
Abstract
Fatty acid translocase/cluster of differentiation 36 (FAT/CD36) functions as a membrane long-chain fatty acid transporter in various tissues in land animals. Not much is known about the CD36 molecule in teleost fish. Therefore, we studied CD36 in grass carp (Ctenopharyngodon idella, ciCD36). The full-length complementary DNA sequence of ciCD36 was 1976 bp, with an ORF of 468 amino acids, which had high sequence similarity to the CD36 of common carp. The messenger RNA (mRNA) expression of ciCD36 was high in the intestine, heart, liver, visceral tissue, and brain, but absent in the kidney. The protein expression of ciCD36 was high in the brain, intestine, liver, heart, muscle, eye, visceral tissue, gonad, and gill, but not in the kidney. Four groups of grass carp (16 tanks) were fed three times daily to satiation with 17.2 kJ gross energy/g diet (control, CON), 19.4 kJ gross energy/g diet (more energy supplied by proteins, HP), 19.9 kJ gross energy/g diet (more energy supplied by fat, HF), and 19.1 kJ gross energy/g diet (more energy supplied by carbohydrate, HC) for 11 weeks, respectively. At the end of the feeding experiment, the fish were fasted for 48 h, and the brain, heart, intestine, and liver were sampled and designated as the 0-h samples. The fish were then fed a single meal of the above four diets, and these tissues were collected at 8- and 24-h intervals after refeeding to analyze ciCD36 mRNA and protein expression levels. The results showed that at the transcriptional and translational levels, ciCD36 expression was significantly affected by refeeding time and the different diets (P < 0.05), and the regulation of its transcription in different tissues varied. At the translational level, the protein expression levels decreased in the CON and HC groups, and increased in the HP and HF groups after refeeding. The results indicated that ciCD36 has a modulatory role in the adaptation to dietary high energy in grass carp. Translational regulation might be responsible for the observed variations in ciCD36 expression.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Grass carp (Ctenopharyngodon idella) is an important economic farmed herbivorous cyprinid freshwater fish, whose total production reached 5.3768 million tons in China in 2014 (Fisheries Bureau of Agriculture Ministry of China 2015). With the development of modern high-density aquaculture industry, the high-energy feed intake or excessive feeding is a shortcut method to achieve fast growth in a relatively short period in fish. However, the excessive energy (mainly from proteins, carbohydrates, and lipids) often leads to ectopic fat accumulation and to fat deposits in the liver, mesenteric arteries, and muscle (Du et al. 2008; Weil et al. 2012). The energy requirements of grass carp are relatively low (Du et al. 2005); therefore, fatty liver is observed commonly in farmed grass carp in China. Severe fatty liver in fish results in reduced growth and feeding efficiency, impaired immune responses, and decreased nutritional quality of fish filets (Du 2014), ultimately affecting the sustainable development of the grass carp farming industry.
Fat deposition is a complex process involving lipid transport, uptake, synthesis, and catabolism. A number of membrane fatty acid (FA) transporters have been identified, including fatty acid translocase/cluster of differentiation 36 (FAT/CD36), a family of FA transport proteins (FATP1-6), and plasma membrane-associated FA-binding proteins (FABPpm) (Glatz et al. 2010). CD36 is a lipid sensor that functions in high-affinity tissue uptake of long-chain fatty acids (LCFAs) and under excessive fat supply; it contributes to lipid accumulation and metabolic dysfunction. CD36-mediated FA uptake might function in a regulatory capacity rather than as a quantitative transport mechanism (Pepino et al. 2014; Goldberg et al. 2009; Glatz et al. 2016). High hepatic CD36 levels were found in humans with nonalcoholic fatty liver disease (Greco et al. 2008). Muscles of insulin-resistant humans and of obese rodents display persistent sarcolemmal CD36 relocation and intracellular lipid accumulation (Glatz et al. 2010). The intestines of CD36-null mice showed a defect in FA uptake by the proximal intestine (Nassir et al. 2007). A high-fat (HF) diet increased plasma lipid concentrations and decreased CD36/FAT levels in rats (Sukhotnik et al. 2000; Sukhotnik et al. 2002). However, consumption of a HF diet increased liver CD36, hepatic FA uptake, and triglyceride (TG) accumulation, and these increases were prevented by CD36 deletion (Le et al. 2013). These studies suggested that deficiency or overexpression of CD36 is associated with alterations in intake and metabolism of LCFAs and is a critical factor in both ventromedial hypothalamic neuronal FA sensing and the regulation of energy and glucose homeostasis, and that CD36 is subject to tissue-specific regulation (Le et al. 2013, 2015).
However, not much is known about the functions of CD36 in teleost fish, with studies only being performed in zebrafish (Danio rerio) and common carp (Cyprinus carpio) (Fink et al. 2015). Therefore, the aim of the present study was to clone the full-length complementary DNA (cDNA) of CD36 in grass carp (Ctenopharyngodon idella, ciCD36) gene from the brain of grass carp and analyze its tissue distribution. Meanwhile, we obtained antibodies directed against ciCD36 and determined the protein levels in tissues. We then assessed the effects of dietary energy on the expressions of CD36. The results will increase our understanding of the relation between energy and lipid transport in herbivorous fish at the molecular level, and the control of fatty liver syndrome of grass carp in aquaculture by transcriptional and translational regulation.
