
Abstract
A hydroponic experiment was carried out to investigate possible hormetic response induced by cadmium (Cd) in a potential hyperaccumulator-Lonicera japonica Thunb. The results showed that Cd at low concentrations induced a significant increase in plant growth, leaf water content and content of photosynthetic pigments in L. japonica, but decreased them at high concentrations, displayed inverted U-shaped dose response curves, confirming a typical biphasic hormetic response. The U-shaped dose response curves were displayed in malondialdehyde (MDA) and electrolyte leakage in leaves at low doses of Cd, indicating reduce oxidative stress and toxic effect. The increase of superoxide dismutase (SOD) and catalase (CAT) activities was observed along with the increased Cd concentration, indicative of increase in anti-oxidative capacity that ensures redox homeostasis is maintained. After 28 days exposure to 10 mg L−1 Cd, stem and leaf Cd concentrations reached 502.96 ± 28.90 and 103.22 ± 5.62 mg kg−1 DW, respectively and the plant had high bioaccumulation coefficient (BC) and translocation factor (TF′). Moreover, the maximum TF value was found at 2.5 mg L−1 Cd treatment, implying that low Cd treatment improved the ability to transfer Cd from medium via roots to aerial structures. Taking together, L. japonica could be considered as a new plant to investigate the underlying mechanisms of hormesis and Cd tolerance. Our results suggest that hormetic effects should be taken into consideration in phytoremediation of Cd-contaminated soil.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy metals are environmental contaminants emanating from industrial and agricultural activities such as mining and smelting of metalliferous ores, wastewater irrigation and abuse of chemical fertilizers and pesticides (Wu et al. 2006; Sun et al. 2009). Among them, Cd has attracted the most attention due to its great toxicity and high mobility from soil to plants and further down the food chain (Vig et al. 2003). Although there is no known biological function of Cd in plants, this metal is readily taken up by roots and translocated into aerial organs where it can accumulate to high levels (Baryla et al. 2001). Excessive amount of Cd to plants has long been known to suppress growth, reduce chlorophyll content (Chen et al. 2011), damage cell membrane integrity (Skórzyńska-Polit et al. 2010) and perturb water homeostasis (Poschenrieder et al. 1989). Cd toxicity also easily cause oxidative stress in plant cells, because Cd can effectively trigger the synthesis/accumulation of reactive oxygen species (ROS), which can cause cellular damage or lipid peroxidation and inhibit the activities of antioxidant enzymes involved in the oxidative defense system (Shah et al. 2001). These adverse effects of Cd on organisms are often reported at relatively higher doses and show linear or nearly linear dose–response relationship.
However, some chemicals, although toxic at higher doses, can be stimulatory at low doses. This biphasic dose–response phenomenon has been termed as hormesis or hormetic effect by many authors (de la Rosa et al. 2004), which are always characterized by J- or inverted U- shaped dose response curve, depending on the different endpoints measured. For example, in the cases of normal physiological functions, such as growth, cell proliferation, memory and longevity, hormetic responses have typically been graphed as an inverted U-shaped dose–response curve, which represents an increase above the control at low doses followed by a decrease at higher doses. On the contrary, if the endpoints denote some dysfunction such as disease incidence, carcinogenesis, mutagenesis, hormetic effects are typically graphed as a J-shaped dose response, which represents a decrease compared with the control at low doses followed by an increase at higher doses (Calabrese and Baldwin 2003a, b).
Some investigations suggest that hormesis is a rule rather than an exception, which represents an evolutionary-based adaptive response to environmentally induced disruption in homeostasis (Stebbing 1998; Calabrese and Baldwin 2001; Calabrese et al. 2006, 2008, 2010). Though the biphasic dose–response is a common result of experiments, the low-dose data have been largely ignored, and the prevailing view is that it is important to reduce level of toxins as much as possible (Mattson and Calabrese 2010). Potential stimulation at low concentrations has received little attention. Cd as one of the most hazardous heavy metals in soil, much lower than concentrations of other common heavy metals such as lead, copper, zinc, nickel. During the past decades, although Cd has been found induces hormetic responses in a number of organisms (Calabrese and Baldwin 1999), most of the studies are more concerned about the toxic effects of Cd at high concentrations. Therefore, much less information is available about the effect of Cd at low concentrations.
Japanese honeysuckle (Lonicera japonica Thunb), a popular ornamental, has become established in temperate and tropical regions worldwide in the past 150 years, and it is also widely used in Asian medicine (Larson et al. 2007). In our previous work, it was shown L. japonica had strong tolerance to Cd and strong accumulation capability in its stem, and could be seen as a potential Cd-hyperaccumulator. When exposed to low concentrations, the plants did not show any visual symptoms, even the plant growth seemed to be improved (Liu et al. 2009; Jia et al. 2012), like the hormetic effect described by many authors. But the evidence was not very strong. Thus, the present study was aimed to retest the possible hormetic effect in L. japonica at low Cd concentrations with various endpoints. Furthermore it may provide an enlightenment of consider the hormetic effect in the phytoremediation Cd-contaminated soil.
