
Abstract
In this study, a F2 population derived from the cross between deep-rooted variety “Moroberekan” with shallow-rooted variety “IR20” were used to identify and validate of SSR markers associated with root morphological traits. The F2 lines were divided into two groups. In the first group, 152 seedling having minimum of four tillers were chosen and separated into four plantlets to plant them in polyvinyl chloride pipes for root study under well-watered (WW) condition at maturity stage. The lines were genotyped using SSR markers. QTLs for maximum root length (MRL) and root dry weight showed co-segregation with RM472, RM7 and RM201. The same material was forwarded to next generation (F3) to validate the linked markers under both WW and low-moisture stress (LMS) conditions. These three markers were associated consistently with MRL across generations. In the second group, 1240 F2 plants were forwarded to F5 using SSD breeding method to test the effectiveness of the marker-assisted selection (MAS) method for drought resistant. The high performing genotypic group was significantly superior to low performing genotypic group for MRL, grain yield, root volume, root dry weight and root number, indicating the efficiency of MAS for root-related traits under LMS. Comparing MAS with farmer selection in F6, the results showed that MAS group means were significantly different from farmer group means for MRL, root volume, root dry weight and root number. Thus, MAS was combined with participatory selection to select five high-yielding and deep rooted promising lines. Identification of stable QTL for root morphological traits under WW and LMS conditions can aid in MAS and to introduce them into varieties with good yield potential and accepted by farmer.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice is the staple food for 2.5 billion people (Anonymous 2004). Growing rice is the largest single use of land for producing food, covering 9% of the earth’s arable land (Anonymous 2002). About half the total world rice production is rainfed, where drought is a major production constraint. Breeding for drought tolerance in rice remains a major challenge (Fukai 1999). Drought frequently causes loss of yield in rice. Grain yield can be drastically reduced if drought stress occurs during tillering or flowering stages. Hence, developing drought resistant cultivars, especially with good performance under late season drought stress is one of the major objectives in rice breeding programs (Boonjung and Fukai 1996; Pantuwan et al. 2002). In breeding programs to design drought tolerance of a crop, it is first necessary to gain an understanding of how the crop reacts to drought. It is also necessary to identify the characters associated with drought tolerance. This is best done under field conditions in the areas where the crop is grown since the seasonal timing of drought stress varies from one location to another. Resistance or tolerance to water stress in crop plants is the combined result of many interacting morphological and physiological characters. Selection and breeding for desirable root characteristics associated with drought tolerance have been practiced in rice and the differential response of rice genotypes to drought has been related to root system characters (Steele et al. 2006). Among the root morphological traits, maximum root length, root dry weight, root volume and root to shoot weight and length ratios are associated with drought tolerance in upland conditions (O’Toole and Soemartono 1981; Yoshida and Hasegawa 1982; Kanbar et al. 2009). Phenotypic selection for root morphological traits contributing drought tolerance is difficult, expensive and labour intensive. Recent developments in the molecular marker technology facilitate indirect selection of these complex traits through tightly linked markers. Identification of stable QTLs for root morphological traits across environments can aid in MAS and to introduce them into varieties with good yield potential (Lafitte and Courtois 1999; Hemamalini et al. 2000; Kanbar et al. 2002). Most of the earlier studies focusing on single marker or two flanked markers associated with a particular trait which, by mean ignoring the different types of interaction between the markers located in the same region (Steele et al. 2006).
Conventional breeding has tended to focus heavily on “broad adaptability”, the capacity of a plant to produce a high average yield over a range of growing environments and years. Unfortunately, candidate genetic material that produces very good yields in one growing zone, but poor yields in another, tends to be quickly eliminated from the breeder’s gene pool. Yet, this may be exactly what small farmers in some areas need (Ceccarelli et al. 1996). Professional breeders, often working in relative isolated from farmers, have sometimes been unaware of the multitude of preferences—beyond yield, and resistance to drought—of their target farmers. Participation of farmers in the very initial stages of breeding, when the large genetic variability created by the breeders is virtually untapped, is expected to exploit fully the potential gains from breeding for specific adaptation through decentralized selection by adding farmer’s perception of their own needs and farmers’ knowledge of the crop (Ceccarelli et al. 1997). Conventional selection procedures take long time for developing lines with suitable genes. In this study, MAS and participatory techniques were combined together to develop drought-tolerant rice varieties exploiting more efficiently and specific adaptation to the farmers own conditions and needs.