Experimental methods
Diets
Four purified diets were prepared. Casein and gelatin were the main protein sources; equal proportions of corn oil and soybean oil were used as fat sources; and dextrin was used as the carbohydrate source. With reference to the recommendations of previous studies, dietary levels of 30–35% protein, 6–7.5% lipid, and 10.7 kJ/g digestible energy could be assumed to be suitable levels to formulate practical diets for juvenile grass carp (Köprücü 2012; Jin et al. 2013). In the present study, we added 30% protein, 8% lipid, and 36% carbohydrate to meet the demand for growth and non-specific immunity in the control diet, which represented 17.2 kJ/g gross energy, a value that has been proven as appropriate for the growth of this size of fish (Tian et al., unpublished observations). The other three diets were formulated to contain 11% higher gross energy compared with the control diet, supplied as higher protein, fat, or carbohydrate. The groups were referred to as CON (control), HP (high protein), HF (high fat), and HC (high carbohydrate), respectively. Diets were prepared by grinding the dry ingredients (<0.3 mm), weighed accurately (to the nearest 0.1 g), and mixed using a Groove-type mixer (CH-50, Changzhou Golden Ball Drying Equipment Co., Ltd., China). The oil was then blended into the mixture. After mixing, the mash was then pressed through a meat grinder (TY-432, Shang Hai Tai Yi Machinery, China) and dried to approximately 10% moisture for 8 h in a ventilated oven at 50 °C. The diets were then broken into small pieces, and sieved to obtain pellets of approximately 2.0 mm in diameter and stored at −20 °C. The ingredients and their approximate compositions are shown in Table 1.
Animals
Juvenile grass carp were obtained from and reared in an indoor recirculation aquarium system (RAS) at the Yangtze River Fisheries Research Institute (Wuhan, China). Fish were maintained in a concrete pool (3 × 3 × 5 m) with a constant flow of filtered water and a water temperature of 25–28 °C at the experimental base for a 2-week acclimatization. During the acclimatization period, fish were fed a CON diet to adjust to the experimental diets and conditions.
At the beginning of the experiment, the fish were fasted for 24 h and weighed after being anesthetized with 80 mg/L MS-222. Health and uniformly sized fish (initial weight 55.0 ± 2.5 g) were assigned randomly to 16 tanks (400 L water) with 20 fish per tank. Each diet was randomly assigned to four tanks. To reduce pellet waste, fish were slowly hand-fed until they appeared to be satiated by observing their feeding behavior, and care was taken to ensure that no uneaten food remained after feeding. The fish were fed thrice daily at 08:30, 12:30, and 17:00 (natural photoperiod). The feeding trial lasted 11 weeks. During this period, the water temperature, pH, and dissolved oxygen concentration were 25–28 °C, 7.4–7.6, and >6 mg/L, respectively.
Sample collection
To analyze the messenger RNA (mRNA) transcription and protein levels of ciCD36 in different tissues, three fish from the initial fish were sampled after feeding the control diet, and the brain, heart, gonad, kidney, spleen, eye, intestine (midgut), gill, liver, muscle, and visceral adipose tissue were separated and frozen at −80 °C.
At the end of the feeding experiment, the fish fed with the same diet were mixed and reassigned equally to three tanks, which were used for three time samples. First, the fish were fasted for 48 h, and then eight sampled fish from one tank for each treatment were designated as the 0-h sample (fasted fish). The remaining fish were fed a single meal of their allocated diet until visible satiation. Samples were collected at 8- and 24-h intervals after refeeding. At each interval, eight fish (from the same tank) were randomly taken from each treatment. Fish were sedated with 80 mg/L MS-222, disinfected with 75% alcohol, and killed. The brain, heart, intestine (midgut), and liver were placed in 2-mL microcentrifuge tubes, frozen in liquid nitrogen, and stored at −80 °C for mRNA and western blotting analyses.
Cloning of CD36 and phylogenetic analysis
PCR primers were designed to amplify the partial sequence of grass carp CD36 (ciCD36), based on the conserved CD36 cDNA sequence of zebrafish, rainbow trout, common carp, and barred knifejaw in the NCBI database. All the primers were designed using the software package Primer premier 5.0 (Table 2). A total of 100 mg of brain tissue was homogenized in liquid nitrogen, and total RNA was prepared using the TRIzol reagent. cDNA was synthesized from each RNA sample (2 μg). PCR reactions were performed in a total volume of 25 μL, including 2 μL of cDNA sample, 12.5 μL of Premix Ex Taq, 1 μL of PCR forward/reverse primers (10 μM), and 8.5 μL of nuclease-free water. The PCR conditions involved an initial denaturation at 95 °C for 3 min; followed by 30 cycles consisting of 30-s denaturation at 94 °C, 30-s annealing at 57 °C, and 45-s extension at 72 °C; with a final extension at 72 °C for 6 min. To obtain the full-length cDNA sequence, 3′ and 5′ rapid amplification of cDNA ends (RACE) were performed using the gene-specific primers (Table 2). The target products were obtained by nested PCR. The obtained products were isolated using an agarose purification kit and ligated into the pMD18-T vector. Following transfection into Escherichia coli DH5a competent cells, recombinants were identified by blue and white spot selection. Putative clones were further screened by PCR amplification, and then three selected clones were sequenced by the Sangon Biotech Company (Shanghai, China). All primers were synthesized by Sangon Biotech (Shanghai, China) and all kits were provided by Takara Biotech (Dalian, China). The sequence analysis was described in our earlier study (Cheng et al. 2011).
