
Abstract
The objectives of this study were: (1) to examine the distribution and bioaccumulation of microcystins in the main components of the food web (phytoplankton, zooplankton, crayfish, shrimp, mussel, snail, fish, frog) of Lake Pamvotis (NW Greece), (2) to investigate the possibility of microcystin biomagnification and (3) to evaluate the potential threat of the contaminated aquatic organisms to human health. Significant microcystin concentrations were detected in all the aquatic organisms during two different periods, with the higher concentrations observed in phytoplankton and the lower in fish species and frogs. This is the first study reporting microcystin accumulation in the body of the freshwater shrimp Atyaephyra desmsaresti, in the brain of the fish species common carp (Cyprinus carpio) and in the skin of the frog Rana epirotica. Although there was no evidence for microcystin biomagnification, the fact that microcystins were found in lake water and in the tissues of aquatic organisms, suggests that serious risks to animal and public health are possible to occur. In addition, it is likely to be unsafe to consume aquatic species harvested in Lake Pamvotis due to the high-concentrations of accumulated microcystins.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Algal blooms are increasing worldwide due to eutrophication of aquatic environments (Anderson et al. 2002). Anthropogenic nutrient enrichment of freshwaters has a direct impact on algal species composition and the formation of noxious and toxic blooms and surface scums. The most common phytoplankton organisms associated with eutrophication of freshwater systems are cyanobacteria (Codd 1995). Cyanobacteria have the ability to form several toxic metabolites (Ressom et al. 1994) known as cyanotoxins. Among them, microcystins are the most abundant group. Microcystins are produced by various genera of cyanobacteria, the predominant one being Microcystis sp. (Carmichael 1997; Codd 2000; Beresovsky et al. 2006). These toxins are highly water-stable and resistant to boiling, and thus pose a threat to water and food quality if not properly monitored (Falconer and Humpage 2005).
Exposure to microcystins represents a health risk to aquatic organisms, wild life, domestic animals, and humans upon drinking or ingesting cyanobacteria in the water (Duy et al. 2000). It is well recognized that among their toxigenic mechanisms they are potent inhibitors of protein phosphatase 1 and 2A that cause increased protein phosphorylation, which is directly related to their cytotoxic effects and tumor-promoting activity (Hooser et al. 1989; Carmichael 1994; Hooser 2000). Chronic toxic effects to humans from exposure through drinking water or contaminated food are particularly probable, especially if there is long-term frequent exposure (Magalhães et al. 2001; Zhang et al. 2009). High incidence of primary liver cancer in China is considered to be related to microcystins in drinking water (Yu 1995).
Bioaccumulation of microcystins in aquatic food chains may be another important factor, suggesting a potential risk for higher trophic level species. The ability of zooplankton and other invertebrates to accumulate microcystins may lead to a transfer of toxins through the food web to more sensitive organisms (Liras et al. 1998). Bivalves, as sessile filter-feeding animals, may be among the most threatened group of aquatic organisms in the presence of toxic cyanobacterial blooms. Gastropods are important links between primary producers and higher consumers (Habdija et al. 1995), thus toxin accumulation by these organisms, increases the risk for higher consumers. In the same way, toxin accumulation by amphibians increases the risk for terrestrial animals. The occurrence of toxic cyanobacteria in many water bodies where macroinvertebrates and large crustacea are abundant leads to the possibility of toxin accumulation and transfer through the food chain. Moreover, people consume a lot of freshwater fish, thus large quantities of cyanotoxins could also be transferred by this way.
However, only a few studies have tracked the presence of microcystins in single or multiple components of aquatic food webs (Kotak et al. 1996; Babcock-Jackson et al. 2002; Magalhães et al. 2001, 2003; Ibelings et al. 2005). Other studies have analyzed the uptake of microcystins by aquatic organisms in an experimental setting (among others, zooplankton (Demott 1999; Rohrlack et al. 2001; Thostrup and Christoffersen 1999), mussels (Amorim and Vasconcelos 1999; Prepas et al. 1997; Yokoyama and Park 2003), or fish (Malbrouck et al. 2003), often using an acutely toxic dosage and purified microcystin or cultured cyanobacteria (Xie et al. 2004). These results, therefore, are not readily transferable to a lake food web.
Many Greek lakes suffer from the presence of cyanobacterial blooms (Vardaka et al. 2005). Moreover, significant microcystin concentrations have been recorded in Greek freshwaters, including Lake Pamvotis (Kagalou et al. 2008; Papadimitriou et al. 2010). Our study was triggered by the high microcystin concentrations recorded in the tissues of an omnivorous fish species (Carassius gibelio), originated from Lake Pamvotis during the last years (Kagalou et al. 2008; Papadimitriou et al. 2010). This led us to consider about the dynamics of microcystin concentrations in significant aquatic food web components of Lake Pamvotis.
The major objectives of this study were: (1) to examine the distribution and bioaccumulation of microcystins in some aquatic food web compartments (phytoplankton, zooplankton, crayfish, shrimp, mussel, snail, fish, frog, (2) to investigate the possibility of microcystin biomagnification, (3) to evaluate the potential risk of the contaminated aquatic organisms to human health.
Materials and methods
Sample collection and preparation
Study area
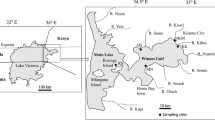
The study was conducted in Lake Pamvotis, NW Greece (39°40′N, 20°53′E) which is a Mediterranean, warm shallow lake (mean depth, 5.5 m). In lake water, many activities take place, like recreational activities, fishing and water uptake for irrigation (Kagalou et al. 2003; Papadimitriou et al. 2010). During the last three decades the trophic status of the lake has changed as a result of anthropogenic activity (among others irrigation and domestic sewage discharge), resulting in serious problems (Kotti et al. 2000; Kagalou et al. 2008). Previews studies, have proven high cyanobacterial biovolumes in the lake (Vardaka et al. 2005). Dense scum of toxic cyanobacterial blooms occurs frequently in the water surface of the lake, mainly in the warm seasons every year (Papadimitriou et al. 2011).
Sampling of phytoplankton and collection of cyanobacterial blooms
Water samples (volume of 2 l) for phytoplankton and microcystins analysis were collected monthly from the surface of the water to a depth of 1 m using polyethylene bottles, during 2008. Subsamples of 50 mL were preserved with both Lugol’s solution and formaldehyde. Phytoplankton identification was performed according Anagnostidis and Economou (1980), using an inverted microscope.
