up of some emmer × durum derivatives, specifically by comparing the differences from their parents. Genetic diversity of advanced breeding lines and varieties derived from a durum × emmer cross has been evaluated on the basis of AFLP and SSR markers in comparison with the corresponding emmer and durum wheat parent for addressing the seminal question of how much ‘wild’ variation remains after selection for agronomic type.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Latin ‘far’ and Mediolatin ‘farrum’ words indicate an Italian ethnobotanical complex that includes the tetraploid hulled wheat Triticum dicoccon Schrank. This crop was widely spread during the past in the Mediterranean basin, but neglected during the last century in favour of naked durum wheats and now rediscovered (Perrino and Hammer 1982). The market interest for this crop, explained by some peculiar agronomical and nutritive characteristics (Castagna et al. 1996; Cubadda and Marconi 1996; D’Antuono and Minelli 1998) besides the interest for traditional food (Porfiri et al. 1998), has resulted in an increase of cultivation in a range of Italian environments. The widening of the cultivation area for this crop is in fact justified by its traits of resistance or tolerance to biotic and abiotic stresses that make it well adapted to low-input agricultural systems (Corazza et al. 1986; Damania et al. 1992; Codianni et al. 1996). However, Italian emmer landraces are still characterized by some negative traits, like low level of productivity, excessive plant height, partially brittle rachis and low quality of meal. In fact, even if quality in wheat is a multidisciplinary aspect, depending on the worker and end-use (Troccoli et al. 2000), emmer landraces show some quality failures at the different levels of farmers, milling, pasta industry and consumers (Galterio et al. 1994). For these reasons, breeding programs carried out by the Istituto Sperimentale per la Cerealicoltura (Foggia, Italy) for the improvement of emmer through crosses with durum wheat (Triticum durum Desf.), have produced ‘Mosè’, ‘Padre Pio’ and ‘Davide’, three new emmer varieties. These varieties and their parents, together with advanced breeding lines obtained from the same cross, have been characterized in this work with SSR and AFLP analysis. Moreover, relationships with a set of emmer landraces, historical and modern durum wheat varieties, hexaploid wheats and wild relatives have also been analysed with AFLP approach. Therefore the purpose of this work has been to uncover the genetic make-up of these pool of varieties, specifically by comparing the differences among emmer × durum wheat derived lines and their parents.
Materials and methods
Plant materials
A total of 45 wheat accessions, including Triticum durum Desf., Triticum dicoccon Schrank, Triticum tauschii (Coss.) Schmalh., Triticum spelta L. (a Swiss landrace, present in the collection of the Istituto Sperimentale per la Cerealicoltura, Fiorenzuola d’Arda, as ‘Rotkorn 32’) and Triticum aestivum L. were used in this study: detailed informations on area of origin and pedigree are reported in Table 1. Six numbered advanced breeding lines (88C, 190C, 193C, 238C, 303C and 390C) and three named varieties (‘Mosè’, ‘Padre Pio’ and ‘Davide’), all derivatives from a cross ‘Simeto’ × ‘Molise’, were used together with parents. The varieties have been bred by using pedigree selection, taking into account hulled spikelet trait, short straw as the parental durum variety and earliness. In particular, multilocation yield trials have been established in F6 and from the agronomic results, nine lines have been chosen and evaluated—as F6—for grain yield, protein composition and rheological properties of dough. On the basis of these results three lines have been included in two-years National List Trial as ‘Mosè’, ‘Padre Pio’ and ‘Davide’ and they are under registration in the Italian Register of Varieties. Multiplication stock is under the control of the Istituto Sperimentale per la Cerealicoltura, Section of Foggia.