Expression and purification of ciCD36 in Escherichia coli and antibody preparation
The coding sequence (CDS), amplified by primers CD915F and CD915R (Table 2), was firstly subcloned into vector pGEX-4T-1 to construct the pGEX–ciCD36 construct. The expression of the glutathione-S-transferase (GST)-tagged fusion protein was induced from pGEX–ciCD36 using 0.5 mM isopropyl-b-D-thiogalactopyranoside (IPTG) at 18 °C for 10 h. pGEX-4T-1 was also used as the control. To detect ciCD36, all the collected samples were separated by electrophoresis through 12% SDS–PAGE gels, and the BandScan software was used to analyze the content of the fusion protein content. To purify the ciCD36 fusion protein, ciCD36 was expressed in E. coli BL21 (DE3) and purified by glutathione-sepharose resin. The concentrations of the soluble proteins were determined using the bicinchoninic acid assay method (BCA, (Beyotime, Shanghai, China). Finally, anti-ciCD36 antibodies were prepared by injecting the purified ciCD36 fusion protein into rabbits, and then purifying them using Protein-Sepharose CL-4B.
SYBR real-time fluorescence quantitative RT-PCR method
β-actin was used as an internal control for cDNA normalization. Using a PrimeScript® RT reagent kit, cDNA was synthesized from each RNA sample (2 μg), following the manufacturer’s recommendations. Quantitative real-time reverse transcription PCR (qRT-PCR) was conducted on the QuantStudio™ 6 Flex Real-Time PCR System (Life Technologies). Real-time PCR was carried out using a 2-μL cDNA sample, 10 μL of SYBR® Premix Ex Taq™, 0.4 μL of PCR forward/reverse primers (10 μM), and 7.2 μL of nuclease-free water. The thermocycling conditions for the reaction were as follows: 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 s, 60 °C for 15 s and 72 °C for 20 s.
Western blotting
The tissues were homogenized with liquid nitrogen and lyzed in radio immunoprecipitation assay (RIPA) buffer with 1 mM/mL phenylmethanesulfonyl fluoride (a protease inhibitor; Amresco, OH, USA) and 1 mM/mL sodium orthavanadium (Beyotime). The protein concentrations were measured using the BCA method (Beyotime). All samples were adjusted to equal concentrations. Tissue lysates (20 μg) were separated by SDS–PAGE and transferred to 0.45 μm polyvinylidene difluoride membranes (Millipore) for western blotting analysis. The membranes were blocked for 2 h with 5% nonfat dry milk (Bio-Rad) in wash buffer (Beyotime), and then probed with primary antibody overnight at 4 °C. The membranes were then washed in wash buffer, probed with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody, and detected using the ECL western blotting substrate (Thermo, MA, USA). Densitometric analyses were performed using the ImageJ software (National Institutes of Health). An anti-α-tubulin antibody was purchased from Beyotime Biotechnology Inc. (Shanghai, China). The measured protein levels of CD36 were normalized to that of α-tubulin.
Statistical analysis
Each parameter was tested for each sampled fish in triplicate. All statistical analyses were performed using SPSS 17.0 for Windows (SPSS Inc., Chicago, IL, USA). The data were expressed as means and their SD. The data for the mRNA expression profiles of ciCD36 gene in different tissues were subjected to one-way ANOVA, followed by Tukey’s multiple range tests. The data for the time course and different diets were analyzed by two-way ANOVA, testing the main effects of time (T) and diet (D), and their full factorial interaction. Tukey’s multiple range tests were conducted to detect treatment differences among the interactions. Statistical significance was defined as P < 0.05.
Results
Isolation and sequence analysis of ciCD36
Based on the 572-bp partial cDNA of ciCD36 from a grass carp brain cDNA library, two fragments with lengths of 752 and 652 bp were obtained by 5′ and 3′ RACE, respectively. After splicing and assembly using Vector NTI 11.5, the three fragments were joined into a full-length ciCD36 cDNA of 1976 bp, which contained a 56-bp 5′ UTR, a 1404-bp open reading frame (ORF) encoding a protein of 468 amino acids, and a 516-bp 3′ UTR. The predicted molecular mass was 52.65 kDa, and the molecule was predicted to be an integral membrane glycoprotein with two transmembrane domains, a large extracellular loop, and two short intracellular tails, one at the N- and one at the C-terminus. The complete nucleotide and deduced amino acids sequences of ciCD36 (GenBank Number: KU821103) are shown in Fig. 1.

The cDNA and deduced amino acid sequences of ciCD36. The open reading frame sequences are shown in capital letters. The initiation codon (ATG) and the termination codon (TAA) are shown in bold format
Phylogenetic analysis
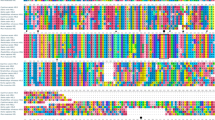
The deduced amino acid sequence of ciCD36 was subjected to a BLAST search, and similar proteins were acquired and used for amino acid sequence alignment and phylogenetic tree construction using the neighbor-joining method (Table 3, Fig. 2). The results indicated that the ciCD36 protein shared the closest relationship with that of the common carp (Cyprinus carpio), and were clustered into one group, whereas the CD36 proteins of other fishes were grouped in another cluster. All the fishes were clustered together and formed a sister group to the branches of other vertebrates; mammalian CD36 proteins were grouped in a different cluster.

