
Abstract
Pattern of diversity among fifty-nine cashew accessions of three breeding populations conserved at the Cocoa Research Institute of Nigeria, Ibadan, Nigeria, was assessed using protein-isozyme marker technique. The accessions grouped into six clusters on the dendrogram of Ward’s method of squared Euclidean distance, indicating “moderate” diversity among Nigerian cashew collections. Clustering pattern reflects the eco-geographical origin of the accessions. Closer genetic affinity was observed between Indian and Local clonal populations. The importance of electrophoresis in genetic diversity study was also elucidated.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cashew (Anacardium occidentale L.) is an important tropical tree nut crop belonging to the family Anacardiaceae and native to tropical America from Mexico and West Indies to Brazil and Peru (Nakasone and Paull 1998; Samal et al. 2003). The crop was first introduced into Nigeria in the sixteen century through the coast in Lagos around Agege and from where it gradually spread to other regions of the country (Woodroof 1967; Venkataramah 1976; Togun 1977; Ohler 1979; Topper et al. 2001). Number of cashew farmers and the areas of land cultivated to the crop in the country had increased considerably in the recent past (Topper et al. 2001). The crop is of a considerable economic importance in Nigeria because of its numerous local uses and as a source of foreign exchange earnings. It is next to cocoa in terms of contribution to gross domestic products as non-oil revenue earning in Nigeria. Cocoa Research Institute of Nigeria (CRIN), an Institute saddled with research and development mandate on cashew has developed many products from this crop. Among these products are cashew type beverages and spirits (Fetuga et al. 1975), cashew-coated chocolate, cashew-kernel-blended biscuit, cashew kernel oil, cashew juice, cashew wine and Oil from the shell called Cashew Nut Shell Liquid (CNSL) which has considerable industrial applications (Menon et al. 1998). Some of these products are been commercially produced by the small and medium entrepreneurs. It is important to mention that cashew kernel is the most important delicacy that is derived from the nuts, and it has been reported to contain about 47% fat and oil, 21% protein and 22% carbonhydrates (Fetuga et al. 1975; Nayar 1998). Moreover, Nayar (1998) reported that 82% of the fat and oil in the kernel is unsaturated and this has helps in reducing cholesterol level in the body.
Cashew production in Nigeria is constrained by low yield and variable nut yields, nut quality and susceptibility to pests and diseases, even though efforts have been made to improve this crop through selection, breeding and other agronomic measures since 1973 (Sanwo 1980). Tree yields in Nigeria are reported to range from 0–35 kg/tree; with an average yield of 800–1,000 kg/ha (Aliyu 2004; Aliyu and Awopetu 2005 (under review)). Studies have shown that over 65% of the cashew trees growing in the country are still in the wild state and these trees produce nuts with low quality that commands very low price in the global trade of raw nuts. Meanwhile, an important way to increase the productivity of the crop and improve on the quality of nuts is to select desirable genotypes from the existing variations and use identified superior materials in the breeding programmes. Genetic improvement is however limited by the lack of knowledge of genetic diversity of the existing germplasm in Nigeria.
Moreover, breeding of cashew is mostly based on traditional methods of selection of useful phenotypic and agronomic traits such as nut size, nut weight, sex ratio, colour of apple, size of the fruits, tree canopy, length of panicle and yield performance (Mneney et al. 2001). Although classical phenotype features are still extremely useful, the efficiency of selection may be reduced by environmental effects on evaluated traits. Broadening the genetic base by introducing new alleles present in exotic germplasm (Faenza et al. 1982) and a systematic exploitation of heterosis (Masawe 1994) have been suggested as means to overcome some of the problems, and this has been vigorously pursue by the Cocoa Research Institute of Nigeria by introducing exotic materials from India and Brazil in the past. Unfortunately, the introduced materials have not been characterized, documented and utilized in the breeding of the crop in Nigeria.