Materials and methods
Plant culture and Cd exposure
Cuttings of L. japonica were collected from a non-contaminated field–Shenyang Arboretum of Chinese Academy of Sciences and propagated in perlite in the laboratory. After 2 months, plants were transformed to slightly modified Hoagland solution (Hoagland and Arnon 1950) containing the following ingredients (mmol l−1): Ca (NO3)2·4H2O 5.00, MgSO4·7H2O 2.00, KNO3 5.00, KH2PO4 1.00, H3BO3 0.05, ZnSO4·7H2O 0.80 × 10−3, MnCl2·4H2O 9.00 × 10−3, CuSO4·5H2O 0.30 × 10−3 (NH4)6Mo7O24·4H2O 0.02 × 10−3, Fe-EDTA 0.10. Uniform plants were cultivated in black plastic boxes (15 plants per box) containing the culture medium with continual aeration; the volume of solution in each box was 1.5 L. The experiment was performed in a growth chamber under controlled conditions: 14 h illumination per day (5.00 a.m. to 7.00 p.m.), 800 μmolm−2 s−1 PPFD, with a 25 ºC/18 ºC day/night temperature and a relative humidity of 60 %. Solutions were renewed once every 3 days to prevent nutrient depletion, and the pH was daily adjusted to 5.8 ± 0.1 with 0.1 M HCl or 0.1 M NaOH. Plants, which had been cultivated hydroponically there for 2 weeks, were transformed into similar solutions but containing 0, 0.5, 2.5, 5, 10 and 25 mg L−1 Cd2+ (added in the form of CdCl2·2.5H2O), respectively. The experiment was repeated three times. After 2 and 4 weeks exposure, the plants were harvested for analysis.
Growth analysis
At the beginning and the end of the exposure, three plants were dried in an oven at 80 °C for 48 h and weighed. Plants used for dry mass measurement had not been used for other measurements, in order to avoid damage that might have affected the final biomass. The relative growth rate (RGR) of whole plants was calculated using the following formula (Susana et al. 2010): RGR = (ln W1 − ln W0) D−1 (mg g−1 day−1). W1 is the final dry mass, W0 is the initial dry mass (an average of the plants dried at the beginning of the experiment) and D is the duration of exposure (days).
Determination of chlorophyll and carotenoid contents
The extraction procedure was similar to Booker and Fiscus (2005) with small modification. Tissue leaf samples (0.2 g) were soaked in 25 ml 95 % (v/v) ethanol at 4 °C in darkness until the tissues became white. Extract was used to measure the absorbance at 649, 665 and 470 nm. Chlorophyll and carotenoid contents were calculated according to Lichtenthaler and Wellburn (1983).
Determination of relative water content (RWC)
Fully expanded third function leaf was sampled, and fresh weight (FW) was determined right after excision, and the leaves were immersed into double distilled water to saturate them in the dark for at least 18 h to determine the weight of leaves with saturated water (TW). Dry weight (DW) was weighed after drying the leaves to a constant weight in an oven at 80 °C. RWC was calculated using the following formula (Smart and Bingham 1974): RWC (%) = (FW − DW)/(TW − DW) × 100 %.
Determination of cell membrane permeability
Membrane permeability was estimated by measuring electrolyte leakage (EL). For electrolyte leakage measurement, twenty leaf discs (0.8 cm diameter) or 0.2 g root tips were cut off and rinsed with distilled water, put into 20 ml distilled water, and incubated at 25 °C for 6 h using shaker to measure the initial electric conductance of the water (S1). Then, the samples were heated in water bath in 80–90 °C for 30 min to kill the cells and determine the final electric conductance (S2). The electrolyte leakage was calculated as: EL (%) = S1/S2 × 100 %.
Determination of lipid peroxidation (MDA content) and antioxidant enzymes’ activities
Fresh tissue (0.2 g) was ground under liquid nitrogen and homogenized in 5.0 ml 50 mmol l−1 cold Na-phosphate buffer (pH 7.8), with 0.1 mM EDTA and 1 % (w/v) polyvinylpyrrolidone (PVP). After centrifuging at 13,000×g for 30 min at 4 °C, the supernatant was used for MDA and SOD assays.