Phylogenetic tree of CD36
Tissue-specific expression of ciCD36 mRNA
Tissue distribution of ciCD36 mRNA in the brain, heart, gonad, kidney, spleen, eye, intestine, gill, liver, muscle, and visceral adipose tissue of juvenile grass carp was determined by real-time PCR using β-actin as an internal control for cDNA normalization. Except in the kidney, the expression of ciCD36 mRNA was detected in all examined tissues of grass carp (Fig. 3). ciCD36 was predominantly expressed in the intestine, heart, liver, visceral adipose tissue, and brain. The highest expression level of ciCD36 mRNA was observed in the intestine.

The mRNA expression profiles of ciCD36 gene in different tissues as detected by real-time PCR. All values represented the mean ± SD (n = 3)
Tissue-specific expression of ciCD36 protein
Specific immunoreactive bands around 52 kDa were detected using the anti-ciCD36 rabbit serum by western blotting. The expression level of ciCD36 varied among the different tissues. Brain showed the highest level of ciCD36, followed by the intestine, liver, heart, muscle, eye, fat, gonad, and gill. There was no expression in the kidney (Fig. 4).

The protein expression profiles of ciCD36 gene in different tissues as detected by western blotting. All values represented the mean ± SD (n = 3)
The effect of dietary energy on ciCD36 mRNA expression
Postprandial gene expression of ciCD36 in the brain, intestine, liver, and heart in response to dietary energy were measured (Fig. 5). Statistical analysis using two-way ANOVA showed that the ciCD36 mRNA expression was significantly affected by refeeding time and the different diets (P < 0.01).

The effect of dietary energy on ciCD36 mRNA expression. (a), (b), (c), (d) mean values among all diets with unlike letters are significantly different when the interaction was significant (P < 0.05). (x), (y), (z) mean values among three time points with unlike letters were significantly different (P < 0.05). T time points, D diets, T × D interaction between T and D
In brain, the expression level of ciCD36 in the CON group was upregulated at 8 h after refeeding, whereas it was downregulated at 24 h after refeeding (P < 0.05). ciCD36 expression was significantly downregulated at 24 h after refeeding, but not at 8 h, in the HC group. In contrast, these postprandial modulations were not obvious in the HP and HF groups. Compared with the CON diet, the HP and HF diets led to significantly lower cerebral ciCD36 expression at 0, 8, and 24 h after refeeding (P < 0.05).
In the intestine, the ciCD36 levels were downregulated at 8 and 24 h after refeeding in the CON and HC groups, and at 8 h after refeeding in the HP and HF groups (P < 0.05). Compared with the CON diet, at 0 and 8 h after refeeding, no significant differences were observed for the ciCD36 expression in the HP and HF groups (P > 0.05). At 24 h after refeeding, the mRNA expressions increased in the HP and HF groups (P < 0.05).
In the liver, at 8 and 24 h after refeeding, the ciCD36 levels were upregulated in the HP and HF groups, and were downregulated in the HC group (P < 0.05). In the CON group, ciCD36 expression was significantly upregulated at 8 h after refeeding, but downregulated at 24 h after refeeding (P < 0.05). Compared with the CON diet, the ciCD36 levels were downregulated in the HF and HP groups, but not in the HC group, at 0 h after refeeding (P < 0.05); at 8 h after refeeding, the ciCD36 expressions were downregulated in all high-energy groups (P < 0.05); at 24 h after refeeding, no significant difference was observed for the ciCD36 mRNA expressions in the HC and HF groups (P > 0.05), but was significantly increased in the HP group (P < 0.05).
In heart, the ciCD36 level in the CON group at 8 h was lower compared with the level at 0 h (P < 0.05), and no postprandial modulations were observed at 24 h after refeeding (P > 0.05). In the HC and HP groups, ciCD36 expressions were significantly upregulated at 8 h after refeeding, but downregulated at 24 h after refeeding (P < 0.05). There was no change in fish fed with the HF diet (P > 0.05). Compared with the CON diet, the ciCD36 mRNA expression was lower in the HP and HF groups at 0 and 24 h after refeeding (P < 0.05) and was higher in the HC group at 0 and 8 h after refeeding (P < 0.05).
The effect of dietary energy on ciCD36 protein expression
Postprandial ciCD36 protein levels in the brain, intestine, liver, and heart are shown in Fig. 6. Statistical analysis using two-way ANOVA indicated that the ciCD36 protein levels were also affected significantly by refeeding time and the different diets (P < 0.01).

The effect of dietary energy on ciCD36 protein expression. a, b, c, d Mean values among all diets with unlike letters are significantly different when the interaction was significant (P < 0.05). x, y, z Mean values among three time points with unlike letters were significantly different (P < 0.05). T, time points; D, diets; T × D, interaction between T and D
In the brain, intestine, and liver, at 0–24 h after refeeding, the relative ciCD36 levels decreased in fish fed with the CON and HC diets and increased in fish fed with the HP and HF diets (P < 0.05). In the heart, at 8 and 24 h after refeeding, fish fed with HP and HF diets had increased relative ciCD36 levels (P < 0.05), and the fish fed with the HC diet showed a decrease in relative ciCD36 levels (P < 0.05). No significant variation was observed for the ciCD36 levels at 0–24 h after refeeding with the CON diet (P > 0.05).