To estimate microcystin concentrations in the lake water (intracellular and extracellular toxins) 1 L of the water sample was filtered through a Whatman glass-fiber filter (Whatmann GF/C 0.45-μm filter). Seston onto the glass-fiber filter and the filtrate were used to detect intracellular and extracellular toxins, respectively.
Cyanobacterial blooms were collected from the surface water with a plankton net (mesh size, 63 μm) during August and November 2008, and immediately frozen at −20°C and then lyophilized for toxin analysis.
Sampling of zooplankton
Zooplankton from which the microcystin content was analyzed was collected from 0.5 m depth, by filtration of 500 L of lake water over a plankton net with a 250 μm mesh size. Collection was performed during August and November of 2008. A subsample was resuspended in 45 mL tap water and preserved with 5% formaldehyde for zooplankton identification. This was done under a microscope based on descriptions of Pennak (1978). The remaining volume of the sample was resuspended in 2 L tap water for microcystin analysis. Before the analytical procedure could be started, samples had to be prepared to avoid contamination with phytoplankton, which would probably contain microcystin. According to Ibelings et al. (2005), the following steps were followed: (1) the water was left standing overnight in measuring cylinders at in situ lake temperature and under dimmed light, allowing detritus to sink to the bottom and buoyant cyanobacteria to float to the top; (2) scums of cyanobacteria were removed, and the remainder of the sample (but excluding the detritus) was rinsed carefully with tapwater and transferred to clean beakers; (3) the water containing the zooplankton was flushed with CO2, by which treatment the zooplankton was sedated and sank to the bottom of the beakers—the overlying water was removed; (4) these steps were repeated until zooplankton was free of cyanobacteria (checked microscopically).
Sampling of macroinvertebrates
During August and November of 2008 (after November, the collection of aquatic organisms is difficult due to the ability of many aquatic organisms to reduce their metabolic activities), a suite of aquatic macroinvertebrates (10 specimens from every species) was collected and analyzed for microcystins. These included water snails (Viviparus contectus), freshwater shrimps (Atyaephyra desmaresti), crayfish (Astacus astacus) and mussels (Anodonta cygnea). These invertebrates were collected with a dip net from the shoreline or from the littoral zone from the side of the boat. In the cases of mussels and crayfish each individual was weighted sacrificed and the hepatopancreas, stomach, mantle, foot and gills (for mussels) and gills, hepatopancreas, brain, muscle, stomach and gonads (for crayfish) were collected, weighted and immediately frozen at −80°C. While for shrimps and water snails (without of shell) the whole body individually was weighted and frozen at −80°C.
Sampling of fish and frogs
Ten specimens of the fish species common carp (Cyprinus caprio) and the endemic Epirus frog (Rana epirotica) were captured in the same as the invertebrate’s period. The collected fish and frogs were measured, weighed (mean weight: 849 g for fish and 29.3 g for frogs), and sacrificed immediately. Liver, intestines, kidney, gonads, brain and muscle (for fish) and intestine, liver, pancreas, gonads, skin and muscle (for frogs) were collected, weighed and immediately frozen at −80°C.
In addition, six specimens of common carp (Cyprinus carpio) originated from artificial propagation, which were kept for 16 months in aquariums with pure recycled water used as control sample.
Microcystin analysis
Microcystins in the seston (intracellular microcystins) of water column and zooplankton samples were analyzed quantitatively: microcystins in the seston and in zooplankton samples extracted with 100% methanol and stirring overnight at room temperature (20–22 C) followed by centrifugation at 1,300×g for 15 min. Extraction procedure was repeated three times and the supernatants of the extractions were pooled. The organic solvent was removed by placing the extract under nitrogen-stream. The concentrated sample extract remaining after removing of the organic solvent was subjected to ELISA assay (ABRAXIS, 520011, USA).
For analysis of microcystins dissolved in water, the filtered water was applied directly to ELISA.
For analysis of microcystins in bloom samples, lyophilized material was extracted with 100% methanol and sonicated for 10 min in ice. After centrifugation at 4,000×g rpm (20 min), the supernatant was retained and filtered through a 0.2 μm filter to exclude cell derbis. Microcystin detection and quantification in bloom extracts were done according to Lawton et al. (1994), using a Shimadzu HPLC system (Duisburg, Germany) equipped with a photodiode array detector (SPD-MZOA) and a CTO 10AS column oven. The column was a ThermoHypersil (ODS) C18 (250 mm × 4.6 mm, 5 μm). The mobile phase system was: (A): aqueous trifluoracetic (TFA) 0.05% (v/v) and (B): acetonitrile (TFA) 0.05% (v/v). A gradient elution program was employed as follows: 50% A isocratically for 10 min, to 40% for 5 min, to 10% A for 5 min and finally to 50% A for 10 min. The HPLC run program time was 30 min. The sample volume injected was 50 μL and the mobile phase run at 0.6 mL min−1. Microcystins were identified on basis of a characteristic spectrum between 237 and 242 nm. The number of different microcystins was determined by the number of peaks that eluted with different retention times and showed a typical microcystin UV spectrum. Hence microcystins could be differentiated but not identified on basis of HPLC retention times. All different microcystins were transformed to microcystin-LR (MC-LR) equivalents for quantification of the total microcystin content. MC-LR (ALEXIS BIOCHEMICALS) was the only standard that was used.
Microcystins were extracted from the tissues of macroinvertebrates, fish and frogs, with 100% methanol (Magalhães et al. 2001), stirred overnight at room temperature and then centrifuged at 1,300×g for 15 min in a Universal 32 centrifuge. The supernatant was collected and stored overnight at 4°C. A 5 mL aliquot of the supernatant was concentrated to 350 μL under nitrogen stream. A 100-μL of the concentrated sample extract was diluted to 900 μL distilled water according to Sipia et al. (2002). The final sample was clarified using membrane filters (pore size, 0.45 μm, and diameter 4 mm). Sample solutions were analyzed immediately with ELISA test.
For each ELISA test, the negative control and four calibrates were assayed at least in duplicate. A commercial ABRAXIS-Microcystin ELISA kit (520011, USA) was used following the instructions of the manufacturer. The ELISA is an indirect-competitive method used for the quantitative analysis of all the microcystin analogues and nodularins. This ELISA test, using the β-amino acid 6E-ADDA as the epitope for antibody recognition, has a limit of quantification of 0.02–0.07 ng/mL, lower than the WHO-proposed guideline (1 ng/mL) for drinking water, irrespective of the sample matrix.