The level of lipid peroxidation in the plant tissues was estimated by measuring the concentration of malondialdehyde (MDA), the major thiobarbituric acid (TBA)-reactive material, as described by Bowler et al. (1992). Absorbance of the supernatant was measured at 532 nm and corrected for nonspecific turbidity by subtracting the absorbance at 600 and 450 nm. The concentration of MDA was calculated as: MDA (μM) = 6.45(A532−A600)−0.56A450. SOD activity was determined according to the method of Krivosheeva et al. (1996). One enzyme unit of SOD activity was defined as the amount of enzyme required to cause 50 % inhibition in the rate of nitro-blue-tetrazolium (NBT) reduction at 560 nm. For APX activity assay, fresh tissues (0.2 g) were ground under liquid nitrogen and then homogenized in 3.5 ml of 50 mM 50 mmol l−1 cold Na-phosphate buffer (pH 7.6) containing 1 mM EDTA, 1 mM ascorbate, 2 % (w/v) PVP. Then 1.5 ml saturated (NH4)2SO4 in the same buffer was added and stirred well. After centrifugation at 4 °C and 13,000×g for 20 min, the supernatant was prepared for APX activity assay. APX activity was estimated by the rate of AsA oxidation, monitored by the change in absorbance at 290 nm (Krivosheeva et al. 1996).
Determination of Cd content in plant tissue
At the end of the experiment, the harvested leaves were rinsed with tap water, and the roots were immersed in 20 mM Na2-EDTA for 15 min to remove Cd adhered to the root surface, and then rinsed with distilled water. The leaf, stem and root samples were divided and dried at 80 °C for 48 h. Dried plant materials were weighed and ground to fine powder, and digested with a concentrated acid mixture of HNO3/HClO4 (3:1, v/v). Cd concentration was determined with atomic absorption spectrophotometer (Perkin-Elmer, USA).
The translocation factor (TF) indicated the ability of plants to translocate heavy metals from roots to shoots (Mattina et al. 2003). Translocation factor was related to cadmium concentration and calculated as Liu et al. (2009): TF = the cadmium concentration in shoots/the cadmium concentration in roots; TF was also related to cadmium accumulation (TF′) = the cadmium accumulation in shoots/the cadmium accumulation in roots.
The bioaccumulation coefficient (BC), or enrich factor, was described as Liu et al. (2009) and Tanhan et al. (2007): BC = the cadmium concentration in the whole plant/the cadmium concentration in the soil.
Statistical analyses
All measurements were replicated three times. Average values and standard deviations (SD) were calculated by the Microsoft Office Excel 2007 for all the data in this paper. One-way analysis of variance was carried out with SPSS13.0 for windows. LSD test was used to determine the difference (p < 0.05) among samples.
Results
Effects of Cd on plant growth
The effect of Cd on L. japonica growth varied with different Cd concentrations in the medium. After 28 days Cd exposure, the DW of plants was higher than that of control plants when exposure to the lower Cd concentrations from 0.5 to 5 mg L−1, but showed significantly decreased at 25 mg L−1 Cd (Table 1). The RGR of the whole plant, roots, stems and leaves were all increased in comparison with the control at Cd concentration from 0.5 to 5 mg L−1, and then turned to decrease from 10 to 25 mg L−1, displaying an inverted U-shaped dose–response curve, which confirmed that the positive effect of low Cd levels occurred (Fig. 1). The fastest growth rate of roots and stems occurred at 0.5 and 2.5 mg L−1 Cd treatment, respectively. The leaves lagged in response to Cd and the fastest growth rate occurred at 5 mg L−1 Cd treatment, but had no significant difference compared with control. Low Cd-concentrations (≤5 mg L−1) markedly stimulated growth of the plants, so that the whole plant biomass was increased by 6.62, 14.74 and 7.83 % compared with the control, respectively.

Effects of Cd concentrations in culture medium on relative growth rate of the whole plant, roots, stems and leaves of L. japonica after 28 days exposure. Values represent mean ± SD, n = 3. Different letters indicate significant difference at the 5 % level according to the LSD test
Effects of Cd on chlorophyll and carotenoid contents
Table 2 shows that the total chlorophyll, chlorophylla and carotenoid contents were significantly increased by 0.5 and 2.5 mg L−1 Cd, chlorophyll b content was significantly increased by 5 mg L−1 Cd, but all of the photosynthetic pigments contents dramatically decreased by exposure to higher concentrations Cd, which indicated that low dosage of Cd may be beneficial to plants but high dosage produced toxic effects. The maximum increase of total chlorophyll, chlorophylla, chlorophyllb and carotenoid content was 9.33, 12.95, 10.00 and 45.24 % of the control, respectively. Among all the Cd treatments, the carotenoid content increased the most obviously and only decreased by the highest Cd concentration tested (25 mg L−1). The highest chlorophyll and carotenoid content occurred at 2.5 mg L−1Cd treatment. This was consistent with the whole plant DW.
Effects of Cd on the RWC of leaves
After 28 days Cd exposure, the RWC of leaves was increased by the low concentrations of Cd (<2.5 mg L−1) (Fig. 2). Significant increase was observed at 0.5 mg L−1, the highest increase was 3.04 % higher than the control. When Cd concentration in the medium was higher than 2.5 mg L−1, the RWC of leaves decreased with increasing Cd concentrations. A significant (6.85 %) decrease was detected at 25 mg L−1 Cd, compared to the control. An inverted U-shaped dose–response curve was observed in this feature.