Compared with the CON diet, in fish fed with the HC diet, the relative ciCD36 levels in the brain, liver, and heart increased, but decreased in the intestine at 0 h; at 8 h after refeeding; its level showed no obvious changes in the intestine and heart, increased in the brain, and decreased in the liver at 8 h. By contrast, at 24 h, the relative ciCD36 level in the intestine, liver, and heart decreased, but increased in the brain. In fish fed with the HP diet, the relative ciCD36 levels showed no significant variation at 0 h (P > 0.05), and increased in the brain, intestine, liver, and heart at 8–24 h after refeeding (P < 0.05). In fish fed with the HF diet, the ciCD36 levels showed no significant variation in the brain and liver (P > 0.05), increased in the heart (P < 0.05), and decreased in the intestine at 0 h (P < 0.05); after refeeding for 8–24 h, the ciCD36 level increased in the brain, intestine, liver, and heart (P < 0.05).
Discussion
The role of CD36 as a high-affinity pathway for cellular FA uptake and utilization, and its influence on lipid metabolism, have been well documented in mice, and the relevance of the data to humans has also been demonstrated (for a review, see (Pepino et al. 2014)). However, information is still lacking regarding the routes of lipid transport in fish. In the present study, the complete cDNA sequence of CD36 was cloned from grass carp by PCR and RACE techniques. A single CD36 gene is present in grass carp, which is similar to zebrafish, whereas there are two CD36 genes in common carp (Fink et al. 2015). Sequence conservation and characteristic motifs of CD36 were identified in the deduced amino acid sequences, and the ciCD36 protein shared its closest relationship with that of common carp, which were clustered into one group; however, it showed low interspecies similarity to mammalian CD36 proteins. Thus, it is possible that the CD36 in grass carp has other, as yet unknown, functions. Further study is needed to explore the diverse functions of CD36 in fish.
CD36 functions as a membrane LCFA transporter in various tissues, including skeletal muscle, heart, liver, adipose tissue, and the small intestine (Smith et al. 2011). Although in mammals CD36 is highly expressed in erythrocytes, monocytes, and macrophages, the expression of carp CD36 is high in the brain, ovary, and testis, but is not expressed in immune organs (Fink et al. 2015). In the present study, the expressions of ciCD36 mRNA and protein were detected in all examined tissues, except the kidney, of grass carp. The ciCD36 mRNA was predominantly expressed in the intestine, heart, liver, visceral adipose tissue, and brain. The highest expression level of ciCD36 mRNA was observed in the intestine. The brain showed the highest level of the ciCD36 protein, followed by the intestine, liver, heart, muscle, eye, fat, gonad, and gill. These high-expression tissues were associated with lipid absorption and metabolism. Previous studies in salmon and rainbow trout showed that in terms of lipid metabolism, CD36 is functionally conserved between mammals and teleosts (Torstensen et al. 2009; Sánchez-Gurmaches et al. 2011). It is probable that these tissues consume large amounts of FAs to generate energy and metabolic intermediates for their development and growth. However, CD36 is acknowledged as an important pattern recognition receptor in innate immunity (Hoebe et al. 2005; Stuart et al. 2005). In fish, the kidney (Quesada-García et al. 2014) is the main immune organ; however, CD36 gene expression is absent in the kidney of grass fish and common carp (Fink et al. 2015). These results suggested that the main function of CD36 is mediating fatty acid transfer, not innate immunity. Clearly, the exact function of fish CD36 remains a subject of further investigation.
Regulation of FAT/CD36 has been observed at the transcriptional and translational levels in mammals. FAT/CD36 mRNA abundance in the heart increases with fasting (Van der Lee et al. 2001), diabetes (Sfeir et al. 1997), neonatal heart development (Pelsers et al. 1999), high LCFAs, and high-fat feeding (Sfeir et al. 1997). In obese Zucker rats, the amount of FAT/CD36 at the heart plasma membrane was increased by 60% (Young et al. 2002). In mice and rats, CD36-mediated FA sensing is a critical factor in the regulation of energy and glucose homeostasis and fat deposition (Le et al. 2013, 2015). In fish, Atlantic salmon were fed either a diet containing fish oil (FO) or a complete replacement of FO with a vegetable oil blend (VO) for 27 months, and then fasted for 24 h before sampling; the results showed that the mRNA expression of CD36 in the white muscle was downregulated significantly in VO-fed compared with FO-fed fish (Torstensen et al. 2009). In the liver of large yellow croakers (Larimichthys crocea) fed with a low-lipid diet, CD36 mRNA expression was significantly increased (Yan et al. 2015). To date, the role of translational regulation of CD36 in fish has yet to be explored.
Numerous studies addressed the control of food intake in fish through energy intake, which is related to digestible energy in the diet (Soengas 2014). Some of the consumed nutrients (mainly from proteins, carbohydrates, and lipids) are catabolized to harness chemical (free) energy, which is required for anabolic and other life-sustaining processes. Once the minimum metabolic requirements have been met, fishes can allocate surplus energy to new biomass or to lipid storage for use as fuel in the future (NRC 2011). Grass carp is a typical herbivorous finfish without a stomach, and its energy requirement is relatively low compared with most carnivorous fish species; its relatively low capacity to endure high-energy intakes was observed in previous studies (Du et al. 2005; Yuan et al. 2013). To investigate the possible relationship between the biological functions of CD36 and dietary energy, the effects of dietary high energy on the mRNA/protein expressions of CD36 in brain, heart, intestine, and liver were carried out.