The recovery of the method was determined by analyzing real samples before and after the addition of pure microcystin and then subtracting the concentration of microcystin present in the sample prior to spiking. The matrix effect (i.e., effect of animal tissue) was checked by spiking control tissues with MC-LR standard (2 μg/g) and the response was compared to 100% methanol spiked with the same amount of the standard. Results are expressed as nanograms of MC-LR equivalents per gram of tissue.
Statistics
The general linear model (GLM) was used in order to examine if there are significant differences between animals, tissues and sampling periods in relation to the microcystin concentrations. Concentrations of microcystin in the tissues of individual animals were compared using One-way analysis of variance (ANOVA). Data were explored using the statistical package SPSS 19.0 for Windows.
Biomagnification of microcystins was examined, through the calculation of the biomagnification factor (BMF). BMF is determined as the average microcystin content in biota (ng MC-LReq g−1 DW) as a percentage of the average value in the diet of the respective organisms (microcystin in organism/microcystin in diet × 100).
Results
Intracellular and extracellular toxins in the water column and concentration in surface blooms and in zooplankton
According to phytoplankton identification, the lake dominated of Microcystis sp. during the study period, with the exception of June and July, when Anabaena was the dominant genus. Zooplankton identification showed that the dominant genus was Alona in November and Polyarthra in August.
Microcystin concentrations in seston (intracellular microcystins) varied considerably among months, ranging from 0.29 μg MC-LReq/L (in March) to 15.83 μg MC-LReq/L (in September) (Fig. 1). In August, microcystin concentrations reached the 14.2 μg MC-LReq/L (or 473.33 μg MC-LReq/g D.W.). In November, microcystin concentrations reached the 6.21 μg MC-LReq/L (or 390.56 μg MC-LReq/g D.W.). The lowest concentration of dissolved in the water column microcystins (extracellular microcystins), was found in December (0.32 μg MC-LReq/L). However, the highest concentration was found in October (4.12 μg MC-LReq/L) (Fig. 1). The microcystin concentration in surface cyanobacterial blooms ranged from 502 μg MC-LReq/g D.W. in November to 2,300 μg MC-LReq/g D.W. in August (F = 149.05, P < 0.05) (Fig. 2).

Monthly variation of intracellular and extracellular microcystin concentrations (μg MC-LReq/L) in Lake Pamvotis, during 2008

Microcystin concentrations (μg MC-LReq/g D.W.) in surface cyanobacterial bloom and zooplankton of Lake Pamvotis, during August and November 2008
Microcystin concentrations were found in all the zooplankton samples. In November, the microcystin concentration reached the 80 μg MC-LReq/g D.W. In August, microcystin concentration was higher and reached the 152 μg MC-LReq/g D.W (F = 132.19, P < 0.05) (Fig. 2).
Distribution of microcystins in the tissues of aquatic animals
Microcystin concentrations were detected in all the animals examined. In contrast, the control samples did not contain any detectable concentrations of microcystins. The GLM analysis showed that there were significant differences between the animals (F = 48.79, P < 0.05), the tissues (F = 121.82, P < 0.05) and the sampling periods (P < 0.05,) concerning microcystin concentration. The recovery of microcystins that we obtained from the spiked samples was 74 ± 2.3% for liver, 71 ± 3.5% for kidneys, 72.5 ± 5.2% for brain, and 70.6 ± 1.46% for muscle.
The matrix effect was negligible (from 0.08 to 5.2% of differences between matrix and methanol results with an average of 1.9 ± 0.7%). Similar results about the matrix effect with the ELISA test were observed by Ernst et al. (2005).
Distribution of microcystins in macroinvertebrates
Microcystin concentrations were detected in all the macroinvertebrates (Fig. 3). The highest microcystin concentrations were found in the mussel Anodonta cygnea. Microcystin concentrations were significant different among the tissues of the mussel (F = 132.81, P < 0.05). Also, microcystin concentrations in the tissues of the mussel differed between the sampling periods (F = 156.94, P < 0.05) with higher concentrations observed in August. High microcystin concentrations were detected in the hepatopancreas of the animal (1346.98 ± 164.78 ng MC-LReq/g and 884.97 ± 56.37 ng MC-LReq/g in August and November, correspondingly). Lower microcystin concentrations were detected in stomach (1188.53 ± 58.23 ng MC-LReq/g and 382.53 ± 86.01 ng MC-LReq/g in August and November, correspondingly), mantle (1150.49 ± 33.65 ng MC-LReq/g and 674.69 ± 137.38 ng MC-LReq/g in August and November, correspondingly), foot (1171.55 ± 108 ng MC-LReq/g and 799.07 ± 184.03 ng microcystin-LReq/g in August and November, correspondingly) and gills (1197.52 ± 52.17 ng MC-LReq/g and 841.69 ± 79.32 ng MC-LReq/g in August and November, correspondingly) (Fig. 3).

Microcystin concentrations (ng MC-LReq/g) in macroinvertebrates of Lake Pamvotis, during August and November 2008
Lower microcystin concentrations were detected in the shrimp Atyaephyra desmaresti and in the water snail Viviparus contectus (Fig. 3). Microcystin concentration in the body of the shrimp was higher in August (2250.48 ± 123.58 ng MC-LReq/g) than in November (749.15 ± 83.61 ng MC-LReq/g) (F = 89.73, P < 0.05). The water snail characterized by lower microcystin concentrations (1074.07 ± 147.84 ng MC-LReq/g and 684.96 ± 40.11 ng MC-LReq/g in August and November, correspondingly) (F = 103.54, P < 0.05).
Finally, the lowest microcystin concentrations among the macroinvertebrates, were detected in crayfish Astacus astacus (Fig. 3). Microcystin concentrations were significant different among the tissues of the crayfish (F = 108.65, P < 0.05). Also, microcystin concentrations in the tissues of the crayfish differed between the sampling periods (F = 56.84, P < 0.05) with higher concentrations observed in August. High microcystin concentrations were detected in the hepatopancreas of the animal (766.99 ± 148 ng MC-LReq/g and 354.82 ± 98.34 ng MC-LReq/g in August and November, correspondingly). Lower microcystin concentrations were detected in gills (701.09 ± 176.98 ng MC-LReq/g and 405.43 ± 91.46 ng MC-LReq/g in August and November, correspondingly), stomach (331.25 ± 70.41 ng MC-LReq/g and 126.94 ± 37.73 ng MC-LReq/g in August and November, correspondingly), muscle (328.61 ± 95.29 ng MC-LReq/g and 216.41 ± 65.71 ng MC-LReq/g in August and November, correspondingly), brain (312.93 ± 67.09 ng MC-LReq/g and 168.54 ± 42.64 ng MC-LReq/g in August and November, correspondingly) and gonads (301.76 ± 85.65 ng MC-LReq/g and 113.74 ± 26.45 ng MC-LReq/g in August and November, correspondingly) (Fig. 3).