Effects of Cd concentrations in medium on RWC for leaves of L. japonica after 28 days exposure. Values represent mean ± SD, n = 3. Different letters indicate significant difference at the 5 % level according to the LSD test
Effects of Cd on MDA content and electrolyte leakage
The MDA content in leaves declined with the increase of Cd from 0 to 2.5 mg L−1 after 14 days exposure and then enhanced, indicating a U-shaped curve of MDA levels along with the increase of Cd in medium (Fig. 3a). With the prolongation of exposure time to 28 days, both in leaves and roots the MDA content were not significantly affected by low Cd concentration (<5 mg L−1), but increased dramatically and reached their maximum, 168.49 % for roots at 10 mg L−1 and 143.52 % for leaves at 25 mg L−1 Cd, compared to the control (Fig. 3a, b). The extent of cell membrane damage estimated on the basis of electrolyte leakage of leaves and roots was similar to the changes in MDA content. The ion leakage in leaves was slightly declined at 0.5 and 2.5 mg L−1 Cd after 14 days exposure, but no significant difference compared with the control (Fig. 4a). After 28 days exposure, both in leaves and roots the ion leakage were not significantly affected by low Cd concentration (<5 mg L−1), but increased along with the increase of Cd in medium, which suggested that low concentrations of Cd (<5 mg L−1) did not cause lipid peroxidation and membrane damage in L. japonica (Fig. 4a, b).

Effects of Cd concentrations in medium on malondialdehyde (MDA) content in leaves (a) and roots (b) of L. japonica after 14 and 28 days exposure. Values represent mean ± SD, n = 3. Different letters indicate significant difference at the 5 % level according to the LSD test

Effects of Cd concentrations in medium on electrolyte leakage in leaves (a) and roots (b) of L. japonica after 14 and 28 days exposure. Values represent mean ± SD, n = 3. Different letters indicate significant difference at the 5 % level according to the LSD test
Effects of Cd on the activities of antioxidant enzymes
In our study, increased SOD and APX activity in leaves and roots were observed along with the increased Cd concentration in the medium (Figs. 5 and 6). In roots, SOD activity increased with the increase in Cd concentration till 10 mg L−1 and then sharply decrease when at the highest tested Cd concentration after 28 days exposure (Fig. 5b), which might be related to the cell damage as indicated by high ion leakage. The APX activity in leaves was slightly declined at low Cd concentrations below 10 mg L−1, but increased about 1.6-fold of the control at 25 mg L−1 Cd after 28 days exposure (Fig. 6a). In roots, APX activity sharply increased at concentrations from 0.5 to 10 mg L−1 Cd, then decreased at higher concentrations at both 14 and 28 days exposure (Fig. 6b). The SOD and APX activities in roots seemed to respond to Cd more rapidly than that in leaves, which might be explained by the fact that roots is the first organ coming in direct contact with Cd and more Cd was accumulated in roots rather than in leaves.

Effects of Cd concentrations in medium on the activity of superoxide dismutase (SOD) in leaves (a) and roots (b) of L. japonica after 14 and 28 days exposure. Values represent mean ± SD, n = 3. Different letters indicate significant difference at the 5 % level according to the LSD test

Effects of Cd concentrations in medium on the activity of ascorbate peroxidase (APX) in leaves (a) and roots (b) of L. japonica after 14 and 28 days exposure. Values represent mean ± SD, n = 3. Different letters indicate significant difference at the 5 % level according to the LSD test
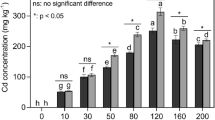
Cd accumulation in L. japonica
After 28 days Cd-exposure, Cd concentrations in leaves, stems and roots of L. japonica all increased significantly with increasing Cd concentrations in the medium (Table 3). The highest Cd concentrations in leaves, stems and roots were 128.52, 603.97 and 1,575.64 mg kg−1 DW, respectively, when L. japonica was exposed to 25 mg L−1 Cd in the medium. The BC of L. japonica exposed to various Cd concentrations were always much higher than 1, but the value decreased along with the increase Cd concentration in the medium. Although the Cd concentrations in roots were much higher than that in aerial parts and the TFs were between 0.36 and 0.72, the TF′s were always higher than 1 and the TF′s were between 3.04 and 9.47. The highest TF and TF′ were found at a low (2.5 mg L−1), Cd concentration and then decreased with the increase of Cd concentrations in the medium.
Discussion
Hormesis is any stimulatory effect induced by low doses of toxicant that cannot be predicted by the extrapolation of detrimental or lethal effects induced by high doses of the same toxicant (Calabrese and Baldwin 2003a, b). The present study demonstrated clearly that Cd induced significant increase in plant growth (Fig. 1; Table 1), leaf water content (Fig. 2) and photosynthetic pigments (Table 2) in L. japonica at lower concentrations (<5 mg L−1) but inhibited at higher concentrations, which coincided with the definition of hormesis in toxicology. Multiple hormetic effects in L. japonica induced by Cd were investigated in this present study. The U-shaped dose–response curve of MDA content in leaves was observed after 14 days exposure indicated that reduced oxidative stress occurred at suitable spacing of Cd doses. Regarding MDA (Fig. 3) content and cell membrane permeability (Fig. 4) both in roots and leaves of L. japonica with the prolongation of exposure time to 28 days, the results indicated that severe oxidative damage occurred only at the highest concentration but not at low concentrations Cd.