Materials and methods
Plant material
The F2, F3, F4, F5, F6 and F7 materials derived from crossing divergent parents viz, Moroberekan and IR20 were used in experiments. While, the female parent Moroberekan was an African japonica processing a deep and thick root and is tolerant to drought but relatively low yielding, the male parent is IR20, an indica type, having short stature, a shallow root system and high yielding but susceptible to drought.
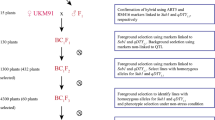
Field experiments for the present study were conducted during 2004–2007 in the experimental field of Hebbal Research Campus, University of Agricultural Sciences, Bangalore, India and farmer’s field at the Shettigere village 20 km North of Bangalore. The laboratory experiment was carried out in the Marker-Assisted Selection laboratory, Department of Genetics and Plant Breeding, University of Agricultural Sciences, GKVK, Bangalore, India. The city of Bangalore is located in the heart of the South—the Deccan Plateau, with an average elevation of 900 m above sea level, it has pleasant weather, with temperature ranging from around 24°C in winter to 35°C during summer, despite being between the very tropical latitudes of 12°39′N and 13°N, longitude being 77°37′E. The F2 seeds were divided into two sub-populations for evaluating root and yield morphological traits simultaneously (Fig. 1).

Diagrammatic representation of plan of work
Identification and validation of SSR markers associated with root morphological traits
In first sub-population, the entries were planted in nursery with 5 cm spacing between plants to produce more number of tillers at early stage of growth. Out of 272 seedlings having minimum of four tillers were chosen, 152 plants were separated into four plantlets to plant them in four replications (RCBD design) in PVC pipes measuring 1 meter in length and 20 cm diameter. The pipes were filled with a mixture of sandy clay loam and FYM in 4:1 proportion. The soil was fertilized according to the recommended package of practices. Ninety-two F2 plants were selected randomly from the 152 vegetatively replicated lines and forwarded to next generation. Phenotypic evaluation of 92 F2:3 families have been conducted in PVC pipes condition under two moisture water regimes (well-watered and low-moisture stress conditions). The experiment was laid out in an combined RCBD design for WW and LMS conditions in four replications for each.
The plants were watered daily to field capacity throughout the cropping period in WW treatment and irrigation was stopped from 70 days after sowing (DAS) up to 85 DAS in LMS treatment during tillering stage. Irrigation to field capacity was resumed and continued up to harvest. This constituted the low-moisture stress environment. During the stress period, a rainout shelter with polythene sheet was erected and a trench was dug all round the experimental plot to avoid any rainwater. Soil moisture was estimated gravimetrically (Brady, 1985) at 50 cm depth during stress period at seven different intervals. The soil moisture just before termination of stress was 20%. The sampling was done at maturity stages. Pipes at ground level were removed carefully and soaked in water overnight to loosen the soil. The next day, roots were cleaned thoroughly and carefully using a fine jet of water. The cleaned plant was collected in poly bags for recording observation. Observation consisted of maximum root length (MRL) in cm, root number at 15 cm (RN15), 30 cm (RN30) and 60 cm (RN60), root volume (RV) in cc, root dry weight (RDW) in g and grain yield/plant (GY) in g.
Genomic DNA was extracted from leaf tissues of 92 F2 lines used for the study, by the modified cetyl trimethyl ammonium bromide method (Cao and Oard 1997). DNA was quantified at 260 nm using an UV spectrophotometer. The SSR reaction mixture consisted of 1 μl of template DNA, 25 ng each of forward and reverse primers (Operon, USA), 1 mM each of dNTPs, 0.3 U of Taq polymerase (Bangalore Genei, India) and 1× PCR buffer (10 mM Tris pH 8.0, 50 mM KCl, 1.8 mM MgCl2 and 0.01 mg/ml gelatin) in a volume of 20 μl. One drop of mineral oil (Sigma, USA) was put on reaction mixture. The reactions were performed in a thermocycler with 35 cycles of 94°C for 20 s, 57°C for 30 s and 72°C for 30 s with an initial denaturation for 5 min at 94°C and a final extension at 72°C for 5 min. Amplified products were electrophoretically resolved on a 2.5% agarose gel using 1× TAE buffer.