Identification of parental lines to exploit heterosis and to introduce valuable characters into the cashew breeding programme will require more reliable information about the level of genetic similarity of gene pools available around the world (Mneney et al. 2001). Apart from this, cultivar identification is mainly based on morphological traits. The cashew descriptor developed by IBPGR (1986) for the evaluation of cashew has been used for the morphological characterization of the 59 cashew accessions selected for this study (Aliyu 2004; Aliyu and Awopetu 2005 (under reviewed)). This approach has, however, limited usefulness as the selected agronomic and economic characters are often highly influenced by environmental conditions and conclusion drawn from such results needed to be corroborated by molecular study. Dhanaraj et al. (2002) reported that data obtained from such morphological evaluations are not always based on the assessment at genic level, and often result in duplication of accessions. In this circumstance, molecular marker techniques provide an excellent tool to study the genetic diversity and genetic relationships, and helps in identification of varieties.
Molecular markers have been successfully used to study the genetic diversity and relatedness in some of the members of Anacardiaceae like mango (Schnell et al. 1995; Ravishankar et al. 2000). Random Amplified Polymorphic DNA (RAPD) was used to study the diversity and relatedness among Tanzanian cashew accessions which revealed that the diversity among the Tanzanian cultivars is low (Mneney et al. 1998, 2001). Unfortunately, such marker (RAPD) technique is not readily available for use in this study and the choice of available protein-isozyme electrophoresis. Works of Alba et al. (2001) and Murtaza et al. (2005) have shown this technique to be useful for assessing genetic diversity and resolving taxonomic problems. This technique can confirm or supplement conventional methods used for such studies (Manzano et al. 2001). However, the advantage of electrophoretic techniques over other methods is that its highly repeatable and unique patterns of protein-isozyme bands can be obtained for each pure line cultivar or species under a given set of environmental conditions (Gates and Boulter 1979; Manzano et al. 2001).
In this study, we investigate genetic diversity among 59 cashew accessions of three populations (local, Brazilian and India) using protein-isozyme markers.
Materials and methods
Planting materials
Fifty-nine accessions of cashew comprising 3 populations; India (23), Brazil (25) and Local clones (11)) growing in the germplasm plots of Cocoa Research Institute of Nigeria (CRIN), Ibadan, were selected for this study. Table 1 showed list of the accessions and their pedigree. The Indian introductions was introduced by Western Nigerian Development Corporation (WNDC), in the 1970s, but reselected and planted in the present location in 1983 by CRIN, while the local clones were selected from farmers’ fields in Ochaja area of Kogi State, North-central region of Nigeria and planted in present location in 1986. The Brazilian materials was introduced in 1986 and planted in the CRINs’ field genebank in 1988. These cashew trees were planted in randomized complete block design of three replications with each accession represented by three entries in each block.
Protein-isozyme analysis
One gram of freshly harvested young leaves of each accession was crushed in 7.5 ml of 0.09% calcium chloride and 0.6 ml of sterile water and prechilled porcelain mortar and pestle. The mixture was immersed in ice-bath for about 1 h and the supernatant was centrifuged at 4,000 rpm in a refrigerated centrifuge for 15 minutes at 10°C. The supernatant was then filtered and three drops each of 10% mercapto-ethanol, sodium dodecyl sulphate (SDS) and sucrose were added and boiled in a water bath for 10 minutes. After cooling at room temperature, three drops of glycerol was added to each sample to weigh down the protein molecules and a drop of 0.05% bromophenol blue was also added to serve as a tracer dye. About 50 μl of each protein sample was loaded into the well containing 7% polyacrylamide gel. The resolving and stacking gels were buffered to pH 8.9 and pH 6.7 with Tris-HCl respectively (Awopetu 1982; and Atkinson et al. 1986). ProteanTM I vertical slabs gel electrophoresis apparatus was employed using a cooled vertical slab and pH 8.3 Tris-glycine electrode buffer. A constant current of 1.5 mA per gel was initially applied and after the protein was stacked in the lower gel, the current was increased to 3 mA per gel (Gates and Boulter 1979). After each run that took about 4.0 h to complete, gels were removed and stained in 0.05% Coomasie brilliant blue solution for 1 h and thereafter destained in solution containing 95% ethanol and 5% acetic acid changed thrice for 12 h.
