
Abstract
The Polima cytoplasmic male sterility (CMS) system has been successfully used in three/two-line hybrid production in rapeseed (Brassica napus L.). However, the sterility of the Polima (pol) CMS lines is sensitive to temperature fluctuations. Also, traces of pollen can cause self-pollination within the CMS lines, which results in reduced levels of F1 hybrid seed purity and leads to a significant yield loss. Self-incompatibility (SI) is another important approach for hybrid seed production in rapeseed. Despite having a wide range of restorers and being easily selected in a breeding program, SI system has some drawbacks. In this study, SI genes from a self-incompatible line of Brassica napus were transferred to a pol CMS line and S372A, a novel line of combined cytoplasmic male sterility with self-incompatibility was bred. Due to the SI genes, this line produced very few seeds when it was selfed at low temperature and no seeds at high temperature. This suggested that the line with CMS + SI had combined the advantages and overcome the disadvantages of both the pol CMS and SI systems. Furthermore, our results showed that most of the maintainers and all the restorers of the pol CMS system were also maintainers and restorers of the CMS + SI line, respectively. This indicates that the CMS + SI system can be easily used to establish three-line hybrids of rapeseed, and we believe this novel system could be extended to other species of Brassica.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The utilization of heterosis is considered to be one of the most effective ways to increase crop yield, because there is significant heterosis in the seed yield of F1 hybrids in crops (Shen et al. 2005; Sodhi et al. 2006; Zhao and Gai 2006). Compared with other plants, Brassica species have been more successful in using heterosis because of effective and economic pollination control systems for the production of F1 hybrid seeds on a large scale. There are several approaches for hybrid breeding in rapeseed conditioned by cytoplasmic male sterility (CMS), genic male sterility (GMS), self-incompatibility (SI) and chemical hybridizing agents (CHA) (Fu et al. 1990; Fu 2000), but most have some disadvantages.
Pol CMS was discovered by Fu in 1972 (Fu et al. 1997). According to the sensitivity of the male sterility to temperature, pol CMS lines can be divided into three categories: high temperature CMS lines, low temperature CMS lines, and stable CMS lines (Fu et al. 1990). Up to now, the most successfully used in hybrid production is the high temperature lines (Fu 2000; Li et al. 2006a, b). Shan2A is another CMS line in B. napus bred by Li and Xia (1990). Testing results showed that both the pol CMS and Shan2A CMS systems had the same maintainers and restorers (Yang et al. 1998). The sterility of these two CMS lines (Shan2A and the high temperature pol CMS) is sensitive to temperature changes when seeds are sown in autumn for hybrid seed production (Zhang et al. 1997; Zhao et al. 2000). Trace pollen can result in self-pollination within the CMS lines, which is very insecure in hybrid seed production. Despite the drawbacks, the two CMS systems, especially the pol CMS system, have been used widely around the world, and it is still the main way of utilizing heterosis in rapeseed in China at the present time (Li et al. 2006a, b). In addition, the pol cytoplasm has been transferred to some vegetable Brassica species, and several vegetable pol CMS sterile lines have been developed (Yarrow 1990; Fu et al. 1997).
Unlike pol CMS and Shan2A CMS, the male sterility of GMS system is environmentally stable. However, only about 50% of the plants in the GMS lines are sterile. Therefore, all the fertile plants must be completely removed prior to flowering when using GMS lines for hybrid seed production, otherwise sib- or self-pollination can occur, which will lead to similar results as in the CMS systems (Li et al. 1995; Zhao et al. 2000).
Based on the GMS and CMS advantages, Yang and Fu (1993) and Li et al. (1995) established the genic and cytoplasmic double male sterility method. This method partially solved the problem of self-pollination within the female line, but because the sterility of about half the plants in the CMS + GMS line remained unstable and sensitive to low temperature, it was still difficult to ensure F1 seed production with high levels of hybridity using this method (Zhao et al. 2000).