Agronomical and quality traits evaluation
Grain yield, plant height, heading date, 1,000-kernel weight, grain protein content and SDS sedimentation values reported in Table 2 are the means of the data obtained from a two-years field trial run at the experimental farm of the Istituto Sperimentale per la Cerealicoltura, Foggia, in Southern Italy. The emmer × durum derived breeding lines and varieties were sown for 2 years at the beginning of November in moist conditions and no irrigation was applied, thereafter adopting low-input practices. Seedling density was 200 seeds m−2 and the previous crop was fallow. About 50 N units per hectare of fertilizers were split in two applications: 1/3 at sowing, as diammonium phosphate (100 kg ha−1) and 2/3 at tillering as ammonium nitrate (120 kg ha ha−1). The soil type was clay loam with a pH of 7.8. The crop cycle was from November to the end of June. The size of plot was 10.2 m2 (eight rows, each 7.0 m long and 0.17 m apart), arranged in a randomized complete block design with three replicates. Plants were harvested after physiological maturity during early July.
Statistical analysis was conducted for each measured agronomic and qualitative parameter by analysis of variance (ANOVA) and the means were separated by Tukey’s multiple range test using Mstat-C version 2.00 software.
Molecular marker analysis
For molecular characterization, each population or line or variety was represented by a bulk of 10 g of seeds, ground in a Cyclotec mill. Genomic DNA was extracted from a subsample of flour from each entry, following Murray and Thompson’s (1988) CTAB procedure.
AFLP reactions were done using the “AFLP plant mapping” kit from Applied Biosystems for fluorescent fragment detection. EcoRI and MseI were used for DNA digestion and selective amplification was done using six different EcoRI–MseI primer combinations. The following combinations were used: E55/M48; E56/M50; E56/M48; E55/M59; E59/M59; E59/M48. AFLP fragments were separated by capillary electrophoresis on ABI PRISM 310 DNA Genetic Analyzer (Applied Biosystems) and GENESCAN 2.1 was used to estimate fragment length: only the fragments between 70 bp and 450 bp were scored and a resolution sensitivity of 1 bp was considered.
For SSR analysis, 53 primer pairs designated as XGWM and found to produce polymorphisms between ‘Simeto’ and ‘Molise’, were selected from a bigger set described by Röder et al. (1998). PCR amplifications were carried out according to Röder et al. (1998).
Data analysis
Both SSR and AFLP fragments were scored as binary characters: either present (1) or absent (0). The cluster analysis was performed with the software package TREECON for the construction and drawing of phylogenetic tree (Van de Peer and De Wachter 1994). GGT Graphical Genotypes software (van Berloo 1999) was used to obtain a graphical representation of SSR data.
Results and discussion
Relationships among emmer × durum wheat derived varieties or lines and their parents
The emergence of organic agriculture, together with the demand for health food products, has stimulated the re-introduction of emmer cultivation, together with the activation, for this cereal, of breeding activities directed to the selection of genotypes adapted to low-input environments (Perrino and Hammer 1982). However, De Vita et al. (2006), observing the variability for agronomic and qualitative traits in 20 emmer accessions consisting of landraces, breeding lines or cultivars selected from landraces, have found that the genotypes selected from landraces showed improvement in grain and protein yield, but not in the quality-related traits. On the contrary, a pedigree selection program after an emmer × durum wheat cross can led to the improvement of some qualitative traits: in fact, new genotypes, obtained after ‘Simeto’ × ‘Molise’ cross, have been selected to be closer to wheat parental not only for agronomical traits such as earliness, reduction of height and yield, but above all for some qualitative aspects, like HMW glutenin subunit composition (Codianni et al. 1998, 2000) and SDS. These results obtained can be observed from data reported in Table 2. Among the three varieties and the six advanced breeding lines obtained from ‘Simeto’ × ‘Molise’ cross, six have yield values equal or major than ‘Simeto’ and all have earliness and plant height significantly different from ‘Molise’ and closer to durum wheat parent. The 1,000-weight kernel character shows large variability, ranging from the minimal values of 35–36 g of ‘Simeto’ and ‘Linea 390C’ to the ‘Davide’ maximum value of 57 g. Only ‘Padre Pio’ and ‘Linea 303C’ have SDS values overcoming ‘Simeto’, whereas protein content ranges from 12% to 16% dm. Therefore, the achievement of advanced breeding lines and varieties like ‘Mosè’, ‘Padre Pio’ and ‘Davide’ improved for several qualitative characteristics, but preserving several morphological, functional and botanical traits of emmer, demonstrates the role of durum wheat as parental in emmer breeding programs.