The specimens of common carp (Cyprinus carpio) examined during the present study contained microcystins (Fig. 4). Significant statistical differences were found between the concentrations of microcystins in the tissues (F = 165.28; P < 0.05) and between the sampling periods (F = 56.95; P < 0.05). The highest level of microcystins was found in the liver (462.09 ± 125.54 ng MC-LReq/g and 367.09 ± 89.58 ng MC-LReq/g in August and November, correspondingly), followed by the kidney (318.76 ± 105.85 ng MC-LReq/g and 245.47 ± 13.98 ng MC-LReq/g in August and November, correspondingly), intestines (253.43 ± 106.25 ng MC-LReq/g and 258.69 ± 105.40 ng MC-LReq/g in August and November, correspondingly), and brain (240.01 ± 95.41 ng MC-LReq/g and 215.89 ± 79.83 ng MC-LReq/g in August and November, correspondingly). The lowest concentrations were detected in the muscle (119.07 ± 32.99 ng MC-LReq/g and 95.78 ± 30.79 ng MC-LReq/g in August and November, correspondingly) and gonads (54.63 ± 11.66 ng MC-LReq/g and 45.35 ± 13.98 ng MC-LReq/g in August and November, correspondingly) (Fig. 4).

Microcystin concentrations (ng MC-LReq/g) in tissues of fish (Cyprinus carpio) and frogs (Rana epirotica) originated from Lake Pamvotis, during August and November 2008
Microcystins in the tissues of fish and frogs
Samples of the endemic frog Rana epirotica also contained microcystins (Fig. 4). Significant statistical differences were found between the concentrations of microcystins in the tissues (F = 121.98; P < 0.05) and between the sampling periods (F = 78.01; P < 0.05). The highest level of microcystins was found in liver (581.46 ± 92.83 ng MC-LReq/g and 208.69 ± 89.58 ng MC-LReq/g in August and November, correspondingly), followed by the pancreas (554.06 ± 98.06 ng MC-LReq/g and 141.59 ± 51.32 ng MC-LReq/g in August and November, correspondingly), intestine (320.56 ± 63.98 ng MC-LReq/g and 102.59 ± 18.74 ng MC-LReq/g in August and November, correspondingly), and skin (419.52 ± 91.62 ng MC-LReq/g and 80.92 ± 23.56 ng MC-LReq/g in August and November, correspondingly). The lowest concentrations were detected in the muscle (325.49 ± 87.01 ng MC-LReq/g and 88.12 ± 20.85 ng MC-LReq/g in August and November, correspondingly) and gonads (164.62 ± 34.02 ng MC-LReq/g and 68.97 ± 15.74 ng MC-LReq/g in August and November, correspondingly) (Fig. 4).
The recovery of microcystins that we obtained from the spiked: (1) bloom samples was 92%, (2) zooplankton samples was 76%, (3) macroinvertebrate samples was 74%, (4) fish samples was 74%, (5) frog samples was 68%. The matrix effect was negligible (from 0.08 to 5.2% of differences between matrix and methanol results with an average of 1.9 ± 0.7%).
Biomagnification of microcystins
Biomagnification factors (BMF) ranged from 0.19 (in Atyaephyra desmaresti), to 32.1 (in zooplankton) (Table 1). Microcystin concentration in none of the food web compartments exceeded the concentration in seston. In general no evidence for biomagnification was found. Nevertheless, it is remarkable that all biomagnification factors were higher in August than November, with the exception of the biomagnification factor in fish species Cyprinus carpio. In August, BMF in Cyprinus carpio was 3.7, lower than in November (5.9).
Discussion
Cyanobacterial blooms have received the attention of many scientists worldwide, due to their destructive impact on the quality of many fresh and brackish waters. The production of cyanotoxins hinders the function and structure of waterbodies (Codd 2000). Moreover, cyanotoxins could be accumulated in animals through food chain, causing health problems, even to humans (Vasconcelos 2006). According to our results, significant concentrations of microcystins were found in the water along with the total of aquatic organisms of Lake Pamvotis. This is in agreement with study of Papadimitriou et al. (2011), who found significant microcystin concentrations (extacellular microcystins: 0.0121–7.88 μg/L, intracellular microcystins: 0.15–15.21 μg/L) in water of Lake Pamvotis, along with toxicity effects of natural cyanobacterial bloom on fish embryos. Phytoplankton appeared with the higher microrocystin concentrations, followed by zooplankton, macroinvertebrates, frogs and fish. Microcystin concentrations found in seston of Lake Pamvotis were higher in warm months when Microcystis sp. was the dominant phytoplankton species. The above concentrations were similar to microcystin concentrations found to other Mediterranean lakes located in Turkey, Spain, France and Algeria (Briand et al. 2002; Albay et al. 2003; Romo et al. 2011). Moreover, cyanobacterial bloom from Lake Pamvotis characterized by high concentrations of microcystins, similar to those found Ghazali et al. (2009) in Lake Lalla Tarkekoust (Morocco) and Zhang et al. (2009) in Lake Taihu (China) while recently, cyanobacterial species as Anabaena flos-aquae, Aphanizomenon flos-aquae and Microcystis sp. were recorded as dominant species along with the presence of the microcystin producing gene mcy-A which also has been detected (Vareli et al. 2009).