Analysis of protein bands
The numbers of protein bands revealed by gels were recorded as present or absent. For analysis, each band was assigned a value of zero (0) when absent, or scored 1 to 4 depending upon their intensity and sharpness; 1 = large band to 4 = minor band, when present and their schematic diagrams were also drawn. Based on the matrix band frequencies and standard data, genetic variability among the accessions was examined by cluster analysis, and Ward’s linkage method (Sneath and Sokal 1973) was employed to calculate Euclidean distances and this was carried on computer programme of SPSS/PC + 11.0 version.
Results and discussion
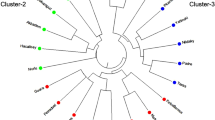
Figure 1 showed schematic representation of the zymograms of the gel electrophoresis of the 59 cashew accessions. A total of 187 protein-isozyme bands were recorded among the accessions, suggesting an average of 3.2 bands per accession. The dissimilarity matrix of the 59 cashew accessions generated by squared Euclidean distance (Fig. 2) based on the protein-isozyme banding characteristics showed that the entries grouped into six main clusters with eleven sub-clusters at 43.0% linkage distance and the summary is presented in Table 2. CSO24 and CSO17 accessions were found to span the extremes of the dendrogram with all the other accessions distributed in between at a maximum linkage distance of 99.0%.

Schematic diagram of protein-isozyme bands in the cashew accessions. (1–59 is as listed in Table 1)

A Ward’s dendrogram showing relationship among 59 cashew accessions based on protein-isozyme marker
Fifteen (15) accessions of Brazilian origin grouped together as cluster I at 43.0% linkage distance of the dendrogram. This cluster comprises of 2 sub-clusters with CSO24, CSO25, CSO22, CSO23 and CSO21 grouping together as sub-cluster A, while the remaining 10 accessions fused as sub-cluster B. Accessions in sub-cluster A are morphologically and agronomically distinct for their extreme bold-sized (jumbo) fruits (nut and apple), late and irregular flowering and fruiting characteristics. The trees are spreading biotypes with extensive branching pattern (IBPGR 1986). These accessions are also unique for low yield in terms of number of nuts per panicle (fruit clustering) and average number of nuts per tree per year. They also tend to exhibit biennial fruiting pattern. Interestingly, these five accessions clustered together in a dendrogram derived from morphological characterization study (Aliyu 2004; Aliyu and Awopetu 2006 (under review)). Juice yield from these five accessions was very high but low in astringency. Sub-cluster B accessions predominantly produced fruits with extra-large nuts and medium-sized apple. These set of cashew trees exhibited consistence fruiting habit with moderate yielding potential.
Twenty-two (22) accessions clustered as group II. These accessions further grouped into four sub-clusters (C, D, E and F). This cluster is heterogeneous as it comprises accessions from the three genetic populations, although with different number of individuals. It is important to note that fusion within this cluster tends to reflect eco-genetic relationship. The major attributes of the accessions in this cluster are prolific and high yielding with regular fruiting characteristics. Medium-sized fruits and very high shelling percentage i.e. kernel-nut-ratio with characteristic W320 and W450 kernel grades differentiated the group from others. The accessions are predominantly characterized by upright tree canopy with short period of fruit harvest. Accessions in sub-clusters D and E are distinct in combination of colour of apple and mature nuts, while sub-cluster D entries produced red apple with dark-grey nut, accessions in sub-cluster E had yellow apples with ash-grey nuts. Three (3) accessions in sub-cluster F were the best in terms of nuts yield/tree among all the 59 accessions as they produced average of 4365 nuts/tree/year and 6.34 g average nut weight, which translated to yield potential of 3.4 t/ha. The high similarity index between CC06 and CSI58 suggest that the two accessions are genetically close even though they belong to different population. Highly scented flowers, profusely flowering, mid-season fruiting, high flower sex ratio and very fertile pollen grains are the peculiar characteristics of these three accessions.