To address the seminal question of what proportion of each of the two parent’s haplotype remains in each derivative and whether the process of breeding to an agronomic type has led to any enrichment of identifiable segments of the parent’s genomes, genetic diversity of parents and breeding lines, has been evaluated on the basis of unmapped AFLP and mapped SSR markers. After a screening step, 162 Eco-AFLP unmapped markers and 53 SSR mapped markers were found to be polymorphic between ‘Simeto’ and ‘Molise’. These two sets of molecular markers were used to evaluate the genetic contribution of the two parents to the nine genotypes obtained from their cross. AFLPs and SSRs evaluate in different way the genetic contribution of the two parents to progenies. Figure 1 shows graphically the percentages of the two parent haplotypes in the progenies. In particular, according to AFLP (Fig. 1A), ‘Molise’ haplotype is over-represented in ‘Mosè’, ‘Padre Pio’, ‘238C’ and ‘390C’ lines, whereas a more balanced contribution of the two parents is observed for ‘190C’, ‘193C’, ‘303C’ and ‘88C’ lines. Figure 1B shows the genetic contribution of the two parents to all progenies measured with SSR markers. From the data obtained all the six advanced breeding lines are characterised by a contribution of ‘Simeto’ and ‘Molise’ haplotypes very near to 50% for each. Interestingly, the three varieties ‘Mosè’, ‘Padre Pio’ and ‘Davide’, selected to be the best not only on the basis of agronomical traits, but even for additional qualitative traits such as protein composition and rheological properties of dough, show the predominance of ‘Simeto’ haplotype, that contributes for 60–70%. Both AFLP and SSR markers show ‘Davide’ closer to ‘Simeto’ parent in comparison with emmer parent. In Fig. 2A is represented the proportion of the two parental haplotypes, estimated on the basis of mapped SSR, in derivative lines and varieties chromosomes: ‘Simeto’ and ‘Molise’ alleles are equally represented in chromosome arms 1B, 2A, 3A, 4B and 6A. The less-balanced chromosome arms are 2B and 6B. In Fig. 2B are reported the graphical representations of these two chromosome arms with opposite assortment of parental alleles: the durum wheat alleles and the emmer alleles are over-represented, respectively in 2B and 6B. In conclusion, the selective pressure for durum wheat traits like plant height, yield and earliness and for emmer traits like hulled spikelets has produced a set of genotypes in which there is not an over-representation of only one of the two parental haplotypes, but a balanced situation is present in the majority of lines. However, it is noteworthly that the lines selected as superior (‘Mosè’, ‘Padre Pio’ and ‘Davide’) are shifted toward the durum genome.

Percentages of the parent contributions to progenies based on 162 AFLP unmapped (A) and 53 SSR mapped markers (B) polymorphic between parents

Percentages of the average parent contributions to progenies chromosomes based on SSR data (A). Graphical representations (B) of parental contributions to chromosomes 2B and 6B of all tested progenies based on SSR mapped markers
Relationships among emmer × durum wheat derived lines and a representative set of tetraploid and hexaploid wheats
AFLP markers were used to study the relationships among emmer × durum wheat derived varieties and lines, a representative set of tetraploid wheats and some hexaploid wheats. The set of tetraploid genotypes studied included emmer Italian ecotypes and durum wheat old and new varieties widely cultivated in Italy today or in the past. An AFLP-based approach has been selected because of the high-throughput characteristics of this kind of markers. A criticism about the use of AFLP markers as a tool for evaluating genetic diversity is that they are unmapped markers, which according to some studies, are clustered in centromeric regions. However, Heun et al. (1997) have mapped 75 AFLP loci on diploid wheat, finding a distribution across all chromosomes and several other authors demostrated that the large number of polymorphisms that can be scored with this approach can reduce the source of error (Barrett et al. 1998; Bohn et al. 1999; Soleimani et al. 2002; Almanza-Pinzón et al. 2003).