Twenty polymorphic SSR markers, have been selected from four hot spots chromosomal regions on chromosomes 1, 3, 7 and 9, were used to screen the entries. RM302, RM212, RM265, RM315 and RM472 located on chromosome 1; RM7, RM218, RM251, RM563 and RM282 on chromosome 3; RM182, RM455, RM234, RM248 and RM420 on chromosome 7; and RM434, RM257, RM242, RM278 and RM201. The band corresponding to Moroberekan was scored as 1, the one corresponding to IR20 was scored as 3 and heterozygote was given a score of 2.
Comparative study between MAS and participatory plant selection
In the second sub-population, single-seed descent (SSD) and participatory plant selection methods were used to advance the material for the study in the farmer’s field at Shettigere village under aerobic condition (non-puddled or non-flooded soils). The details of the material developed by the breeding procedure are depicted in Fig. 1. In SSD breeding method, one seed was taken from each 1240 F2 plants to forward to F3 generation. These seeds were sown in plots of 3 × 3 m size at low density (25 cm between rows and 15 cm between plants in each row). At harvesting, one seed was taken from each plant, the seeds bulked and then used to raise the next generation of plants. The same procedure was repeated in F4. In F5, 1240 plants were grown. At maturity, 112 plants were chosen at random to represent the whole population and the seed from each of these plants harvested separately to produce a mapping population comprising 112 recombinant inbred lines (RILs) for different study. This population was screened in F6 using the five selected SSR markers (RM7, RM472, RM201, RM242 and RM257) showed strong association with drought in F2 and F2:3 to see the effectiveness of MAS. Twenty-two lines were selected based on five associated markers data and divided equally into two groups based on the banding pattern of the markers (allele type). While, in first group, the genotypes lacked the Moroberekan alleles in all the markers, the second group of genotypes possessed the Moroberekan banding patterns in most of loci. These selected lines were forwarded to next generation for comparative study.
The material of participatory plant selection involved the F3, F4, F5 and F6 generations. In F2, F3, and F4 generations, around 45 experienced rice farmers visited our field when more than 90% of the plant families were already mature. These farmers have been invited from three villages (including Shettigere) located near to the experiment field. Every participant was given a plastic label with code number and has to tie it to the selected plant. Visual selection was performed by the farmers. The best plants of farmer’s selection were forwarded to F5 generation. In F5, twenty heterogeneous inbred families (HIFs) of farmer’s selection were used as a material for marker-assisted selection.
Twenty-two genotypes were selected based on marker data from SSD population and evaluated along with lines selected by farmers for yield and root characters in F6. These lines were evaluated under WW and LMS conditions in combined RCBD design of four replications for each at Shettigere village (farmer host land) for yield study. At 50 DAS, low-moisture stress was imposed by irrigating the plants once each 12 days up to 87 DAS. The range of soil moisture level was between 25 and 35% at depth of 50 cm before giving next irrigation. In WW condition, irrigation was given once every 5 days up to harvesting. The minimum level of soil moisture reached was 65%.
Root evaluation for the same material was performed under WW and LMS conditions using PVC pipes. The experiment was laid out in combined CRD design with four replications for each WW and LMS conditions at Hebbal Campus, UAS, Bangalore, during summer season of 2006. The LMS was induced in four replications by with-holding irrigation and providing rainout shelter between 70 and 85 DAS during tillering stage. After that stress was released by supplying water daily up to maturity as explained before.
Farmer selection assisted by DNA markers
For combining MAS with participatory selection under the target environment, five promising lines from farmer selections were chosen based on banding pattern of the five associated markers with root-related traits (RM7, RM472, RM201, RM242 and RM257). The lines showed Moroberekan banding pattern in all loci have been selected and forwarded to F7 for yield and root evaluation in field and pipes respectively under WW and LMS conditions. The procedure followed for root and yield study was as explained before in F6.