Dietary energy is one of the main factors that influence food intake in fish, and the brain controls food intake and energy balance in fish (Polakof et al. 2007; Soengas 2014). After protein consumption, peptide hormones released from the gastrointestinal tract communicate information about the energy status to the brain (Journel et al. 2012). HP meals suppress energy intake in lean and obese subjects, and obese individuals appear to be less sensitive to the satiating effects of an HF diet (Brennan et al. 2012). In the present study, the results in brain showed that, compared with the CON group, the ciCD36 mRNA expression levels in the HF group were downregulated at 0, 8, and 24 h; however, the protein levels were upregulated at 8 h and 24 h. Similar results were observed in the HP group. However, the mRNA and protein levels were upregulated by the HC diet. By contrast, CD36 expression was significantly increased over a range of glucose concentrations (medium containing 100, 200, or 600 mg/dL glucose) in macrophages derived from peripheral blood mononuclear cells (Griffin et al. 2001). This may be because fish and mammals have different mechanisms to adapt to a high-carbohydrate diet. Teleost fish are generally considered to be glucose intolerant, and increased glucose generates fat because of the obstacle of glucokinase, glucose facilitative carrier type 2, and components of the KATP channel; the glucosensing system in fish brain is activated in response to high glucose levels, and such activation probably modulates the fatty acid metabolism (Moon 2001; Yuan et al. 2013; Soengas 2014).
The heart can shift from one substrate to another depending on food intake, exercise, and pathophysiological state. The heart has an enormous capacity for FA catabolism, rapidly channeling approximately 80% of FA uptake into oxidation (Lopaschuk et al. 2010), CD36 processes approximately 50–70% of the total FA taken up in cardiomyocytes, and this large capacity explains why CD36 deletion reduces FA uptake dramatically in the heart (Hames et al. 2014; Kim and Dyck 2016). However, the structure and function of the heart in teleost fish is simpler compared with that of mammals (Laurent et al. 1983). Meanwhile, the intestine is the major organ for the digestion and absorption of lipids (Iqbal and Hussain 2009), and the liver is the key organ in lipid metabolism. In rats fed with an HF diet, decreased LCFA delivery by increased tissue CD36/FAT mRNA levels was observed (Sukhotnik et al. 2002). In CD36-overexpressing transgenic (CD36Tg) mice, overexpression of CD36 promoted glycogen synthesis, and when challenged with an HP diet, the CD36Tg mice showed an unexpected attenuation of hepatic steatosis, increased very low-density lipoprotein (VLDL) secretion, and improved glucose tolerance and insulin sensitivity (Garbacz et al. 2016). Our results showed that the ciCD36 mRNA and protein levels in the heart, intestine, and liver were significantly affected by refeeding time and the different diets, but there was no obvious pattern in the changes in the mRNA levels. This might reflect the stability of the RNA or protein in the cell, the different responses to high-nutrient diets, or translational regulation of CD36 expression. At the protein level in these three tissues, compared with the CON group, the ciCD36 levels were upregulated at 8 and 24 h in the HP and HF groups. In the HC group, the protein levels were downregulated at 8 and 24 h. The results indicated that the CD36 is regulated in a tissue-specific manner and has a modulatory role in grass carp’s adaptation to dietary high energy.
Conclusions
The present study cloned the full-length cDNA of ciCD36, which showed tissue-specific expression in grass carp. The ciCD36 mRNA and protein expression in the brain, heart, intestine, and liver were significantly affected by refeeding time courses and different energy source diets; however, there was no obvious regularity in the mRNA level, thus translational regulation might be responsible for the observed ciCD36 levels. The results indicated that ciCD36 is subjected to tissue-specific regulation and plays a modulation role in the adaptation to dietary high energy in grass carp. The results of this study will be useful for further research on the metabolic regulation of grass carp.
References
Brennan IM, Luscombe-Marsh ND, Seimon RV, Otto B, Horowitz M, Wishart JM, Feinle-Bisset C (2012) Effects of fat, protein, and carbohydrate and protein load on appetite, plasma cholecystokinin, peptide YY, and ghrelin, and energy intake in lean and obese men. Am J Physiol-Gastr L 303:G129–G140. doi:10.1152/ajpgi.00478.2011
Cheng H, Ji N, Peng Y, Shen X, Xu J, Dong Z, Wu C (2011) Molecular characterization and tissue-specific expression of the acetyl-CoA carboxylase α gene from grass carp, Ctenopharyngodon idella. Gene 487:46–51. doi:10.1016/j.gene.2011.07.027
Du ZY (2014) Causes of fatty liver in farmed fish: a review and new perspectives. J Fish China 38:1628–1638