Cadmium is most commonly found metal in soil, as we all know that it is a non-essential and toxic element for plant. Elevating Cd level usually leads to growth inhibition as this metal tend to accumulate within plant tissues and negatively interfere with essential physiological processes (di Toppi and Gabbrielli 1999). In this research, Cd enhanced L. japonica growth up to 5 mg L−1 and significantly decreased it at 25 mg L−1. Relative growth rates showed that Cd stimulated growth of all plant parts at different extent. The RGR of roots only stimulated by 0.5 and 2.5 mg L−1 Cd, and dramatically decreased by 10 and 25 mg L−1 Cd, indicating that the roots more susceptible to Cd than aerial parts. At the end of exposure, the maximum increase in DW of roots, stems and leaves were 46.09 %, 17.44 and 9.98 % compared with the control, respectively when the plants was exposed to 0.5, 2.5 and 5 mg L−1 Cd. With further increase of Cd concentration in the medium, the growth decreased. The maximum reductions in DWs of roots, stems and leaves were 32.59, 6.22 and 21.70 %, respectively when the plants was exposed to 25 mg L−1 Cd (Table 1). Similar biphasic dose responses were also found in RWC (Fig. 2), chlorophylls and carotenoids contents of leaves (Table 2) in L. japonica within this study. The results confirmed our previous finding that there is some potentially positive effect of lower concentration Cd on plant growth (Liu et al. 2009). Chlorophyll is vital for photosynthesis and carotenoids play an important role in protecting the photosynthetic apparatus against various harmful environmental factors (Strzałka et al. 2003). In this study, low Cd concentration resulted in an increase of chlorophyll and carotenoids contents in L. japonica, demonstrating that low dosage of Cd induced some beneficial effect on photosynthetic system, which might explains the increase in plant growth rate. The same phenomenon has been found in Sedum alfredii H. under Cd stress by Zhou and Qiu (2005), and the main explanation of the increase of chlorophyll contents could be a result of Cd-induced increase of Fe uptake. Our previous studies also revealed that there is a synergistic interaction in accumulation and translocation between Cd and Fe in L. japonica, and the uptake of Zn and Mn in leaves has been improved at low concentrations of Cd exposure (Liu et al. 2011). However, the interactions between Cd and nutrient trace elements, and the linkage between Cd treatment and the increase of chlorophyll content need for further investigated.
Some physiological responses related with the hormetic effect were also investigated in this present study. The accumulation of malondialdehyde (MDA) is the most widely accepted indicator of potential oxidative damage, which is a breakdown product of lipid peroxidation (Smirnoff 1993). The plant cell membranes are also generally considered as primary sites of metal injury (Belkhadi et al. 2010). In the present study, the levels of MDA and ion leakage in leaves declined below their respective controls at 0.5–2.5 mg L−1 Cd after 14 days Cd exposure, indicating reduced oxidative stress and enhanced beneficial effect in leaves. Similar results were reported in roots of Vicia faba seedlings under Pb stress (Wang et al. 2010). At the end of the experiment, the MDA content and leakage of ions both in leaves and roots were only increased upon addition of Cd at higher concentrations, demonstrating that the structure and functions of cell membranes did not affected by low levels of Cd stress (Figs. 3, 4). The data of MDA content combined with ion leakage indicated that the capabilities of the plant to adapt to lower concentration of Cd might be related to a low degree of lipid peroxidation and thus an improved growth was maintained.
Cadmium toxicity can disturb the balance between the ROS generation and removal, and cause MDA accumulation in plants which results in oxidative damage (Scebba et al. 2006; Lin et al. 2007; Shan et al. 2012). As a defensive mechanism, antioxidative enzymes, such as SOD and APX, play an important role in scavenging ROS protecting cells in higher plants (Gill and Tuteja 2010). SOD dismutates O2 − into O2 and H2O2 and H2O2 then is reduced by POD, APX and others. After 4 weeks, the MDA and ion leakage did not affected by low Cd concentration (<5 mg L−1), whereas the SOD and APX activities in roots increased above their respective controls, implying enhanced oxidative stress and adverse effect. In other words, enhanced defense response was induced by low doses of Cd, indicating that the antioxidant enzyme defense system might be dominate at low Cd concentrations that ensures redox homeostasis is maintained. Biological systems adapt for Cd oxidative stress. Nonetheless, the overall net positive effect was observed at low concentrations of Cd in medium. This is consistent with the findings of Scebba et al. (2006) who found a positive effect of Cd on plant growth at low Cd concentrations. Parson (2003) reported that the induction of defense systems that can ameliorate the effect of toxic chemicals in an organism has also been proposed to be a cause of hormetic responses. This hypothesis was also supported by Kovalchuk et al. (2003), who proposed that the defense mechanisms induced by oxygen free radicals can lead to increased growth in the presence of low doses of phytotoxic chemicals. Toxicants could induce beneficial homeostatic responses at suitable spacing of doses and are responsible for enhanced defense ability to repair damaged cells in organisms (Calabrese and Baldwin 2003a, b). But the reason for the hormesis phenomenon is not completely understood, which also might be regulated by multiple mechanisms such as metabolism homeostatic regulation (i.e. allosteric regulation of enzyme activities, translation of accumulated mRNAs, protein turnover) and the trigger of specific genetic programs with new protein synthesis. The underlying mechanism of hormesis needs to be further investigated.