The increase of microcystin concentrations in seston of Lake Pamvotis was followed by an increase of microcystin concentrations in zooplankton. This is the first record of microcystin concentrations in zooplankton of a Mediterranean lake. According to Ferrão-Filho and Kozlowsky-Suzuki (2011), zooplankton is clearly the best bioaccumulator of microcystins, reaching values over 1,000 μg g−1 DW and an average value of about 383 μg g−1 DW. However, recent studies have shown that some zooplankton species not only are able to co-exist with toxic cyanobacteria and even growth in their presence (Leonard and Pearl 2005; Hansson et al. 2007), but also that a sensitive species can indeed develop tolerance when pre-exposed to toxic cyanobacteria (Gustafsson and Hansson 2004; Gustafsson et al. 2005; Guo and Xie 2006). For example negative physiological effects and negative fitness consequences on Daphnia spp. have been observed (Rohrlack et al. 2001, 2005; Gustafsson et al. 2005) while some populations of Daphnia galeata and Daphnia pulicaria exposed to cyanobacteria can adapt to grow on and being more tolerant (Sarnelle and Wilson 2005). Microcystin concentrations in zooplankton of Lake Pamvotis are similar to those detected in zooplankton of Lake Ijsselmeer in Netherlands (57–192 μg MC-LReq/g D.W.) (Ibelings et al. 2005) but higher than those detected in zooplankton of Jacarepagua Lagoon in Brazil (0.3–16.4 μg MC-LReq/g D.W.) (Ferrão-Filho et al. 2002). These differences in microcystin concentrations are probably associated with the different zooplankton community structure and functional complexity of each lake. Moreover, high intraspecific (clonal) variation in the sensitivity to toxic cyanobacteria has also observed, maybe due to different trophic state of the lakes (Sarnelle and Wilson 2005). Some observations suggest that smaller species of herbivorous zooplankton such as rotifers, copepods, and small cladocerans are less susceptible to cyanobacteria than larger zooplankton (Ferrão-Filho et al. 2002). According to Papadimitriou (2010), zooplankton community of Lake Pamvotis is dominated by small-sized species, thus, zooplankton of Lake Pamvotis could avoid feeding on cyanobacteria and consequently the accumulation of high microcystin concentrations due to its small size.
Microcystins can also be accumulated on other aquatic invertebrates such as bivalves, crustaceans and gastropods. According to our results, high concentrations of mycrocystins detected in tissues of Anodonta cygnea, Atyaephyra desmaresti and Viviparus contectus. According to previews studies, gastropods may contain concentrations up to 436 μg g−1 DW (Sinotaia histrica in hepatopancreas, Ozawa et al. 2003), with an average of 58 μg g−1 DW. Although bivalves are efficient filter-feeders, maximum concentration was 630 μg g−1 DW (Unio douglasiae hepatopancreas, Yokoyama and Park 2003), and the average was only 57 μg MC g−1 DW. Larger crustaceans (i.e., Decapoda) seem to be the less efficient accumulators of microcystins, with an average of 4.2 μg g−1 DW and a maximum concentration of 12.4 μg g−1DW (Macrobrachium nipponensis stomach, Chen and Xie 2005b). According to our results, higher microcystin concentrations were accumulated in the hepatopancreas of Anodonta cygnea indicating that this is the target organ for microcystins. Nevertheless, Anodonta cygnea accumulated microcystins also in other tissues, possible due to the well documented process of microcystin circulation (Fischer et al. 2005). However, bioaccumulation patterns depend on the mussel species (Chen and Xie 2005a; Martins and Vasconcelos 2009). It is worth note that according to our results, zooplankton accumulated higher microcystin concentrations than the mussel Anodonta cygnea. An explanation would be that the mussels are more resistant than zooplankton to microcystins, appearing selective rejection of cyanobacteria (Ibelings et al. 2005). Microcystin concentrations found in the body of Viviparus contectus from Lake Pamvotis were lower than those (19.5 μg MC-LReq/g) found in the gastropod Sinotaia histrica from Lake Biwa in Japan (Ozawa et al. 2003). However, accumulation of microcystins in gastropods is of specific interest, as these organisms are important links between primary producers and higher consumers. Viviparus contectus from Lake Pamvotis probably accumulated the toxins through consumption of Microcystis sp. presented in lake water and/or on the leaves of aquatic plants. This is in accordance with studies of Lance et al. (2006) who suggested that gastropods accumulate microcystins mainly by grazing toxic phytoplankton, and to a lesser extent, via uptake of dissolved toxins.
In the present study, the presence of high microcystin concentrations in the freshwater shrimp Atyaephyra desmaresti is reported. As far as we know there is no previous evidence concerning the accumulation ability of this species. Atyaephyra desmaresti feeds on algae and decaying matter (Anastasiadou Ch et al. 2004) and probably accumulates microcystins through its diet or/and during the continuous contact with the aquatic environment. Atyaephyra desmaresti is included in the diet of many fish species, thus it may be a key organism for the transfer of microcystins through the food chain. We also observed microcystins in tissues of crayfish Astacus astacus. It was confirmed that microcystins are accumulated primarily in their hepatopancreas, however microcystin concentrations were found also in the other tissues. Unexpectedly, significant concentrations of microcystins were found in the brain, suggesting the ability of microcystins to be transported to the brain. The presence of microcystins in the gonads of Astacus astacus, is in accordance with the studies of Chen and Xie (2005b) about the transport of microcystins from adults crustaceans to embryos crustaceans (Palaemon modestus and Macrobrachium nipponensis). Lower microcystins accumulated in the muscles of Palaemon modestus and Macrobrachium nipponensis. This, in addition to the fact that Astacus astacus has a commercial value, increases the risk of human contamination through the consumption of this animal. Chen and Xie (2005b) also found microcystin concentrations in hepatopancreas (8.40 μg MC-LReq/g) and in muscle (0.53 μg MC-LReq/g) of the shrimp Palaemon modestu. The same authors also found microcystin concentrations in the muscle (0.05 μg MC-LReq/g), stomach (9.97 μg MC-LReq/g) and hepatopancreas (0.08 μg MC-LReq/g) of the crayfish Procambarus clarkii.
Microcystins are toxic to fish at concentrations as low as a few micrograms per Liter (mg/L) or possibly even fractional μg/L (reviewed by Wiegand and Pflugmacher 2005; Malbrouck and Kestemont 2006). Considering that microcystins have been measured in concentrations up to 25,000 μg/L in water ecosystems with cyanobacterial blooms (reviewed by W.H.O. 1998), it is not surprising that potential impacts on fish are receiving increased attention. Fish typically are exposed to microcystins either during ingesting cyanobacteria or prey that has fed on cyanobacteria (Malbrouck and Kestemont 2006). Previews studies, have focused on toxicity of microcystins in Cyprinus carpio, under laboratory conditions (Li et al. 2004; Palikova et al. 2007; Adamovsky et al. 2007) thus there is only limited information on microcystin accumulation in tissues of Cyprinus carpio, under field conditions at environmental relevant concentrations. According to our results, the microcystin content (extracellular and intracellular) of the lake water was sufficient to promote microcystin accumulation in the tissues of fish species Cyprinus carpio. Even though the target organ for microcystins is thought to be the liver, microcystins were also found in the rest of tissues, including muscle tissue. Unexpectedly, it was also found high levels of microcystins in the brain of the fish. Thus, our results give additional evidence on the ability of microcystins to cross through the blood–brain barrier and rose questions on the probable neurotoxicity of microcystins. Common carps are generally omnivorus but obligate herbivorus and also known as cyanobacterivorous (Fischer and Dietrich 2000a) while it is thought that M.aeruginosa comprises a significant portion of their diet when bloom formation occurs (Carbis et al. 1996). Zhang et al. (2009) also found significant microcystin concentrations in liver (0.025 μg MC-LReq/g), intestinal wall (0.032 μg MC-LReq/g), gonads (0.027 μg MC-LReq/g) and muscles (0.067 μg MC-LReq/g) of Cyprinus carpio originated from the eutrophic Lake Taihu in China providing more evidence about the increased succesibility of carps on cyanotoxins in eutrophicated lakes (Tencalla et al. 1994; Xie et al. 2007). It seems that in eutrophicated lakes cyprinids can accumulate toxins surviving at sub-lethal concentrations.