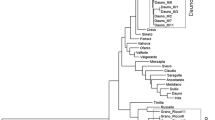
In this work, six AFLP primer pair combinations were sufficient to detect 750 markers among the 45 wheat genotypes and to univocally identify all the genotypes. Starting from AFLP data, the similarity matrix based on Simple Matching coefficient was generated, giving values of the pairwise similarity between 0.94 and 0.67. Figure 3 shows the tree constructed on this matrix using UPGMA clustering method. The relationships among emmer landraces, old and modern durum wheat genotypes widely cultivated in Italy, lines and cultivars derived from emmer × durum wheat crosses and hexaploid wheats are shown in the tree. Cluster analysis underlies the higher level of genetic diversity of emmer landraces respect to the other tetraploid wheats. In fact, the majority of durum wheat varieties is included in the cluster A. The pedigree of several of these varieties is characterized by the presence of ‘S. Cappelli’ cultivar. This genotype is in fact, directly or indirectly, present in the pedigrees of ‘Arcangelo’, ‘Capeiti’, ‘Appulo’, ‘Trinakria’, ‘Valnova’, ‘Fortore’, ‘Ofanto’, ‘Varano’ and ‘Duilio’. These cultivars have been released in Italy during the period between 1970s and 1990s as results of CYMMIT germplasm introgression into ‘S. Cappelli’ and its derivatives background (Pecetti et al. 2001).

Tree constructed from AFLP data (6 primer combinations, 750 fragments) encompassing 23 durum wheat cultivars, 4 bread wheat genotypes, 7 emmer landraces, 9 emmer × durum accessions, one spelta landrace and one Triticum tauschii genotype. Ordination of genotypes is based on Simple Matching coefficient; UPGMA clustering. The names of Triticum durum genotypes are written in green colour, blue for Triticum dicoccon, red for lines and varieties obtained from emmer × durum wheat cross, black for hexaploid wheats and violet for Triticum tauschii. Each emmer landrace is associated with a number: the same number on the map of Italy appoints the location of the collection site for this specific landrace. The two parents of all the lines and varieties derived from emmer × durum wheat cross used in this work are indicated with arrows
Cluster B includes the hexaploid varieties ‘Centauro’, ‘Vaiolet’, ‘Chinese Spring’ and the hexaploid line 28P, all characterized by the A, B and D genomes. The highest values of genetic distance are shown by Triticum tauschii, characterized by the D genome.
A third major cluster includes emmer genotypes from Central and South Italy, including ‘Molise’, tightly clustered with ‘Molisana’ and ‘Potenza’ emmer landraces. ‘Leonessa’ and ‘Umbria’ local populations, whose origins are, respectively Latium and Umbria regions, are clustered together. Among emmer genotypes, the highest values of genetic diversity are shown by the two landraces ‘Garfagnana’, from Tuscany, and ‘Abruzzo’, from Central Italy. This is in line with the results of Barcaccia et al. (2002), who found that ‘Garfagnana’ type was the most differentiated from the rest of the Italian emmer accessions studied. These authors found great variability even within ‘Garfagnana’ landrace, according to Pagnotta et al. (2005) who, in a morphological and molecular characterization of Italian emmer accessions, have shown that ‘Garfagnana’ has the highest level of within accession variation. All these authors have underlied the high level of genetic variation that can be found both among and within Italian emmer populations. In the major cluster of the dendrogram is included even the durum wheat parental ‘Simeto’, together with the lines and varieties obtained from the cross ‘Simeto’ × ‘Molise’: these, as expected, are dispersed between the parentals (indicated in the figure by arrows). The overall results underlie the high variability that can be found among emmer landraces and therefore their value as an irreplaceable bank of genetically diversified genotypes. Moreover, genetic diversity of Italian emmer populations in comparison with a wide collection of Triticum turgidum accessions from several countries has been evaluated with RFLP and SSR markers by Figliuolo and Perrino (2004). These authors found that the emmer germplam from Italy, showing high values of genetic distances, appear to belong to a primitive genepool. Therefore, the high level of genetic variability, together with the peculiar traits of emmer, make Italian landraces a valuable starting point not only for the release of emmer varieties selected from landraces, but even for broadening the genetic base of tetraploid wheat breeding germplasm through emmer × durum wheat interspecific crosses. Moreover, after an extensive characterization of the HMW glutenin subunit variations among 205 cultivated emmer accessions, Li et al. (2006) have concluded that emmer is expected to be a valuable genetic resource for quality improvement even of cultivated hexaploid wheat.