SI is a widespread mechanism in flowering plants that prevents inbreeding and promotes outcrossing (Liu 1985; Banks and Beversdof 1994). Since Olsson (1960) first noticed the phenomenon in rapeseed, SI has been considered an important way for hybrid seed production (Fu 2000). However, there are some limitations with SI based heterosis, such as the difficulty to produce seeds of SI lines. Gertz and Wricke (1991) reported that high temperature could induce pseudo-compatibility in rye (Secale cereale L.), and Wilkins and Thorogood (1992) observed that SI in perennial ryegrass (Lolium perenne L.) could be broken at high temperature. We once observed the breakdown of SI in Brassica napus at a relatively high temperature in late March in 1999 in Wuhan (unpublished). This potential temperature-induced pseudo-compatibility might be a disadvantage of the SI for F1 seed production.
To combine the advantages and overcome the disadvantages of the CMS and SI systems, we transferred recessive SI genes from an SI line of Brassica napus to a pol CMS line and developed a novel line of cytoplasmic male sterility with SI. In this paper, we propose the approach of CMS + SI to utilizing heterosis in oilseed rape and have screened the maintainers and restorers for the CMS + SI system. The possible methods of applying the system are also discussed.
Materials and methods
Plant materials
The high temperature pol CMS 1141A and self-incompatible line SI-1300 of Brassica napus used in this study were bred by Professor Fu (Fu et al. 1997; Ma et al. 1998). Testing varieties/lines used are listed in Table 1. Ms and Rs were known maintainers and restorers for pol CMS, of which M3, M4, M5 and R1, R2, R3 were provided by Dr. Tian. The other testing varieties/lines were taken from the Genetic Germplasm Banks in Huazhong Agricultural University and Oil Crops Research Institute, Chinese Academy of Agricultural Sciences.
Testing experiment
Female parents from the pol CMS line, the developed cytoplasmic male sterility with self-incompatibility (CMS + SI) line and the SI line were crossed with the 22 testing varieties/lines by bud-pollination to produce F1 hybrids in March, 2005. To test the F1 male fertility, the 66 F1 hybrids, together with the pol CMS, SI and CMS + SI lines, were sown two rows each in a plot of two rows 2.33 m length and 0.27 m apart on the Experimental Farm of Oil Crops Research Institute. Thirty two days later, the plants were thinned to about 20 seedlings per row. During flowering, the F1 plants were examined for male fertility. The main inflorescences with several upper branches of all the plants in the testing experiment were bagged to produce selfed seeds. At maturity, all plants of the hybrids between self-incompatible, SI + CMS lines and testing varieties/lines were harvested to count the number of seeds, and the self-compatibility index (SCI) of each plant was calculated using the formula: SCI = number of seeds/number of flowers (Liu 1985). Finally, the SCI of an F1 hybrid was the mean of the plot. For the pol CMS system, the F1 male fertility was scored in three classes: maintained sterility (M, trace or no pollen at low temperature), partially maintained sterility (PM, trace pollen even at high temperature) and restored fertility (R, produced normal amount of pollen) (Fu et al. 1990). For the SI system, the F1 male fertility was scored in two classes: maintained self-incompatibility (M, SCI ≤ 2.0) and restored fertility (R, SCI > 2.0).
Results
Development of CMS + SI line
During the flowering time in March 2000, a female parent of the high temperature pol CMS line 1141A was crossed to a male parent of the SI line SI-1300. Because SI-1300 was the maintainer of 1141A, the F1 of the two was sterile. As the backcrossing parent, SI-1300 was crossed with the F1 plants in the summer of 2000. In the spring of 2001, the main inflorescence of the BC1F1 plants were covered with paper bags and hand-pollinated with trace pollen at the early flowering stage to test the self-compatibility of the plant. Several branches were also used to cross with SI-1300 by bud pollination. Seeds from the self-incompatible plants were harvested at the end of the rapeseed growing season. A third backcross was made in the summer of 2001, and the plants of the BC3F1 were tested for self-incompatibility and selfed by bud pollination in the flowering stages from September 2002 to May 2004. At maturity, seeds on individual selected plants were harvested. In March 2005, a line with combined cytoplasmic male sterility and self-incompatibility, named S372A, was selected based on the field experiments for male sterility and agronomic characteristics. The development of pol CMS with self-incompatibility line S372A is outlined in Fig. 1.