Du ZY, Clouet P, Huang LM, Degrace P, Zheng WH, He JG, Tian LX, Liu YJ (2008) Utilization of different dietary lipid sources at high level in herbivorous grass carp (Ctenopharyngodon idella): mechanism related to hepatic fatty acid oxidation. Aquac Nutr 14:77–92. doi:10.1111/j.1365-2095.2007.00507.x
Du ZY, Liu YJ, Tian LX, Wang JT, Wang Y, Liang GY (2005) Effect of dietary lipid level on growth, feed utilization and body composition by juvenile grass carp (Ctenopharyngodon idella). Aquac Nutr 11:139–146. doi:10.1111/j.1365-2095.2004.00333.x
Fink IR, Benard EL, Hermsen T, Meijer AH, Forlenza M, Wiegertjes GF (2015) Molecular and functional characterization of the scavenger receptor CD36 in zebrafish and common carp. Mol Immunolo 63:381–393. doi:10.1016/j.molimm.2014.09.010
Fisheries Bureau of Agriculture Ministry of China (2015) China fishery statistical yearbook. Chinese Agricultural Press, Beijing in Chinese
Garbacz WG, Lu P, Miller TM, Poloyac SM, Eyre NS, Mayrhofer G et al (2016) Hepatic overexpression of cd36 improves glycogen homeostasis and attenuates high-fat diet induced hepatic steatosis and insulin resistance. Mol Cellular Biol 36:2715–2727. doi:10.1128/MCB.00138-16
Glatz JFC, Luiken JJFP, Bonen A (2010) Membrane fatty acid transporters as regulators of lipid metabolism: implications for metabolic disease. Physiol Rev 90:367–417. doi:10.1152/physrev.00003.2009
Glatz JFC, Nabben M, Heather LC, Bonen A, Luiken J (2016) Regulation of the subcellular trafficking of CD36, a major determinant of cardiac fatty acid utilization. BBA-Mol Cell Res 1861:1461–1471. doi:10.1016/j.bbalip.2016.04.008
Goldberg IJ, Eckel RH, Abumrad NA (2009) Regulation of fatty acid uptake into tissues: lipoprotein lipase- and CD36-mediated pathways. J Lipid Res 50:S86–S90. doi:10.1194/jlr.R800085-JLR200
Greco D, Kotronen A, Westerbacka J, Puig O, Arkkila P, Kiviluoto T, Laitinen S, Kolak M, Fisher RM, Hamsten A (2008) Gene expression in human NAFLD. Am J Physiol-Gastr L 294:G1281–G1287. doi:10.1152/ajpgi.00074.2008
Griffin E, Re A, Hamel N, Fu C, Bush H, McCaffrey T, Asch AS (2001) A link between diabetes and atherosclerosis: glucose regulates expression of CD36 at the level of translation. Nat Med 7:840–846. doi:10.1038/89969
Hames KC, Vella A, Kemp BJ, Jensen MD (2014) Free fatty acid uptake in humans with CD36 deficiency. Diabetes 63:3606–3614. doi:10.2337/db14-0369
Hoebe K, Georgel P, Rutschmann S, Du X, Mudd S, Crozat K, Sovath S, Shamel L, Hartung T, Zähringer U, Beutler B (2005) CD36 is a sensor of diacylglyc-erides. Nature 433:523–527. doi:10.1038/nature03253
Iqbal J, Hussain MM (2009) Intestinal lipid absorption. Am J Physiol-Endoc M 296:E1183–E1194. doi:10.1152/ajpendo.90899.2008
Jin Y, Tian L, Zeng S, Xie S, Yang H, Liang G, Liu Y (2013) Dietary lipid requirement on non-specific immune responses in juvenile grass carp (Ctenopharyngodon idella). Fish Shellfish Immun 34:1202–1208. doi:10.1016/j.fsi.2013.01.008
Journel M, Chaumontet C, Darcel N, Fromentin G, Tome D (2012) Brain responses to high-protein diets. Adv Nutr 3:322–329. doi:10.3945/an.112.002071
Kim TT, Dyck JRB (2016) The role of CD36 in the regulation of myocardial lipid metabolism. BBA-Mol Cell Biol L 1861:1450–1460. doi:10.1016/j.bbalip.2016.03.018
Köprücü K (2012) Effects of dietary protein and lipid levels on growth, feed utilization and body composition of juvenile grass carp (Ctenopharyngodon idella). Journal of Fisheriessciences Com 6:377–380. doi:10.3153/jfscom.2012028
Laurent P, Holmgren S, Nilsson S (1983) Nervous and humoral control of the fish heart: structure and function. Comp Biochem Phys A 76:525–542. doi:10.1016/0300-9629(83)90455-3
Le FC, Dunn-Meynell A, Musatov S, Magnan C, Levin BE (2013) FAT/CD36: a major regulator of neuronal fatty acid sensing and energy homeostasis in rats and mice. Diabetes 62:2709–2716. doi:10.2337/db12-1689
Le FC, Dunnmeynell AA, Levin BE (2015) Role of FAT/CD36 in fatty acid sensing, energy, and glucose homeostasis regulation in DIO and DR rats. Am J Physiol-Reg I 308:R188–R198. doi:10.1152/ajpregu.00367.2014
Lopaschuk GD, Ussher JR, Folmes CD, Jaswal JS, Stanley WC (2010) Myocardial fatty acid metabolism in health and disease. Physio Rev 90:207–258. doi:10.1152/physrev.00015.2009
Moon TW (2001) Glucose intolerance in teleost fish: fact or fiction? Comp Biochem Phys B 129:243–249. doi:10.1016/S1096-4959(01)00316-5
Nassir F, Wilson B, Han X, Gross RW, Abumrad NA (2007) CD36 is important for fatty acid and cholesterol uptake by the proximal but not distal intestine. J Biol Chem 282:19493–19501. doi:10.1074/jbc.M703330200
NRC (National Research Council) (2011) Nutrient requirements of fish and shrimp. National Academy press. Washington, DC: pp34
Pelsers MM, Lutgerink JT, Nieuwenhoven FA, Tandon NN, van der Vusse GJ, Arends JW, Hoogenboom HR, Glatz JF (1999) A sensitive immunoassay for rat fatty acid translocase (CD36) using phage antibodies selected on cell transfectants: abundant presence of fatty acid translocase/CD36 in cardiac and red skeletal muscle and up-regulation in diabetes. Biochem J 337:407–414. doi:10.1042/0264-6021:3370407