Statistical analysis
The data of individuals was subjected to ANOVA to partite the variance using SAS statistical program (SAS Institute, Inc. 1996), and Duncan test was employed to classify mean values of different treatments when F-values were significant (P < 0.05). QTLs linked to markers for different phenotypic traits were determined in WW and both conditions (WW and LMS) in F2 and F2:3 populations, respectively using single-marker analysis (Tanksley et al. 1982). For phenotypic comparison between heterozygous and homozygous genotypes, the allele at the nth locus from Moroberekan is designated as Mn, where as IR20n for the allele from IR20. Single point analysis for detecting the association of a marker with a QTL lying at or close to the marker was tested using one-way analysis of variance (ANOVA). In single marker analysis each marker was considered as a treatment with three levels and phenotypic traits as the dependent variable. This analysis involved comparing for each SSR marker, the phenotypic means of three marker classes (Mn/Mn vs. IR20n/IR20n or Mn/Mn vs. Mn/IR20n or IR20n/IR20n vs. Mn/IR20n). The difference between the phenotypic means of heterozygous and homozygous marker classes was used as an estimate of the phenotypic effect of different marker genotypes. The proportion of the total phenotypic variation explained by each marker associated with a QTL (R2) was calculated as the ratio of the sum of squares examined by the marker locus to the total sum of squares. The statistical model for an F2 and F2:3 populations can be written as:
where Yij is the phenotype measurement on the j th plant in the population, μ is the population mean, gi fixed effect of marker class i and εij is the residual which includes random effect of plant genotype + measurement error for the i th plant in the j marker class. We assume εij is N (0, σ2) independent for each i and having same variance for each j.
As association between the SSR markers and the phenotypic performance of F2:3 individuals for various traits were also established using step-wise regression analysis. All SSR markers from each chromosome were treated as independent variables (Xi) in a multiple regression analysis and different traits were treated as dependent variable (Y) (root length, root volume and root dry weight). These analyses were performed on data from each condition and the mean of two environments using the GLM procedure in SAS. The final regression equation:
where Y = The response variable (MRL, RDW, RV), β0 = The intercept, β1, β2, …, βP = The partial regression coefficients corresponding to F2 and F2:3 traits, X1, X2, …, Xp = The p explanatory variables (SSR markers) and ε = The “error” (Gill 1993).
The t-test tests the hypothesis that the true means of two groups (MAS and farmer) of observations are the same. This analysis can be considered a special case of a one-way analysis of variance with two levels of classification. PROC TTEST computes the t statistic using the following formula of (Gosset 1908);
where, \( \bar{X} \) = means of MAS group (A) and farmer group (B), respectively, SE = Standard error. Standard error was calculated using the following formula:
Results and discussion
The experiment was conducted to identify a stable QTLs for root morphological traits across environments and generations in Moroberekan/IR20 cross and to combine MAS with participatory plant selection (PPS) under the target environment for developing varieties of rice tolerant to drought and accepted by the farmer.
Identification and validation of SSR markers associated with root-related traits
Making use of the single-tiller approach, replicated PVC pipe trial was being able to perform in an F2 population with four replications. Theoretically, data collected from this sort of field experiment best serve the purpose of estimation of genetic parameters and QTLs analysis; because the F2 is genetically the most informative population that enables a direct estimation of all genetic components, and replicated field trial can provide estimates and also reduce the experimental errors. The results of the analysis of variance revealed significant differences for all the traits (data not shown) in F2 and F2:3. A wide variability was observed for all root-related traits and Duncan test revealed significant differences between the two moisture regimes (WW and LMS) in F2:3 (Table 1).
Genetic variability is a prerequisite for breeders to adopt appropriate selection criteria for genetic improvement of the crop. The F2 plants exhibited wide range of variability for all the six characters (Table 1). The range in the mean values indicates the extent of phenotypic variability. Considerable amount of variability was present in the population, which is essential to increase the selection efficiency. Semi-dwarf irrigated improved indicas are known to possess a plant type of short stature, with high tiller and root number and shallow roots. On the other hand, the traditional tropical japonicas are known to possess contrasting characters. The same has been observed in this study with respect to Moroberekan and IR20. This feature of the two parents is responsible for the wide range of variation for the characters studied. The F2 population is expected to be heterozygous and heterogeneous, because of different combination of alleles in different loci in the genome. This is evident from the occurrence of the transgressants far exceeding the parental values and even the checks for several traits like maximum root length (131 cm), root volume (215 cc) and root dry weight (18 g). The mean of the F2 population fell between the parental values for different parameters and the transgressants were more than two standard deviation units away from the mean.