References
AACC (1995) Approved methods of the American association of cereal chemists. The Association, St Paul, Minnesota
Almanza-Pinzón MI, Khairallah M, Fox PN, Warburton ML (2003) Comparison of molecular markers and coefficients of parentage for the analysis of genetic diversity among spring bread wheat accessions. Euphytica 130:77–86
Barcaccia G, Molinari L, Porfiri O, Veronesi F (2002) Molecular characterization of emmer (Triticum dicoccon Schrank) Italian landraces. Genet Resour Crop Evol 49:415–426
Barrett BA, Kidwell KK, Fox PN (1998) Comparison of AFLP and pedigree-based genetic diversity assessment methods using wheat cultivars from the pacific northwest. Crop Sci 38:1271–1278
Bohn M, Utz HF, Melchinger AE (1999) Genetic similarities among winter wheat cultivars determined on the basis of RFLPs, AFLPs, and SSRs and their use for predicting progeny variance. Crop Sci 39:228–237
Castagna R, Minoia C, Porfiri O, Rocchetti G (1996) Nitrogen level and seeding rate effects on the performance of hulled wheats (Triticum monococcum L., T. dicoccum Schübler and T. spelta L.) evaluated in contrasting agronomic environments. J Agron Crop Sci 176:173–181
Codianni P, Ronga G, Di Fonzo N, Troccoli A (1996) Performance of selected strains of “farro” (Triticum monococcum L., Triticum dicoccum Schübler, Triticum spelta L.) and durum wheat (Triticum durum Desf cv Trinakria) in the difficult flat environment of Southern Italy. J Agron Crop Sci 176:15–21
Codianni P, Troccoli A, Galterio G, Pogna NE, Di Fonzo N (1998) Davide, prima varietà di farro migliorata geneticamente. L’Informatore Agrario 25:46–49
Codianni P, Galterio G, Pogna NE, Di Fonzo N (2000) Mosè e Padre Pio, due nuovi genotipi di farro. L’Informatore Agrario 24:37–39
Corazza L, Pasquini M, Perrino P (1986) Resistance to rusts and powdery mildew in some strains of Triticum monococcum L. and Triticum dicoccum Schübler cultivated in Italy. Genet Agraria 40:243–254
Cubadda R, Marconi E (1996) Technological and nutritional aspects in emmer and spelt. In: Padulosi S, Hammer K, Heller J (eds) Hulled wheats. First International Workshop on Hulled Wheats, 21–22 July 1995, Castelvecchio Pascoli, Tuscany, Italy. Promoting the conservation and use of underutilized and neglected crops. International Plant Genetic Resources Institute, Rome 4, pp 203–211
Damania AB, Hakim S, Moualla MY (1992) Evaluation of variation in Triticum dicoccum for wheat improvement in stress environments. Hereditas 116:163–166
De Vita P, Riefolo C, Codianni P, Cattivelli L, Fares C (2006) Agronomic and qualitative traits of T. turgidum ssp. dicoccum genotypes cultivated in Italy. Euphytica 150:195–205
D’Antuono LF, Minelli M (1998) Yield and yield components analysis of emmer wheat (Triticum dicoccum Schübler) landraces from Italy. In: Jaradat AA (ed) 3rd Int. Triticeae Symposium, May 4–8, 1997, Aleppo, Syria, Science Publisher, Inc., USA, pp 393–404
Figliuolo G, Perrino P (2004) Genetic diversity and intra-specific phylogeny of Triticum turgidum L. subsp. dicoccon (Schrank) Thell. revealed by RFLPs and SSRs. Genet Resour Crop Evol 51:519–527