In the past decade, there has been considerable interest in the area of soil remediation of contaminated sites. Phytoremediation has become a promising soil remediation technique where hyper-accumulators or accumulators are used to take up large quantities of pollutant metals (Salt et al. 1998; Roosens et al. 2003). Numerous studies in the area have reported hormetic responses such that low levels of site contamination have been associated with enhanced plant growth and enhanced accumulation of Cd, by the plant species employed (Calabrese and Blain 2009). In the present study, after 28 days exposure to 10 mg L−1 Cd, Cd concentrations got up to 502.96 ± 28.90 mg kg−1 DW in stem and 103.22 ± 5.62 mg kg−1 DW in leaf (Table 2), more than 100 mg kg−1 dry tissue, which is the threshold value of Cd-hyperaccumulator (Baker and Brooks 1989; Sun et al. 2008). Especially, the plant did not show any significant damage, and the leaves, stems and total biomass had no significant differences compared with the control when exposed to 10 mg L−1. Phytoremediation efficiency depends on plant biomass and the ability of metal to be translocated to the shoots (Wu et al. 2010). Thus, transport factor (TF′) could be better parameter in indicating metal transport efficiency than TF. TF′ values of L. japonica were always much higher than 1 at all the tested Cd concentrations, which indicated that L. japonica could transport large amount Cd from root to aerial structures. Based on higher BC and TF′ values, the high Cd accumulation in the stems and leaves indicated that L. japonica has the potential as a hyperaccumulator in phytoremediation application for Cd-contaminated soils. This is further confirmed our previous study (Liu et al. 2009; 2011). Additionally, the highest TF and TF′ values of L. japonica were 0.72 and 9.47 at 2.5 mg L−1 Cd, and then turn to decline with the increase in Cd concentration in the nutrient solution, implying that low Cd (2.5 mg L−1) improved the ability to transfer Cd from roots to aerial structures, which might be related with the mechanisms of Cd accumulation and translocation in L. japonica. The same phenomenon has been found in Cd-accumulators Bidens pilosa L. (Sun et al. 2009). But the underlying mechanisms needs to be further investigated. The hormetic phenomenon induced by heavy metal in plant was also observed in many other studies, especially in some hyperaccumulators, such as Brassica juncea L. (Seth et al. 2008) and Arabis paniculata F. (Qiu et al. 2008; Tang et al. 2009). Because hormesis is ubiquitous, as a potential Cd-hyperaccumulator, L. japonica may have a wider range of low-dose stimulation effect than many other plants. Taken together, the present study suggested that hormetic effects should be taken into consideration in phytoremediation of Cd-contaminated soil.
Conclusion
In our study, all the evidence points to the fact that plant growth, leaf water content and photosynthetic pigments in L. japonica were increased by Cd at low concentrations but inhibited at high concentrations, which indicates the occurrence of a hormetic phenomenon. The low level of MDA content and membrane permeability observed in the experiment indicating reduced oxidative stress and enhanced beneficial effect in the plants. Rapidly enhanced defense response in roots was induced by low doses of Cd, implying that the antioxidant enzyme defense system might be dominate at low Cd concentrations that ensures redox homeostasis is maintained. The presence of hormesis induced by Cd in L. japonica may be related with such biological mechanisms as oxidative stress level. However, the hormetic responses may occur as a result of numerous mechanisms, depending on the cell type. Further work is needed, L. japonica could be considered as a new plant to investigate the underlying mechanisms of the hormesis. On the other hand, L. japonica had a potential ability as phytoremediator of Cd-contaminated soils, and moreover, low Cd treatments improved the ability to transfer Cd from roots to aerial structures, therefore, our results suggest that hormetic effects should be taken into consideration in phytoremediation of Cd-contaminated soil.