Two India introductions and a local clonal collection fused together as cluster III and they are characterized by very deep red apple with high astringent juice. The accessions were noted for production of apple juice with very high tannin content (Aliyu 2004). These set of cashew trees also exhibited secondary flowering habit and their production during the main season were very irregular. Eight (8) accessions with four Indian introductions and local clones each clustered as cluster IV. These trees produced small fruits with average weight of apple being 24 g and 3.01 g average nut weight. The apple shape is often round-compact and yellow in colour. They are unique for their sweet apple with very high content of ascorbic acids (Aliyu 2004; Akinwale and Aladesua 1999). The trees are highly prolific with yield potential of about 5,000 nuts/trees/year, but the nuts are extremely small and this result in the production of with low (small-sized) premium kernel.
Two sub-clusters I and J with three (3) and six (6) accessions in each sub-cluster respectively clustered as cluster V. The accessions are predominantly Indian introductions with large and cylindrically shaped apples. These trees commonly produce fruits with large red apples but with medium-sized nuts. Another distinct attributes of these set of accessions are high juice yield, large open-upright tree canopies with disperse main branches and early fruiting habit. Only CSO17 accession of Brazilian origin exists as a cluster VI. The accession was characterized by the highest number of bands (9). The tree is the poorest of all the selected entries in terms of flowering and fruiting. Flowering in this tree is erratic and inconsistent. It was particularly noted for its dark-greenish and rough texture leaves. It produces a green non-juicy and rough-skinned mature apple. It is however not impossible that the accession is either a mutant or has a different ploidy. The tree rarely produced more than 5 fruits in any of its production year. Tree bark is highly fissured and the canopy is open with sparse branches. The panicle is very large with very few flowers that produced low fertile pollen grains with many triads (Aliyu 2004).
Generally, from the clustering pattern showed in the dendogram, it is evident that many accessions within a cluster joined together at about 98.0% similarity linkage level, depicting high genetic similarities among such accessions. However, the clustering pattern of the accessions probably reflects the genetic history, eco-geographical origin or collection, and agronomic affinity of the selections. Awopetu (1982) opined that entries from the same ecology might have evolved similar growth forms which often reflect in the pattern of fusion in the dendrogram. This submission possibly lend credence to the clustering together of the accessions of Brazilian origin in cluster I and sub-cluster E in cluster II of the dendogram. Variation in the protein-isozyme banding pattern of the selected three cashew populations showed the extent of genetic variability of the individual accession, thereby corroborating the result obtained in the morphogenetic marker study (Aliyu 2004). This study showed close genetic affinity and proximity between the Indian cashew population and the local clonal collections than the Brazilian population. This may be attributed to the long time of cultivation of these two populations (Indian introduction and local clones) and genetic introgression among the individual accessions. Suffice that cashew was introduced into Nigeria far back in the 16th century, and the commercial cultivation dated back into 1950s’, systematic research was not initiated until 1971. During pre-research era, exchange of planting materials must have taking place among cashew farmers and this could be the reason why accessions from different geographical regions clustered together.
Results obtained in this study appear to confirm that electrophoresis is a promising tool in detecting genetic variability in plant species. Similarity of the number, position and intensity of the bands in some accessions that clustered together morpho-genetically also elucidate the potential value of the electrophoresis. This correlation between the results of electrophoresis and morphogenetic study observed was similar to the result obtained by Manzano et al. (2001) on Colocasia esculenta. The electrophoretic descriptor of the cashew protein-isozyme markers observed in this study suggests that the technique can be used as a reliable and effective characterization tool in assessment of genetic relationships involving cashew cultivars.
Variations in protein-isozyme bands obtained in this study probably suggest usefulness of the technique in the characterization of the selected cashew accessions into morphotypes. The combinations of some protein-isozyme bands observed in the experimental lots are relatively specific for a set of accessions. Moreover, some set of accessions showed exactly the same band spectra, probably suggesting that such materials are genetically close. Such accessions were observed to fuse together in the same sub-cluster at about 98.0% similarity level on the dendrograms of both multivariate (Aliyu 2004; Aliyu and Awopetu 2005 (under reviewed) and protein-isozyme analyses. Variations in the protein-isozyme bands observed point to the fact that the cashew populations in Nigeria exhibit moderate genetic base especially with the introduction of Brazilian accessions and therefore grouping of materials into distinct morphogenetic groups is not impossible.