We also found significant microcystin concentrations in various tissues of the endemic frog (Rana epirotica). Bioaccumulation of microcystins in amphibians may be critical if these represent linkages to terrestrial environments (White et al. 2007). For example, pre-metamorphic stages (tadpoles) may be consumed by terrestrial birds, thus allowing toxin to enter terrestrial food webs. However, until now, only one report refers to accumulation of microcystins in amphibians (Gkelis et al. 2006) and few reports exist on microcystin toxicity in amphibians (Gromov et al. 1997; Oberemm et al. 1999; Fischer and Dietrich 2000b; Dvorakova et al. 2002; White et al. 2007). The higher microcystin concentrations accumulated in liver and pancreas confirming the preferential accumulation in those organs. Lower concentrations were found in intestines, gonads and muscle probably as a result of microcystin transport via the bloodstream, or bypassing the hepatopancreas, or downstream of the hepatopancreas subsequent to an overload of presystemic hepatic elimination capacities (Fischer and Dietrich 2000a). Significant microcystin concentrations were detected in the skin of the frog, suggesting that transdermal uptake of extracellular microcystins is possible. This is in agreement with studies of Gromov et al. (1997) who found that larvae of the frog Rana temporaria died after exposure with lyophilised Microcystis aeruginosa biomass. These larvae were so young that they would not have been feeding during the experimental period, thus only transdermal uptake of extracellular toxin was possible. Gkelis et al. (2006) have also found similar microcystin concentrations in viscera (465 ng MC-LReq/g) and muscle (262 ng MC-LReq/g) of Rana epirotica from Lake Pamvotis. Our study along with other related studies, support that algal toxin exposure may be a contributing factor in the recent worldwide decline of amphibians, as reported in Kiesecker et al. (2001), especially since it is well known that cyanobacterial toxic blooms pose an increased risk globally (de Figueiredo et al. 2004).
The risk of being exposed to toxins through food consumption would be further increased if biomagnification were to take place, since some popular food, like fish species may stand at the top of the food web. Thus, the estimation of the BMF in a lake with presence of toxic cyanobacteria would be valuable for the protection of public health. The calculation of BMF in Lake Pamvotis, showed that there is no magnification of microcystins through foodweb. BMF was higher when microcystin concentrations in seston were higher too. However, BMF concerning the fish species Cyprinus carpio was higher in November, probably due to the ability of fish to reduce their metabolic activities during the cold period. Rather than biomagnification, the opposite process, i.e., biodilution of hepatotoxins in the food web is supported by part of the literature (Engström-Öst et al. 2002; Sipia¨ et al. 2004; Ibelings et al. 2005; Karjalainen et al. 2005). However, the fact that microcystins are found in the tissues of organisms caught from Lake Pamvotis, clearly indicates that transfer of microcystins within the foodweb takes place, thus may be a potential health risk to human. The species Astacus astacus and Cyprinus carpio are considered as commercial species concerning inland fisheries while the latter is among the most important aquaculture products in many regions of the world included Greece. Additionally, the frog Rana epirotica is considered as a local food which attracts the liking of many tourists.
Estimated daily intake (EDI) calculations, included in our discussion, are aimed at assessing the potential exposure of humans to toxins thus contributing in the context of public health. Regarding at the risk to humans, EDI estimation is necessary before comparing our measured concentrations of microcystins in the edible tissue samples to the World Health Organization (WHO) tolerable daily intake (TDI) provisional guidelines. At the levels found in muscle tissue of Astacus astacus in our study the calculated EDI was two times (in November) and six times (in August) higher than the advisable TDI value (0.04 μg/kg of body weight). In muscle of Cyprinus carpio in our study the calculated EDI was 11 times (in November) and 14 times (in August) higher than the advisable TDI value. In muscle of Rana epirotica the calculated EDI was one time (in November) and six times (in August) higher than the advisable TDI value (Fig. 5). Therefore, Astacus astacus, Cyprinus carpio and Rana epirotica from Lake Pamvotis could possibly pose a health hazard and they were not safe for human consumption. Finally, it is necessary to point out that only uptake of microcystins from aquatic organisms was used to evaluate exposure risk of human to microcystins. However, water is often the major route for human microcystins’ intoxication. In the present study, concentration of dissolved microcystins in the water samples of Lake Pamvotis ranged from 0.32 μg MC-LReq/L to 4.12 μg MC-LReq/L, with the values in warm period to be well above the safety limit of 1 μg/L MC-LR for drinking water. However, the amount of dissolved and intracellular microcystins found in Lake Pamvotis, poses moderate health risks through recreational activities (W.H.O. 1998). Although Lake Pamvotis is not used as source of drinking water, the combined risks from microcystin contaminated water and consumption of contaminated aquatic organisms, to human health cannot be overlooked in the region around Lake Pamvotis.