References
Baker AJM, Brooks RR (1989) Terrestrial higher plants which hyperaccumulate metallic elements—a review of their distribution, ecology and phytochemistry. Biorecovery 1:81–126
Baryla A, Carrier P, Franck F, Coulomb C, Sahut C, Havaux M (2001) Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cadmium-polluted soil: causes and consequences for photosynthesis and growth. Planta 212:696–709
Belkhadi A, Hediji H, Abbes Z, Nouairi I, Barhoumi Z, Zarrouk M, Chaïbi W, Djebal W (2010) Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content in Linum usitatissimum L. Ecotox Environ Saf 73:1004–1011
Booker FL, Fiscus EL (2005) The role of ozone flux and antioxidants in the suppression of ozone injury by elevated CO2 in soybean. J Exp Bot 56:2139–2151
Bowler C, Montagu MV, Inzé D (1992) Superoxide dismutase and stress tolerance. Plant Physiol Plant Mol Biol 43:83–116
Calabrese EJ, Baldwin LA (1999) Chemical hormesis: its historical foundations as biological hypothesis. Toxicol Pathol 27:195–216
Calabrese EJ, Baldwin LA (2001) The frequency of U-shaped dose-responses in the toxicological literature. Toxicol Sci 62:330–338
Calabrese EJ, Baldwin LA (2003a) Toxicology rethinks its central belief: hormesis demands a reappraisal of the way risks are assessed. Nature 421:691–692
Calabrese EJ, Baldwin LA (2003b) The hormetic dose-response model is more common than the threshold model in toxicology. Toxicol Sci 71:246–250
Calabrese EJ, Blain RB (2009) Hormesis and plant biology. Environ Pollut 157:42–48
Calabrese EJ, Staudenmayer JW, Stanek EJ, Hoffmann GR (2006) Hormesis outperforms threshold model in NCI anti-tumor drug screening data. Toxicol Sci 94:368–378
Calabrese EJ, Stanek EJ, Nascarella MA, Hoffmann GR (2008) Hormesis predicts low-dose responses better than threshold models. Int J Toxicol 27:369–378
Calabrese EJ, Hoffmann GR, Stanek EJ, Nascarella MA (2010) Hormesis in high-throughput screening of antibacterial compounds in E coli. Hum Exp Toxicol 29:667–677
Chen X, Wang J, Shi Y, Zhao MQ, Chi GY (2011) Effects of cadmium on growth and photosynthetic activities in pakchoi and mustard. Bot Stud 52:41–46
de la Rosa G, Peralta-Videa JR, Montes M, Parsons JG, Cano-Aguilera I, Gardea-Torresdey JL (2004) Cadmium uptake and translocation in tumbleweed (Salsolakali), a potential Cd-hyperaccumulator desert plant species: ICP/OES and XAS studies. Chemosphere 55:1159–1168
di Toppi LS, Gabbrielli R (1999) Response to cadmium in higher plants. Environ Exp Bot 41:105–130
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. Calif Agric Exp Stn Circ 347:1–32
Jia L, Liu ZL, Chen W, He XY (2012) Stimulative effect induced by low-concentration cadmium in Lonicera japonica Thunb. Afr J Microbiol Res 6:826–833
Kovalchuk I, Filkowski J, Smith K, Kovalchuk O (2003) Reactive oxygen species stimulate homologous recombination in plants. Plant Cell Environ 26:1531–1539
Krivosheeva A, Tao DL, Ottander C, Wingsle G, Dube SL, Oquist G (1996) Cold acclimation and photoinhibition of photosynthesis in Scots pine. Planta 200:296–305
Larson BMH, Catling PM, Waldron GE (2007) The biology of Canadian weeds.135. Lonicera japonica Thunb. Can J Plant Sci 87:423–438
Lichtenthaler HK, Wellburn AR (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans 11:591–592
Lin RZ, Wang XR, Luo Y, Du WC, Guo HY, Yin DQ (2007) Effects of soil cadmium on growth, oxidative stress and antioxidant system in wheat seedlings (Triticum aestivum L.). Chemosphere 69:89–98
Liu ZL, He XY, Chen W, Yuan FH, Yan K, Tao DL (2009) Accumulation and tolerance characteristics of cadmium in a potential hyperaccumulator–Lonicera japonica Thunb. J Hazard Mater 169:170–175
Liu ZL, He XY, Chen W (2011) Effects of cadmium hyperaccumulation on the concentrations of four trace elements in Lonicera japonica Thunb. Ecotoxicology 20:698–705
Mattina MJI, Lannucci-Berger W, Musante C, White JC (2003) Concurrent plant uptake of heavy metals and persistent organic pollutants from soil. Environ Pollut 124:375–378
Mattson MP, Calabrese EJ (2010) Hormesis: what it is and why it matters. Hormesis. Springer, New York, pp 1–13
Parson PA (2003) Metabolic efficiency in response to environmental agents predicts hormesis and invalidates the linear no-threshold premise: ionizing radiation as a case study. Crit Rev Toxicol 33:443–450