Based on earlier investigations (Champous et al. 1995; Price and Tomos 1997; Courtois and Nguyen 2000; Courtois et al. 2000; Shashidhar et al. 2000; Shashidhar et al. 2001; Temnykh et al. 2001; Kanbar et al. 2002; Kamoshita et al. 2002; Venuprasad et al. 2002; Kanbar et al. 2004; Steele et al. 2006), twenty polymorphic SSR markers, showed association with drought resistant in different mapping populations, were used to screen the entries. In this study, root sampling was done at maturity stage under WW in F2 and both WW and LMS in F2:3. Therefore, the QTLs identified for root-related characters are active at the end of the life cycle of rice. In F2 population, QTLs for MRL and RDW showed co-segregation with RM472, RM7 and RM201. Root volume was found to be associated with RM422 and RM201 (Table 2). Courtois et al. (2000) identified a QTL for MRL using doubled haploid lines of IR64/Azucena. One QTL was strongly associated with RZ12 on chromosome 9 which is very close to RM201 and RM242 markers. RM472, RM7 and RM201 were associated consistently with MRL across generations (Fig. 2). At the same time, RM472 and RM7 were linked consistently to RDW in F2 and F3. Only one marker, RM201 showed co-segregation consistently with root volume across generations. Yadav et al. (1997) also reported a common QTL on chromosome 9 co-segregating with RZ12-RG667 in IR64/Azucena and RZ12 in CO39/Moroberekan mapping populations. Shen et al. (2001) have replaced RZ228 and RZ12 by RM201 and RM242 for the target region of chromosome 9 to use this region in MAS for deep rooted lines in IR64/Azucena backcross population. They also selected RM234, CDO418, RZ978, CDO38 and RM248 for the target region of chromosome 7 for the same purpose. Several other scientists reported the strong association of RM201 and RM7 with MRL across populations (Shashidhar et al. 2001; Kanbar et al. 2002; Kanbar et al. 2004; Steele et al. 2006). A low statistical threshold was used in this study to avoid false-negative and due to the relatively small size of the mapping population used for genotyping, as it is difficult to detect QTLs using a high threshold in a smaller population. Price et al. (2000) found that 89% of the QTLs detected by composite interval mapping concurred with those detected by single-marker analysis (SMA).

Segregation of moroberekan/IR20 F2 population with SSR marker RM201 associated with root length
Markers within each group were analyzed as a multiple regression against data from individual generation, conditions and the combined data from both conditions (mean environment in F2:3) to determine if a region of the genome could better explain the variation for root length, root volume and root dry weight performance (Table 3). Of the four marker groups, three showed significant association with all the traits in at least one environment. The marker group on chromosome one was not showing any association with any of these three traits. Based on the result of SMA, such interaction was expected. No marker showed association with MRL, RDW and RV in F3 population under LMS condition and the linkage of markers to QTLs associated with root morphological traits was low significantly. The markers group 9 was significantly associated with MRL across generations and conditions, but was not significantly linked to root volume. The MRL locus co-segregating with RM201 and RM242 at probability of 0.001 (LOD = 3), which make this region more specific to root-related traits. Marker group 7 was significantly associated with MRL under WW in F3 and with root volume and root dry weight under LMS condition. Our findings suggest that root length-dominancy loci in chromosome regions may interact to influence extreme root length under a particular regime. The identification of gene–gene interactions may prove crucial to understanding the contributions of genes, which, by themselves, have relatively small effects on root length (deep and shallow).
In the present study, the marker groups on chromosome 3, 7 and 9 were associated with the mean environment of MRL in F3. While the marker group 7 and 3 showed association with mean environment of root dry weight and root volume, respectively. It believed that mean environment would be the most efficient condition for identification of QTLs because of the reduced standard error of trait values (Knapp et al. 1990; Venuprasad et al. 2002).
Efficiency of MAS in F6 generation
Identification of stable QTL for root morphological traits under WW and LMS conditions can aid in MAS. This would facilitate introducing them into varieties with good yield potential (Lafitte and Courtois 1999; Shen et al. 2001; Steele et al. 2006). The approach of single large-scale MAS (SLS-MAS) has been described (Ribaut and Betran 1999). This interactively combines the use of DNA markers and conventional breeding. Hence, the MAS was applied in a single and small population without flanking markers, the modified SLS-MAS has been named as single small-scale participatory-selection assisted by DNA marker in this study. In contrast to the conventional SLS-MAS, Steele et al. (2006) did not use flanking markers to reduce costs and the time required and they only attempted to identify lines with one root QTL.