Pepino MY, Kuda O, Samovski D, Abumrad NA (2014) Structure-function of CD36 and importance of fatty acid signal transduction in fat metabolism. Annu Rev Nutr 34:281–303. doi:10.1146/annurev-nutr-071812-161220
Polakof S, Míguez JM, Soengas JL (2007) In vitro evidences for glucosensing capacity and mechanisms in hypothalamus, hindbrain, and Brockmann bodies of rainbow trout. Am J Physiol-Reg I 293:1410–1420. doi:10.1152/ajpregu.00283.2007
Quesada-García A, Valdehita A, Kropf C, Casanova-Nakayama A, Segner H, Navas JM (2014) Thyroid signaling in immune organs and cells of the teleost fish rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immu 38:166–174. doi:10.1016/j.fsi.2014.03.016
Sánchez-Gurmaches J, Ostbye TK, Navarro I, Torgersen J, Hevrøy EM, Ruyter B, Torstensen BE (2011) In vivo and in vitro insulin and fasting control of the transmembrane fatty acid transport proteins in Atlantic salmon (Salmo salar). Am J Physiol-Reg I 301:947–957. doi:10.1152/ajpregu.00289.2011
Sfeir Z, Ibrahimi A, Amri E, Grimaldi P, Abumrad N (1997) Regulation of FAT/CD36 gene expression: further evidence in support of a role of the protein in fatty acid binding/transport. Prostag Leukotr Ess 57:17–21. doi:10.1016/S0952-3278(97)90487-7
Smith BK, Jain SS, Rimbaud S, Dam A, Quadrilatero J, Ventura-Clapier R, Bonen A, Holloway GP (2011) FAT/CD36 is located on the outer mitochondrial membrane, upstream of long-chain acyl-CoA synthetase, and regulates palmitate oxidation. Biochem J 437:125–134. doi:10.1042/BJ20101861
Soengas JL (2014) Contribution of glucose- and fatty acid sensing systems to the regulation of food intake in fish. A review. Gen Comp Endocr 205:36–48. doi:10.1016/j.ygcen.2014.01.015
Stuart LM, Deng J, Silver JM, Takahashi K, Tseng AA, Hennessy EJ, Ezekowitz RA, Moore KJ (2005) Response to Staphylococcus aureus requires CD36-mediated phagocytosis triggered by the COOH-terminal cytoplasmic domain. J Cell Biol 170:477–485. doi:10.1083/jcb.200501113
Sukhotnik I, Gork S, Coran A, Chen M, Harmon C (2000) Effect of a high fat diet on fatty acid translocase (FAT/CD36) expression and fatty acid transport in isolated enterocytes in a rat model of short bowel syndrome. J Am Coll Surgeons 191:S41–S42. doi:10.1016/S1072-7515(00)00500-7
Sukhotnik I, May N, Gork AS, Chen M, Drongovski RA, Coran AG, Harmon CM (2002) Effect of bowel resection and high-fat diet on heart CD36/fatty-acid translocase expression in a rat model of short-bowel syndrome. Pediatr Surg Int 18:620–623. doi:10.1007/s00383-002-0861-9
Torstensen BE, Nanton DA, Olsvik PA, Sundvold H, Stubhaug I (2009) Gene expression of fatty acid-binding proteins, fatty acid transport proteins (cd36 and FATP) and β-oxidation-related genes in Atlantic salmon (Salmo salar L.) fed fish oil or vegetable oil. Aquac Nutr 15:440–451. doi:10.1111/j.1365-2095.2008.00609.x
Van der Lee KA, Willemsen PH, Samec S, Seydoux J, Dulloo AG, Pelsers MM, Glatz JF, Van der Vusse GJ, Van Bilsen M (2001) Fasting-induced changes in the expression of genes controlling substrate metabolism in the rat heart. J Lipid Res 42:1752–1758
Weil C, Lefèvre F, Bugeon J (2012) Characteristics and metabolism of different adipose tissues in fish. REV Fish Biol Fisher 23:1–17. doi:10.1007/s11160-012-9288-0
Yan J, Liao K, Wang T, Mai K, Xu W, Ai Q (2015) Dietary lipid levels influence lipid deposition in the liver of large yellow croaker (Larimichthys crocea) by regulating lipoprotein receptors, fatty acid uptake and triacylglycerol synthesis and catabolism at the transcriptional level. PLoS One:e0129937. doi:10.1371/journal.pone.0129937
Young ME, Guthrie PH, Razeghi P, Leighton B, Abbasi S, Patil S, Youker KA, Taegtmeyer H (2002) Impaired long-chain fatty acid oxidation and contractile dysfunction in the obese Zucker rat heart. Diabetes 51:2587–2595. doi:10.2337/diabetes.51.8.2587
Yuan X, Zhou Y, Liang X-F, Li J, Liu L, Li B, He Y, Guo X, Fang L (2013) Molecular cloning, expression and activity of pyruvate kinase in grass carp Ctenopharyngodon idella: effects of dietary carbohydrate level. Aquaculture 410–411:32–40. doi:10.1016/j.aquaculture.2013.06.009
Acknowledgements
This work was supported by the Special Basic Research Fund for Central Public Research Institutes (No. 2015JBFM33).
Author information
Authors and Affiliations
Corresponding author
Additional information
Juan TIAN and Wei LIU contributed equally to this work.
Rights and permissions
About this article

Cite this article
Tian, J., Liu, W., Gao, W. et al. Molecular cloning and gene/protein expression of FAT/CD36 from grass carp (Ctenopharyngodon idella) and the regulation of its expression by dietary energy. Fish Physiol Biochem 43, 875–888 (2017). https://doi.org/10.1007/s10695-017-0342-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-017-0342-7