Galterio G, Cappelloni M, Desiderio E, Pogna N (1994) Genetic, technological and nutritional characteristics of three Italian populations of “farrum” (Triticum turgidum spp. dicoccum). J Genet Breed 48:391–398
Heun M, Schäfer-Pregl R, Klawan D, Castagna R, Accerbi M, Borghi B, Salamini F (1997) Site of einkorn wheat domestication identified by DNA fingerprinting. Science 278:1312–1314
Li QY, Yan YM, Wang AL, An XL, Zhang YZ, Hsam SLK, Zeller FJ (2006) Detection of HMW glutenin subunit variations among 205 cultivated emmer accessions (Triticum turgidum ssp. dicoccum). Plant Breed 125:120–124
Murray HG, Thompson WF (1988) Rapid isolation of high molecular weight plant DNA. NAR 8:4321–4325
Pagnotta MA, Mondini L, Atallah MF (2005) Morphological and molecular characterization of Italian emmer wheat accessions. Euphytica 146:29–37
Pecetti L, Doust MA, Calcagno L, Raciti CN, Boggini G (2001) Variation of morphological and agronomical traits and protein composition in durum wheat germplasm from eastern Europe. Genet Resour Crop Evol 48:609–620
Perrino P, Hammer K (1982) Triticum monococcum L. and T. dicoccum Schübler (syn. of T. dicoccon Schrank) are still cultivated in Italy. Presence, collecting and action. Genet Agraria 36(3–4):343–351
Porfiri O, D’Antuono LF, Codianni P, Mazza L, Castagna R (1998) Genetic variability of a hulled wheats collection for agronomic characteristics. In: Jaradat AA (ed) 3rd Int. Triticeae Symposium, May 4–8, 1997, Aleppo, Syria. Science Publisher, Inc., USA, pp 387–392
Preston KR, March PR, Tipples KH (1982) An assessment of the SDS sedimentation test for the prediction of Canadian bread wheat quality. Can J Plant Sci 62:545–553
Röder MS, Korzun V, Wendehake K, Plaschke J, Tixier M-H, Leroy P, Ganal MW (1998) A microsatellite map of wheat. Genetics 149:2007–2023
Soleimani VD, Baum BR, Johnson DA (2002) AFLP and pedigree-based genetic diversity estimates in modern cultivars of durum wheat (Triticum turgidum L. subsp. durum Desf. Husn.). Theor Appl Genet 104:350–357
Troccoli A, Borrelli GM, De Vita P, Fares C, Di Fonzo N (2000) Durum wheat quality: a multidisciplinary concept. J Cereal Sci 32:99–113
Van Berloo R (1999) GGT: software for the display of graphical genotypes. J Heredity 90:328–329
Van de Peer Y, De Wachter R (1994) TREECON for windows: a software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Comput Appl Biosci 10:569–570
Aknowledgements
This work has been supported by the “BIOCER”, “Risorse Genetiche” and “SafeEat” projects.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Terzi, V., Morcia, C., Stanca, A.M. et al. Assessment of genetic diversity in emmer (Triticum dicoccon Schrank) × durum wheat (Triticum durum Desf.) derived lines and their parents using mapped and unmapped molecular markers. Genet Resour Crop Evol 54, 1613–1621 (2007). https://doi.org/10.1007/s10722-006-9173-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-006-9173-6