Under WW condition, t-test results showed that there were no significant differences between the two performing genotypic groups (Table 4) for all the traits except grain yield (Table 5). However, the mean values of high performing genotypic (HPG) group were higher than those of corresponding low performing genotypic (LPG) group for MRL, GY/plant, RN15 and RN30. Under LMS condition, the HPG group was significantly superior to the genotypes identified as LPG group for MRL, GY, RV, RDW and RN30. However, the mean values of LPG group were numerically higher than those of corresponding HPG group for root number at 30 cm. Clearly, MAS was effective for root morphological traits under LMS condition and ineffective under WW condition. As quantitative genetic theory suggests the effectiveness of MAS is inversely proportional to the heritability of given trait (Lande and Thompson 1990; Paterson et al. 1991). The results from this study are consistent with this statement. The heritability for root morphological traits under WW was higher than the heritability under LMS condition in F6 generation (data not shown). The effectiveness of Moroberekan alleles on root related traits was more than the IR20 alleles in LMS condition where the genotypes express themselves to their fullest potential under stress condition.
Although currently MAS is used less in forward crossing than in backcrossing programs, forward MAS should become more common after novel alleles have been introgressed from exotic germplasm or wild relatives into elite genetic backgrounds. Selection for root traits by MAS could help participatory drought tolerance breeding in upland rice (Price and Courtois 1999; Hemamalini et al. 2000; Steele et al. 2006). Steele et al. (2006) have transferred five QTLs for root morphological traits from Azucena into KalingaIII using MAS.
Efficiency of MAS compared with conventional selection
Under WW condition, MAS group means were significantly different from farmer group means for MRL, RN15 and RN30 (Table 6). While under LMS condition, MAS group means were significantly different from farmer group means for MRL, RV, RDW and RN30. There is a substantial improvement for MRL and RDW, which suggests that MAS is more effective than farmer selection for drought tolerance in this rice population. The high efficiency of MAS for these traits (MRL and RDW) could be due to the strong association of the target regions with these two traits compared with other root-related traits. Additionally the genetic variation was much greater in stress treatment than non-stress condition.
Participatory plant breeding (PPB), like conventional breeding, is predicted to give a low response to selection for traits with low heritability. These include many traits that farmers consider desirable, such as drought tolerance. The technology of MAS can be used to make selection for traits with low heritability more effective (Schneider et al. 1997). Compared with participatory selection, MAS was not effective in improving yield above the experimental means under stress and non stress conditions. This result is not surprising, because the five SSR markers are very specific for root morphological characters and the MAS was done at later stages where most of the families were stable.
Combining MAS and participatory techniques
Combining MAS with PPB selection could help the breeders to develop varieties resistance to drought and accepted by the farmers. Many breeders have observed that selection efficiency decreases as the difference between the environment being selected in and the target environment increases (Ceccarelli and Grando 1999), due to high genotype x environment (G × E) interactions. Thus, varieties selected on research stations under high input conditions do not in general do well in low potential environments such as marginal farms in dry areas (Ceccarelli et al. 1998; Hollington and Steele 2007).
In this study, PPS was done under target environment in aerobic condition where uncontrolled environmental variation increases the problems. So, we have augmented MAS with PPS to improve its application in more aerobic environments and to select stable and high-yielding genotypes have a good root structure and accepted by the farmers. Based on the five SSR markers data which showed a strong association with root-related traits in F2 and F2:3 populations of the same cross, five promising lines have been selected from the farmer’s heterogeneous inbred families (HIFs) in F6. These lines were evaluated for root in PVC pipes and for yield in field condition. The result showed that a combination of five root QTLs did have a significant effect in increasing the root length of PPS lines comparing with deep rooted parent ‘Moroberekan’ (Table 7). The Moroberekan alleles in five loci have increased the root length of MIP-25-6-2 around 21% (56 cm) comparing with Moroberekan and the yield of this line was 16 g/plant. All selected lines showed root length deeper than the tolerant parent.
Selection for good root traits using stable QTLs could help drought tolerance breeding in aerobic rice and to introduce them into varieties with good yield potential (Price and Courtois 1999; Lafitte and Courtois 1999; Hemamalini et al. 2000; Steele et al. 2006). PPB has been combined with MAS (Steele et al. 2006) to introgress root QTL from the tropical japonica drought resistant rice Azucena into high yielding Kalinga III variety. Finally, primary yield trials with three or more replications have to be conducted to evaluate these lines for yield, disease, maturity time and quality etc.