The development of pol CMS + SI line S372A
Exhibition of CMS + SI line at low and high temperature
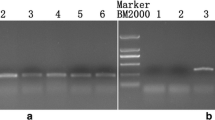
Phenotypically, the CMS + SI line S372A at seedling and bolting stages exhibited leaf shape and color similar to the backcross parent, SI-1300. During flowering, the petals of the plant were small and just overlapping, similar to 1141A. At early flowering stage (low temperature), the plants possessed normal stigmata and almost normal anthers. The quantity of pollen of S372A was less than that of SI-1300 and a little more than that of 1141A (Fig. 2). At late flowering stage (high temperature), the stamens of the S372A plants had narrow anthers with no pollen or only a trace of abnormal pollen (Fig. 2). Because the S372A line had the genes for both self-incompatibility and male sterility, the plants gave almost no selfed seeds regardless of whether the temperature was high or low (Fig. 2).

Exhibition of S372A at low and high temperature
Maintainers of S372A
Eight varieties/lines were the maintainers for the pol CMS line 1141A, of which, six testing varieties/lines were also the maintainers for the CMS + SI line S372A (Table 2). The F1 hybrids between S372A and the maintainers possessed stigmata, petals, anthers and pollen just as those of S372A described above. Under the paper bag condition, the F1 hybrids between S372A and its maintainers produced far fewer selfed seeds than the hybrids between 1141A and its maintainers and SI-1300 and its maintainers (Table 2; Fig. 3).

Podding display of the testing result as autumn sowing in Wuhan
Restorers of S372A
As shown in Table 2, all the restorers of pol CMS line 1141A were restorers of S372A. The F1 hybrids between S372A and the restorers possessed normal anthers, stigmata, and pollen, and produced many selfed seeds as normal (Fig. 3).
Discussion
Some rapeseed breeders believed that the more trace pollen the high temperature pol CMS (or Shan2A) lines had at low temperature, the higher yield of F1 seed the CMS lines produced and the stronger heterosis the hybrids obtained from the CMS lines had. This belief might account for why the pol CMS (Shan2A) system is still the main way to utilize heterosis in rapeseed. In fact, the hybrids from the stable pol CMS lines and the restorers generally produce less heterosis than those from the high temperature CMS lines and the restorers. In the growing areas of the winter- and semi-winter- type rapeseed (seeding in autumn), the high temperature pol CMS and Shan2A lines very often produce traces of pollen at low temperature during the flowering time (Zhao et al. 2000; Fan et al. 2002). When the CMS lines were used to produce F1 hybrid seeds, the sibs/selfs in the CMS hybrid seeds were about 20–30% on average (Zhao et al. 2000). It is clear in Table 2 and Fig. 3 that the progeny between the pol CMS line and the maintainer produced many selfed seeds at a low temperature. In some cases, the sib/self seeds would reduce hybridity levels of F1 hybrid seeds and significantly decrease the yield of the hybrid cultivar in a large area (Liu and Yang 1992). In China there are currently two methods to prevent the CMS lines from producing sib/self seeds: one is to do tip pruning at early flowering stage in the case of autumn seeding, and the other is to sow in spring in the area of high elevation in the Northwest (the F1 hybrid seeds are transported to the South and used in the next year). These methods are actually quite costly and time consuming. For the self-incompatibility system, the sibs/selfs in the SI hybrid seeds are about 10% (Shen et al. 2004), however the progenies from the line S372A combined cytoplasmic male sterility with self-incompatibility and could produce much fewer selfed seeds than those from pol CMS or SI systems at either low or high temperatures (Fig. 3; Table 2). The self-pollination of the CMS and SI lines will reduce hybridity levels of the hybrid seed and, consequently, lower heterosis for seed yield of the hybrid (Fu 2000). Therefore, the CMS + SI system might be very helpful to hybridity and heterosis of F1 hybrid.