Estimated daily intake (EDI) of microcystins based on a 60 kg person ingesting a 300 g serving of Cyprinus carpio or 50 g serving of Astacus astacus, or 50 g of Rana epirotica from Lake Pamvotis, during November and August of 2008. The horizontal line represents the tolerable daily total intake (TDI) established by WHO
References
Adamovsky O, Kopp R, Hilscherova K, Babica P, Palikova M, Paskova V, Navratil S, Blaha L (2007) Microcystin kinetics (bioaccumulation, elimination) and biochemical responses in common carp and silver carp exposed to toxic cyanobacterial blooms. Environ Toxicol Chem 26:687–2693
Albay M, Akcaalan R, Tufekci H, Metcalf J, Beattie K, Codd G (2003) Depth profiles of cyanobacterial hepatotoxins (microcystins) in three Turkish freshwater lakes. Hydrobiology 505:89–95
Amorim A, Vasconcelos V (1999) Dynamics of microcystins in the mussel Mytilus galloprovincialis. Toxicon 37:1041–1052
Anagnostidis K, Economou-Amilli A (1980) Limnological studies on Lake Pamvotis (Ioannina), Greece I. Hydroclimatology, phytoplankton-periphyton with special reference to the valence of some microorganisms from sulphureta as bioindicators. Arch Hydrobiol 89:313–342
Anastasiadou Ch, Koukouras A, Mavidis M, Chartosia N, Mostakim Md, Christodoulou M, Aslanoglou Ch (2004) Morphological variation in Atyaephyra desmarestii (Millet, 1831) within and among populations over its geographical range. Mediterr Mar Sci 5(2):5–13
Anderson DM, Glibert PM, Burkholder JM (2002) Harmful algal blooms and eutrophication: nutrient sources, composition, and consequences. Estuaries 25:704–726
Babcock-Jackson L, Carmichael WW, Culver DA (2002) Dreissenid mussels increase exposure of benthic and pelagic organisms to toxic microcystins. Verh Internat Verein Limnol 28:1082–1085
Beresovsky D, Hadas O, Livne A, Sukenik A, Kaplan A, Carmeli S (2006) Toxins and biologically active secondary metabolites of Microcystis sp. isolated from Lake Kinneret. Israel J Chem 46:79–87
Briand JF, Robillot C, Quiblier-Lloberas C, Bernard C (2002) A perennial bloom Planktothrix agardhii (Cyanobacteria) in a shallow eutrophic French lake: limnological and microcystin production studies. Arch Hydrobiol 153:605–622
Carbis CR, Mitchell GF, Anderson JW, McCauley I (1996) The effects of microcystins on the serum biochemistry of carp, Cyprinus carpio L., when the toxins are administered by gavage, immersion and intraperitoneal routes. J Fish Dis 19:151–159
Carmichael WW (1994) The toxins of cyanobacteria. Sci Am 270:78–86
Carmichael WW (1997) The Cyanotoxins. In: Callow J (ed) Advances in botanical research, vol. 27. Academic Press, London, pp 211–256
Chen J, Xie P (2005a) Seasonal dynamics of the hepatotoxic microcystins in various organs of four freshwater bivalves from the large eutrophic Lake Taihu of subtropical China and the risk to human consumption. Environ Toxicol 20:572–584
Chen J, Xie P (2005b) Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and -RR in two freshwater shrimps. Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtrophical China. Toxicon 45:615–625
Codd GA (1995) Cyanobacterial toxins: occurrence, properties and biological significance. Water Sci Technol 32:149–156
Codd GA (2000) Cyanobacterial toxin, the perception of water quality, and the prioritisation if eutrophication control. Ecol Eng 16:51–60
de Figueiredo DR, Azeiteiro UM, Esteves SM, Goncalves FJ, Pereira MJ (2004) Microcystin-producing blooms—a serious global public health issue. Ecotox Environ Saf 59:151–163
DeMott WR (1999) Foraging strategies and growth inhibition in five daphnids feeding on mixtures of a toxic cyanobacterium and green alga. Freshw Biol 42:263–274
Duy TN, Lam PKS, Shaw GR, Connell DW (2000) Toxicology and risk assessment of freshwater cyanobacterial (Blue- Green Algal) toxins in water. Rev Environ Contam Toxicol 163:113–186
Dvorakova D, Dvorakova K, Blaha L, Marsalek B, Knotkova Z (2002) Effects of cyanobacterial biomass and purified microcystins on malformations in Xenopus laevis: teratogenesis assay (FETAX). Environ Toxicol 17:547–555
Engström-Öst J, Lehtiniemi M, Green S, Kozlowsky-Suzuki B, Viitasalo M (2002) Does cyanobacterial toxin accumulate in mysid shrimps and fish via copepods? J Exp Mar Biol Ecol 276:95–107
Ernst B, Dietz L, Hoeger SJ, Dietrich DR (2005) Recovery of MC-LR in fish liver tissue. Environ Toxicol 20:449–458
Falconer IR, Humpage AR (2005) Health risk assessment of cyanobacterial (blue-green algal) toxins in drinking water. Int J Environ Res Public Health 2:43–50
Ferrão-Filho AS, Kozlowsky-Suzuki Betina (2011) Cyanotoxins: bioaccumulation and effects on aquatic animals. Mar Drugs 9:2729–2772
Ferrão-Filho AS, Domingos P, Azevedo SMFO (2002) Influences of a Microcystis aeruginosa Kützing bloom on zooplankton populations in Jacarepaguá Lagoon (Rio de Janeiro, Brazil). Limnologica 32:295–308
Fischer W, Dietrich D (2000a) Pathological and biochemical characterization of MC-induced hepatopancreas and kidney damage in carp (Cyprinus carpio). Toxicol Appl Pharmacol 164:73–81
Fischer WJ, Dietrich DR (2000b) Toxicity of the cyanobacterial cyclic heptapeptide toxins microcystin-LR and-RR in early life-stages of the African clawed frog (Xenopus laevis). Aquat Toxicol 49:189–198
Fischer WJ, Altheimer S, Cattori V, Meier PJ, Dietrich DR, Hagenbuch B (2005) Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol Appl Pharmacol 203:257–263
Ghazali I, Saqrane S, Carvalho AP, Ouahid Y, Oudra B, Del Campo FF, Vasconcelos V (2009) Compensatory growth induced in Zebrafish Larvae after pre-exposure to a Microcystis aeruginosa natural bloom extract containing microcystins. Int J Mol Sci 10:133–146
Gkelis S, Lanaras T, Sivonen K (2006) The presence of microcystins and other cyanobacterial bioactive peptides in aquatic fauna collected from Greek freshwaters. Aquat Toxicol 78(1):32–41