Conclusions
The data obtained from this study of MAS is encouraging. Significant gains in the MAS compared with farmer selection were obtained from selection based on DNA markers linked to QTL controlling root-related traits. These results are of particular importance since the population sizes used, was relatively small, indicating that one may not need a large population size to achieve progress from MAS. In most cases, MAS provided simultaneous improvements for multiple traits (root length, root dry weight and root volume). The identification of rice selections with deep root system has been difficult because root evaluation is tedious, time consuming, relatively labor intensive and not well suited for mass screening. MAS can economically compete with PPS and the gains resulting from the reduced size and duration of breeding programs. Therefore, incorporating DNA markers into traditional breeding programs can reduce the time and money needed to achieve breeding goals.
References
Anonymous (2002) Annual report for 2001. IRRI, Los Banos, Philippines
Anonymous (2004) Annual report for 2003. IRRI, Los Banos, Philippines
Boonjung H, Fukai S (1996) Effects of soil water deficit at different growth stage on rice growth and yield under upland conditions, II. Phenology, biomass and yield. Field Crop Res 48:47–55
Brady NC (1985) The nature and properties of soils. McMillan publishing company, New York
Cao D, Oard JH (1997) Pedigree and RAPD-based DNA analysis of commercial US Rice cultivars. Crop Sci 37:1630–1635
Ceccarelli S, Grando S (1999) Decentralised participatory plant breeding. ILEIA Newsl 15:36–37
Ceccarelli S, Grando S, Booth RH (1996) International breeding programmes and resource-poor farmers: crop improvement in difficult environments. In: Eyzaguirre P, Iwanaga M (eds) Participatory plant breeding. ICARDA, Syria
Ceccarelli S, Bailey E, Grando S, Tutwiler R (1997) Decentralized-participatory plant breeding: a link between formal plant breeding and small farmers. In: Proceedings of an international seminar on a participatory research and gender analysis for technology development. Cali, Colombia
Ceccarelli S, Grando S, Impiglia A (1998) Choice of selection strategy in breeding barley for stress environments. Euphytica 103:307–318
Champous MC, Wang G, Sarkarung S, Mackill J, O’Toole JC, Huang N, McCouch SR (1995) Locating genes associated with root morphology and drought avoidance in rice via linkage to molecular markers. Theor Appl Genet 90:969–981
Courtois B, Nguyen HT (2000) Quantitative trait loci for root penetration ability and root thickness in rice: Comparison of genetic backgrounds. Genome 43:53–61
Courtois B, McLaren G, Sinha PK, Prasad K, Yadav R, Shen L (2000) Mapping QTL associated with drought avoidance in upland rice. Mol Breed 6:55–66
Fukai S (1999) Phenology in rainfed lowland rice. Field Crops Res 64:51–60
Gill JL (1993) Design analysis of experiments in the animal and medical sciences. Iowa State University Press, Ames
Gosset WS (1908) The probable error of a mean. Biometrika 6:1–25
Hemamalini GS, Shashidhar HE, Hittalmani S (2000) Molecular marker assisted tagging of morphological and physiological traits under two contrasting moisture regimes at peak vegetative stage in rice (Oryza sativa L.). Euphytica 112:69–78
Hollington PA, Steele KA (2007) Participatory breeding for drought and salt tolerant crops. In: Jenks MA et al (eds) Advances in molecular breeding toward drought and salt tolerant crops. Springer, Netherlands
Kamoshita A, Wade LJ, Ali ML, Pathan MS, Zhang J, Sarkarung S, Nguyen HT (2002) Mapping QTLs for root morphology of a rice population adapted to rainfed lowland conditions. Theor Appl Genet 104:880–893
Kanbar A, Shashidhar HE, Hittalmani S (2002) Mapping of QTL associated with root and related traits in DH population of rice (Oryza sativa L.). Indian J Genet 62(4):287–290
Kanbar A, Chandrashekara M, Manjunatha M, Shashidhar HE (2004) Molecular markers for root morphological traits under LMS using transgresant backcrosses of rice (Oryza sativa L.). Indian J Genet 64(3):185–188
Kanbar A, Toorchi M, Shashidhar HE (2009) Relationship between root and yield morphological characters in rainfed low land rice (Oryza sativa L.). Cereal Res Commun 37:261–268