However, the presence of common bands among the accessions of different populations depicts two things. Firstly, it probably suggests an evidence of evolutionary origin or common progenitor for the selected cashew accessions. Secondly, that the protein-isozymes are under the control of the same gene(s), and could probably be adaptive gene(s), which has/have evolved, dispersed as a result of genetic introgression and fixed in the accessions over an evolutionary time. Numerous inter- and intra-accession bands at the same distances from the cathodes reflect some level of affinity among the selected accessions. This agrees with the concept of “biochemical distances” among cultivars from the same species of known genetic relationship is a measure of affinity (Dass and Nybom 1967). It is however possible that the out-breeding nature of cashew has makes reticulate transfer of traits (proteins inclusive) from one biotype to the other. This gene flow and exchange can be responsible for common monomorphic band recorded among the selected accessions. The wide spread of some protein bands among the accessions probably attests to the effective gene exchange which was corroborated by the results of both cytological and compatibility studies (Aliyu 2004).
This study has effectively revealed the relevant of the use of protein-isozyme marker analysis in the assessment of genetic diversity in cashew. This effort is probably among the first initiative towards efficient germplasm management and utilization through characterization and identification of cultivars in a tree crop like cashew in Nigeria.
References
Akinwale TO, Aladesua OO (1999) Comparative study of the physico-chemical properties and the effect of different techniques on the quality of cashew juice from Brazilian and local varieties. Niger J Tree Crop Res 3(1):60–66
Alba E, Polignano GB, De Carlo D, Minicione A (2001) Electrophorectic phenotypes of different enzymes in some entries of Lathyrus sativus L. Lathyrus Lathyrism Newslett 2:15–20
Aliyu OM (2004) Characterization and compatibility studies in Cashew (Anacardium occidentale L.). Ph. D Thesis, University of Ilorin, Nigeria, 266 pp
Atkinson MD, Lyndsey I, Simpson MIA (1986) Characterization of cocoa germplasm using isoenzyme markers. 1. A preliminary survey of diversity using starch gel electrophoresis and standardization of procedure. Euphytica 35:741–750
Awopeu JA (1982) Genetic variability studies in Lupinus albus L. Ph. D Thesis, University of Reading, UK
Daas BJ, Nybom N (1967) In: Olsson U. Chemotaxonomic analysis in some cytotypes in Mentha × Vercillata complex (Labiatae). Botaniska Notiser 120:255–267
Dhanaraj A, Bhaskara Rao EVV, Swamy KRM, Bhat MG, Prasad DT, Sondur SN, (2002) Using RAPDs to assess the diversity in Indian cashew (Anacardium occidentale L.) germplasm. J Hortic Sci Biotechnol 77(1):41–47
Faenza V, Conticini L, Partel L (1982) Le collezioni nazional e internazional di Anacardio in Tanzania. Riv Agric Subtrop Trop 76:191–219
Fetuga BL, Babatunde GM, Ekpenyong TB, and Oyenuga VA (1975) The feeding stuff potential of cashew nut scraps kernel meal. Proceedings of the Conference of Animal Feed of Tropical and Sub-tropical origin. Tropical Products Institute, London (UK), pp 201–207
Gates P, Boulter D (1979) The use of seed isoenzymes as an aid to the breeding of field beans (Vicia faba L.). New Phyt 83:783–791
I B P G R (1986) International Board for Plant Genetic Resources. Descriptors for cashew (Anacardium occidentale L.), Rome, Italy. 33 pp
Masawe PAL, 1994: Aspect of breeding and selecting improved cashew genotypes (Anacardium occidentale L.). Ph. D. Thesis, University of Reading, UK