In hybrid breeding, an effective and economic pollination control system is necessary for the production of F1 hybrid seed. Since male sterility has been considered to be the most effective pollination control system and is widely used in the application of heterosis in crops (Fu 2000; Zhao and Gai 2006), breeders have been trying to discover a new male sterile system (Yang and Fu 1993; Meng et al. 1995; Zhao et al. 2000). However, it would take a long time to elucidate the mechanisms of male sterility and find the maintainers and the restorers before the new system can be used in agricultural practice (Meng et al. 1995). Therefore, it is worth improving the currently available systems. The CMS + SI system we proposed here is such a system. The inheritance of the pol CMS and self-incompatibility of the SI line SI-1300 is clear (Fu 2000; Yang et al. 2001). As stated in the results section of this paper, the system could be promptly put into application because the maintainers and restorers for the system are basically ready to use (most of the maintainers and all the restorers for the pol CMS system were the maintainers and restorers of the CMS + SI system).
New maintainers and restorers are imperative for the development of new hybrid cultivars with high yield and good qualities. In the results section of this paper, we only presented the methods used to screen the maintainer(s) and restorer(s) for the CMS + SI system from the bred lines or cultivars, especially from the known maintainers and restorers for the pol CMS system. Actually, new maintainer(s) or restorer(s) of the system could be developed by using the schedules described as Fig. 4 or 5.

Development of new maintainer lines for the CMS + SI system based on cross-breeding

Development of a new restorer line for the CMS + SI system based on backcross-breeding
The CMS + SI line reproduction and F1 hybrid seed production were similar to the pol CMS system.
Pol cytoplasm has been transferred to Brassica vegetable species, and many vegetable pol CMS sterile lines have been developed, such as pol-Brassica rapa vegetable lines (Zhuang and Dong 1993; Fu et al. 1997) and a pol-Brassica oleracea line (Yarrow 1990). Because of the sensitivity of pol CMS in Brassica napus to low temperature, most of the pol CMS lines in vegetable Brassica species are also sensitive to temperature changes (Zhuang and Dong 1993). This is unfavorable for F1 seed production. In fact, more than 80% of the species of the Brassicaceae family are self-incompatible (Liu 1985). It might be easier to breed a line of pol cytoplasmic male sterility with self-incompatibility in vegetable Brassica species than in rapeseed (Brassica napus). This new strategy may be very useful in F1 hybrid vegetable seed production.
References
Banks PP, Beversdorf WD (1994) Self-incompatibility as a pollination control mechanism for spring oilseed rape, Brassica napus L. Euphytica 75:27–30
Fan LY, Chen WJ, Qu L, Li M (2002) The main affecting factors and the solving ways about seed purity in hybrid rape seed production (Brassica napus L) by CMS system in Hunan province. Hunan Agri Sci 2:8–11
Fu TD (2000) Breeding and utilization of rapeseed hybrid, 2nd edn. Hubei Science and Technology Press, Wuhan
Fu TD, Yang GS, Yang XN (1990) Studies on “three line” polima cytoplasmic male sterility developed in Brassica napus L. Plant Breeding 104:115–120
Fu TD, Yang GS, Yang XN, Ma CZ (1997) Discovery, study and utilization of polima cytoplasmic male sterility in Brassica napus L. Prog Nat Sci 5:169–177
Gertz A, Wricke G (1991) Inheritance of temperature-induced pseudo-compatibility in rye. Plant Breeding 107:89–96
Li DR, Xia RZ (1990) Genetic researches on the selection of male sterile trilines in Brassica napus L. Crop Res 4(3):20–26