Knapp SJ, Bridges WC, Birkes D (1990) Mapping quantitative trait loci using quantitative traits using RFLP linkage maps. Theor Appl Genet 79:583–592
Lafitte HRM, Courtois B (1999) Development of rice yield components in irrigated upland and lowland environments. In: Peng S, Hardy B (eds) Rice research for food security and poverty alleviation. IRRI, Los Banos, Philippines
Lande R, Thompson R (1990) Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 124:743–756
O’Toole JC, Soemartono N (1981) Evaluation of a simple technique for characterizing rice root systems in relation to drought resistance. Euphytica 30:283–290
Pantuwan G, Fukai S, Cooper M, Rajatasereekul S, O’Toole JC (2002) Yield response of rice (Oryza sativa L.) genotypes to different types of drought under rainfed lowlands. I. Grain yield and yield components. Field Crops Res 73:153–168
Paterson AH, Damon S, Hewitt JD et al (1991) Mendelian factors underlying traits in tomato: comparison across species, generations and environments. Genetics 127:181–197
Price A, Courtois B (1999) Mapping QTLs associated with drought resistance in rice: progress, problems and prospects. Plant Growth Regul 29:123–133
Price AH, Tomos AD (1997) Genetic dissection of root growth in rice (Oryza sativa L.). II: mapping quantitative trait loci using molecular markers. Theor Appl Genet 95:143–152
Price AH, Steele KA, Moore BJ, Barraclough PB, Clark LJ (2000) A combined RFLP and AFLP linkage map of upland rice (Oryza sativa L.) used to identify QTLs for root-penetration ability. Theor Appl Genet 100:49–56
Ribaut JM, Betran J (1999) Single large-scale marker assisted selection (SLS-MAS). Mol Breed 5:531–541
SAS institute, Inc (1996) SAS language guide for personal computers, 6.12th edn. Cary, North Carolina
Schneider KA, Brothers ME, Kelly JD (1997) Marker-assisted selection to improve drought resistance in common bean. Crop Sci 37:51–60
Shashidhar HE, Sharma N, Venuprasad R, Toorchi M, Hittalmani S (2000) Identification of traits and molecular markers associated with components of drought resistance in rainfed lowland rice (Oryza sativa L.). In: International rice genetics symposium, IRRI, Los Banos, Philippines
Shashidhar HE, Sharma N, Kanbar A, Hittalmani S (2001) Two DNA markers for maximum root length in rice validated across mapping populations and wide germplasm accessions. In: 8th National rice biotechnology network meeting, 25–29 October 2001, Aurangabad, India
Shen L, Courtois B, Mcnally KL, Robin S, Li Z (2001) Evaluation of near-isogenic lines of rice introgressed with QTLs for root depth through marker-aided selection. Theor Appl Genet 103:75–83
Steele KA, Price AH, Shashidhar HE, Witcombe JR (2006) Marker-assisted selection to introgress of rice QTLs controlling root traits and aroma into an Indian upland rice variety. Theor Appl Genet 112:208–221
Tanksley SD, Medina-Filho H, Rick H (1982) Use of naturally occurring enzyme variation to detect and map genes controlling quantitative traits in an interspecific backcross of tomato. Heredity 49:11–25
Temnykh S, Declerck G, Lukashova A, Lipovich L, Cartinhour S, McCouch S (2001) Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res 11:1441–1452
Venuprasad R, Shashidhar HE, Hittalmani S, Hemamalini GS (2002) Tagging quantitative trait loci associated with grain yield and root morphological traits in rice (Oryza sativa L.) under contrasting moisture regimes. Euphytica 128:293–300
Yadav R, Courtois B, Huang N, Mclaren G (1997) Mapping genes controlling root morphology and root distribution in a doubled-haploid population of rice. Theor Appl Genet 94(5):619–632
Yoshida S, Hasegawa S (1982) The rice root system: its development and function. In: Drought resistance in cereals crops with emphasis on rice. IRRI, Los Banos, Philippines
Acknowledgments
Financial assistance and training from the Rockefeller Foundation, New York, USA (RF950013#321) for Marker-Assisted Selection in Rice to Dr. Shashidhar is thankfully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kanbar, A., Shashidhar, H.E. Participatory selection assisted by DNA markers for enhanced drought resistance and productivity in rice (Oryza sativa L.). Euphytica 178, 137–150 (2011). https://doi.org/10.1007/s10681-010-0284-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-010-0284-y