Menon ARR, Pillai CKS, Nando GB (1998) Effects of phosphorylated cashew nut shell lidquid on the physico-chemical properties of natural rubber. Vulcanizates 39(17):4033–4036
Mneney EE, Mantell SH, Tsoktouridis G, Amin S, Bessa AMS, and Thangavelu M (1998) RAPD – profiling of Tanzanian cashew (Anacardium occidentale L.). Proceedings of International Cashew and Coconut Conference Dar es Salaam – 1997- Tanzania, pp 108–115
Mneney EE, Mantell SH, Bennett M (2001) Use of random amplified polymorphic DNA (RAPD) markers to reveal genetic diversity within and between populations of cashew (Anacardium occidentale L.). J Hortic Sci Biotechnol 76:375–383
Murtaza N, Kitaoka M, Ali GM (2005) Genetic differentiation of cotton by Polyacrylamide gel electrophoresis. J Cent Eur Agric 6(1):69–76
Nakasone HY, Paull RE (1998) Tropical Fruits. CAB International, Oxford, UK, New York, USA, 432 pp
Nayar KG, (1998) Cashew: A versatile nut for health. Proceedings of International Cashew and Coconut Conference – 1997 – Dar es Salaam. Tanzania, pp 195–199
Ohler JG (1979) Cashew. Koninklijk instituut voor de Tropen; Teskin. Zutphen Co. Amsterdam, The Netherlands
Ravishankar KV, Anand L, Dinesh MR (2000) Assessment of genetic relatedness among mango cultivars of Indian using RAPD markers. J Hortic Sci Biotechnol 75:198–201
Manzano RA, Rodriguez Nodals AA, Roman Gutierrez MI, Fundora Mayo Z, Alfonso LC (2001) Morphological and Isozyme variability of taro (Colocasia esculenta L. Schott.) germplasm in Cuba. Plant Genet Resour Newsl 126:31–40
Samal S, Rout GR, Lenka PC (2003) Analysis of genetic relationships between populations of cashew (Anacardium occidentale L.) by using morphological characterization and RAPD markers. Plant Soil Environ 49(4):176–182
Sanwo JO (1980) Correlation coefficient between “fruits” traits as possible aids in the selection of desirable cashew (Anacardium occidentale L.). Niger J Sci 14(1):225–233
Schnell RJ, Ronning CM, Knight RJ (1995) Identification of cultivars and validation of genetic relationships in Mangifera indica L., using RAPD markers. Theor Appl Genet 90:269–274
Sneath PHA, Sokal RR (1973) Numerical taxonomy: The principles and practice of numerical classification. W. H. Freeman and Co., San Fransisco
Togun A (1977) A review of the prospect of Cashew Industry. Unpublished paper submitted to Cocoa Institute of Nigeria, 39 pp
Topper CP, Caligari PDS, Camara M, Diaora S, Djaha A, Coulibay F, Asante AK, Boamah A, Ayodele EA and Adebola PO (2001) West African regional cashew survey covering Guinea, Guinea Bissau, Cote d’Ivore, Ghana and Nigeria. Sustainable Tree Crop Programme (STCP) Report No. BHA-01109. Biohybrids Agrisystem Ltd. P. O. Box 2411, Earley, Reading RG6 5FY, UK
Venkataramah TM (1976) Cashew nut production and processing – Nigeria agronomic aspect of cashew nut production. Unpublished paper submitted to Cocoa Research Institute of Nigeria, 39 pp
Woodroof JG (1967) Tree nuts: Production, Processing, and Products. Volume I. Avi Publishing Company Incorporation, UK
Acknowledgements
The authors acknowledge the support of the Executive Director Cocoa Research Institute of Nigeria and for permission to publish this work. Technical assistance of Mr. I. A. Ogunbosoye of Plant Breeding Laboratory is also appreciated.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Aliyu, O.M., Awopetu, J.A. Assessment of genetic diversity in three populations of cashew (Anacardium occidentale L.) using protein-isoenzyme-electrophoretic analysis. Genet Resour Crop Evol 54, 1489–1497 (2007). https://doi.org/10.1007/s10722-006-9138-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-006-9138-9