Li DX, Rao R, Zhang TP (2006b) Breeding of a new Brassica napus cultivar Qianyou 16. Seed 25(5):85–87
Li J, Tang Z, Chen L (1995) Breeding a genic and cytoplasmic double-MS line of rapeseed (Brassica napus L). Plant Breeding 114:552–554
Li YC, Hu Q, Mei DS, Li YD, Xu YS, Zhao Q (2006a) Breeding of Zhongyouza 8, a canola variety with large seeds and high oil yield. Agri Sci China 5:257–264
Liu HL (1985) Genetics and breeding of rapeseeds. Shanghai Publishing House of Science and Technology, Shanghai
Liu DZ, Yang YZ (1992) Preliminary study on seed production of rape Qinyou No 2 in winter. Seed 10:45–46
Ma CZ, Fu TD, Yang GS, Tu JX, Yang XN (1998) Breeding for self-incompatibility lines with double zero in Brassica napus L. J Huazhong Agri Univ 17:211–213
Meng JL, Gan L, Cheng BF (1995) Two new cytoplasmic male sterile lines of Brassica napus bred through interspecific hybridization. J Huazhong Agri Univer 14:21–25
Olsson G (1960) Self-incompatibility and outcrossing in rape and white mustard. Hereditas 46:241–252
Shen JX, Lu GY, Fu TD, Yang GS, Wei ZL (2004) Purity analysis on hybrid of self-incompatibility in Brassica napus L. Chin J Oil Crop Sci 26:12–15
Shen JX, Fu TD, Yang GS, Ma CZ, Tu JX (2005) Genetic analysis of rapeseed self-incompatibility lines reveals significant heterosis of different patterns for yield and oil content traits. Plant Breeding 124:111–116
Sodhi YSA, Chandra A, Verm JK, Arumugam N, Mukhopadhyay A, Gupta V, Pental D, Pradhan AK (2006) A new cytoplasmic male sterility system for hybrid seed production in Indian oilseed mustard Brassica juncea. Theor Appl Genet 114:93–99
Wilkins PW, Thorogood D (1992) Breakdown of self-incompatibility in perennial ryegrass at high temperature and its uses in breeding. Euphytica 64:65–69
Yang GS, Fu TD (1993) A potential way of utilizing heterosis in rapeseed -genic and cytoplasmic male sterility (GCMS). J Huazhong Agri Univer 12:307–316
Yang GS, Chen CB, Zhou GL, Geng CN, Ma CZ, Tu JX, Fu TD (2001) Genetic analysis of four self-incompatible lines in Brassica napus. Plant Breeding 120:57–61
Yang GS, Fu TD, Brown GG (1998) The genetic classification of cytoplasmic male sterility systems in Brassica napus L. Sci Agri Sin 31:27–31
Yarrow SA (1990) The transfer of “Polma” cytoplasmic male sterility from oilseed rape (B napus) to broccoli (B oleracea) by protoplast fusion. Plant Cell 9:185–188
Zhang DF, Feng L, Li FY (1997) Some investigation on the way for raising cross-rate in producing seed of three-line sensitive to lower temperature in Brassica napus. Oil Crops China 19(2):1–4
Zhao Y, Wang ML, Zhang YZ, Du LF, Fan T (2000) A chlorophyll-reduced seedling mutant in oilseed rape, Brassica napus, for utilization in F1 hybrid production. Plant Breeding 119:131–135
Zhao TJ, Gai JY (2006) Discovery of new male-sterile cytoplasm sources and development of a new cytoplasmic nuclear male-sterile line NJCMS 3A in soybean. Euphytica 152:387–396
Zhuang SQ, Dong ZS (1993) Breeding of allocytoplasmic male sterile line in Brassica campestris L. J Huazhong Agri Univer 12:551–555
Acknowledgements
The authors thank Dr. Xiaoping Fang for assistance with the photos of the CMS + SI line S372A. Dr. Lecong Zhou and Carol Volker are acknowledged for editing and critique of the manuscript. This work was financially supported by funds from the Hi-Tech Research and Development Programs of China (2003AA207150).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Shen, Jx., Wang, Hz., Fu, Td. et al. Cytoplasmic male sterility with self-incompatibility, a novel approach to utilizing heterosis in rapeseed (Brassica napus L.). Euphytica 162, 109–115 (2008). https://doi.org/10.1007/s10681-007-9606-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-007-9606-0