Poschenrieder C, Gense B, Barcelo J (1989) Influence of cadmium on water relations, stomatal resistance and abscisic acid content in expanding bean leaves. Plant Physiol 90:1365–1371
Qiu RL, Zhao X, Tang YT, Yu FM, Hu PJ (2008) Antioxidative response to Cd in a newly discovered cadmium hyperaccumulator, Arabis paniculata F. Chemosphere 74:6–12
Roosens N, Verbruggen N, Meerts P, Ximenez-Embun P, Smith J (2003) Natural variation in cadmium tolerance and its relationship to metal hyperaccumulation for seven populations of Thlaspi caerulescens from Western Europe. Plant Cell Environ 10:1657–1672
Salt DE, Smith RD, Raskin I (1998) Phytoremediation. Annu Rev Plant Physiol Plant Mol Biol 49:643–668
Scebba F, Arduini I, Ercoli L, Sebastiani L (2006) Cadmium effects on growth and antioxidant enzymes activities in Miscanthus sinensis. Biol Plantarum 50:688–692
Seth CS, Chaturvedi PK, Misra V (2008) The role of phytochelatins and antioxidants in tolerance to Cd accumulation in Brassica juncea L. Ecotoxicol Environ Saf 71:76–85
Shah K, Kumar RG, Verma S, Dubey RS (2001) Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedling. Plant Sci 161:1135–1144
Shan SH, Liu F, Li CJ, Wan SB (2012) Effects of cadmium on growth, oxidative stress and antioxidant enzyme activities in peanut (Arachis hypogaea L.) seedlings. J Agric Sci 4:142–151
Skórzyńska-Polit E, Drażkiewicz M, Krupa Z (2010) Lipid peroxidation and antioxidative response in Arabidopsis thaliana exposed to cadmium and copper. Acta Physiol Plant 32:169–175
Smart RE, Bingham GE (1974) Rapid estimates of relative water content. Plant Physiol 53:258–260
Smirnoff N (1993) The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol 125:27–58
Stebbing ARD (1998) A theory for growth hormesis. Mutat Res 403:249–258
Strzałka K, Kostecka-Gugała A, Latowski D (2003) Carotenoids and environmental stress in plants: significance of carotenoid-mediated modulation of membrane physical properties. Russ J Plant Physiol 50:168–172
Sun YB, Zhou QX, Diao CY (2008) Effects of cadmium and arsenic on growth and metal accumulation of Cd-hyperaccumulator Solanum nigrum L. Bioresour Technol 99:1103–1110
Sun YB, Zhou QX, Wang L, Liu W (2009) Cadmium tolerance and accumulation characteristics of Bidens pilosa L. as a potential Cd-hyperaccumulator. J Hazard Mater 161:808–814
Susana RZ, Enrique MN, Luis AM (2010) Accumulation and tolerance characteristics of cadmium in a halophytic Cd-hyperaccumulator, Arthrocnemum macrostachyum. J Hazard Mater 184:299–307
Tang YT, Qiu RL, Zeng XW, Ying RR, Yu FM, Zhou XY (2009) Lead, zinc, cadmium hyperaccumulation and growth stimulation in Arabis paniculata Franch. Environ Exp Bot 66:126–134
Tanhan MKP, Pokethitiyook P, Chaiyarat R (2007) Uptake and accumulation of cadmium lead and zinc by Siam weed [Chromolaena odorata (L.) King & Robinson]. Chemosphere 68:323–329
Vig K, Megharaj M, Sethunathan N, Naidu R (2003) Bioavailability and toxicity of cadmium to microorganisms and their activities in soil: a review. Adv Environ Res 8:121–135
Wang CR, Tian Y, Wang XR, Yu HX, Lu XW, Wang C (2010) Hormesis effects and implicative application in assessment of lead-contaminated soils in roots of Vicia faba seedlings. Chemosphere 80:965–971
Wu SC, Cheung KC, Luo YM, Wong MH (2006) Effects of inoculation of plant growth-promoting rhizobacteria on metal uptake by Brassica juncea. Environ Pollut 140:124–135
Wu F, Yang W, Zhang J, Zhou L (2010) Cadmium accumulation and growth responses of a poplar (Populus deltoids×Populus nigra) in cadmium contaminated purple soil and alluvial soil. J Hazard Mater 177:268–273
Zhou WB, Qiu BS (2005) Effects of cadmium hyper-accumulation on physiological characteristics of Sedum alfredii Hance (Crassulaceae). Plant Sci 169:737–745
Acknowledgments
The authors are very thankful to Dr. Dali Tao, for his help in improving the manuscript. This work was funded by the National Science and Technology Pillar Program (2012BAC05B05) and the major National Science and Technology project “Water Pollution Control and Management” (2012ZX07202008) of China.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jia, L., He, X., Chen, W. et al. Hormesis phenomena under Cd stress in a hyperaccumulator—Lonicera japonica Thunb. Ecotoxicology 22, 476–485 (2013). https://doi.org/10.1007/s10646-013-1041-5
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-013-1041-5