Gromov B, Mamkaeva K, Filatova E (1997) Effect of toxigenic strains of cyanobacterium Microcystis aeruginosa on the larvae of frog Rana temporaria. Doklady Akademii Nauk (Proceedings of the Russian Academy of Science) 356:422–423 (Original article in Russian)
Guo N, Xie P (2006) Development of tolerance against toxic Microcystis aeruginosa in three cladocerans and the ecological implications. Environ Pollut 143:513–518
Gustafsson S, Hansson L (2004) Development of tolerance against toxic cyanobacteria in Daphnia. Aquat Ecol 38:37–44
Gustafsson S, Rengefors K, Hansson LA (2005) Increased consumer fitness following transfer of toxin tolerance to offspring via maternal effects. Ecology 86:2561–2567
Habdija I, Latjner J, Belinic I (1995) The contribution of gastropod biomass in macrobenthic communities of a karstic river. Int Rev Ges Hydrobiol 80:103–110
Hansson L, Gustafsoson S, Rengefors K, Bomark L (2007) Cyanobacterial chemical warfare affects zooplankton community composition. Freshw Biol 52:1290–1301
Hooser SB (2000) Fulminant hepatocyte apoptosis in vivo following microcystin-LR administration to rats. Toxicol Pathol 28:726–733
Hooser SB, Beasley VR, Lovell RA, Carmichael WW, Haschek WM (1989) Toxicity of microcystin LR, a cyclic heptapeptide hepatotoxin from Micocystis aeruginosa, to rats and mice. Vet Pathol 26:246–252
Ibelings BW, Bruning K, de Jonge J, Wolfstein K, Dionisio PLM, Postma J, Burger T (2005) Distribution of microcystins in a lake foodweb: no evidence for biomagnification. Microbial Ecol 49(4):487–500
Kagalou I, Papastergiadou E, Tsimarakis G, Petridis D (2003) Evaluation of the trophic state of Lake Pamvotis Greece, a shallow urban Lake. Hydrobiology 506–509:745–752
Kagalou I, Papadimitriou T, Bacopoulos V, Leonardos I (2008) Assessment of microcystins in lake water and the omnivorous fish (Carassius gibelio, Bloch) in Lake Pamvotis (Greece) containing dense cyanobacterial bloom. Environ Monit Assess 137:185–195
Karjalainen M, Reinikainen M, Spoof L, Meriluoto JAO, Sivonen K (2005) Trophic transfer of cyanobacterial toxins from zooplankton to planktivores: consequences for pike larvae and mysid shrimps. Environ Toxicol 20(3):354–362
Khan TA (2003) Dietary studies on exotic carp (Cyprinus carpio L.) from two lakes of western Victoria, Australia. Aquat Sci 65:272–286
Kiesecker JM, Blaustein AR, Belden LK (2001) Complex causes of amphibian population declines. Nature 410:681–684
Kotak BJ, Semalulu S, Friytz DL, Prepas EE, Hrudey SE, Coppock RW (1996) Hepatic and renal pathology of intraperitoneally administered microcystin-LR in rainbow trout (Oncorhynchus mykiss). Toxicon 34:517–525
Kotti M, Vlessidis A, Evmiridis N (2000) Determination of phosphorous and nitrogen in the sediment of Lake Pamvotis (Greece). Int J Environ Anal Chem 78(3–4):455–467
Lance E, Brient L, Bormans M, Gérard C (2006) Interactions between cyanobacteria and Gastropods I. Ingestion of toxic Planktothrix agardhii by Lymnaea stagnalis and the kinetics of microcystin bioaccumulation and detoxification. Aquat Toxicol 79:140–148
Lawton LA, Edwards C, Codd GA (1994) Extraction and high-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst 119:1525–1530
Leonard JA, Pearl HW (2005) Zooplankton community structure, micro-zooplankton grazing impact, and seston energy content in the St. Johns River system, Florida as influenced by the toxic cyanobacterium Cylindrospermopsis raciborskii. Hydrobiology 537:89–97
Li XY, Chung IK, Kim JI, Lee JA (2004) Subchronic oral toxicity of microcystin in common carp (Cyprinus carpio L.) exposed to microcystis under laboratory conditions. Toxicon 44:821–827
Liras V, Lindberg M, Nyström P, Annadotter H, Lawton L, Graf B (1998) Can ingested cyanobacteria be harmful to the signal crayfish (Pacifastacus leniusculus). Freshw Biol 39:233–242
Mackie GL (1984) Bivalves. In: Wilbur KM (ed) The Mollusca, vol. 7, reproduction. Academic Press, Orlando, pp 351–418
Magalhães VF, Soares R, Azevedo S (2001) Microcystin contamination in fish from the Jacarepagua Lagoon (Rio de Janeiro, Brazil): ecological implication and human health risk. Toxicon 39:1077–1085
Magalhães VF, Marinho MM, Domingos P, Oliveira AC, Costa SM, Azevedo LO, Azevedo SMFO (2003) Microcystins (cyanobacteria hepatotoxins) bioaccumulation in fish and crustaceans from Sepetiba Bay (Brasil, RJ). Toxicon 42:289–295
Malbrouck C, Kestemont P (2006) Effects of microcystins on fish. Environ Toxicol Chem 25:72–86
Malbrouck C, Trausch G, Devos P, Kestemont P (2003) Hepatic accumulation and effects of microcystin-LR on juvenile goldfish Carassius auratus L. Comp Biochem Physiol Part C 135(1):39–48
Martins JC, Vasconcelos VM (2009) Microcystin dynamics in aquatic organisms. J Toxicol Environ Health 12:65–82
Oberemm A, Becker J, Codd GA, Steinberg C (1999) Effects of cyanobacterial toxins and aqueous crude extracts of cyanobacteria on the development of fish and amphibians. Environ Toxicol 14:77–88
Ozawa K, Yokoyama A, Ishikawa K, Kumagai M, Watanabe MF, Park HD (2003) Accumulation and depuration of microcystin produced by the cyanobacterium Microcystis in a freshwater snail. Limnology 4:131–138
Paliková M, Krejč R, Hilscherová K, Babica P, Navratil S, Kopp R, Blaha L (2007) Effect of different cyanobacterial biomasses and their fractions with variable microcystin content on embryonal development of carp (Cyprinus carpio L.). Aquat Toxicol 81: 312–318
Papadimitriou T (2010) Effects of Microcystins on aquatic organisms. Dissertation, University of Ioannina
Papadimitriou T, Kagalou I, Bacopoulos V, Leonardos I (2010) Accumulation of microcystins in water and fish tissues: an estimation of risks associated with microcystins in most of the Greek lakes. Environ Toxicol 25(4):418–427
Papadimitriou T, Armeni E, Stalikas C, Kagalou I, Leonardos I (2011) Detection of microcystins in Pamvotis lake water and assessment of cyanobacterial bloom toxicity. Environ Monit Assess. doi:10.1007/s10661-011-2169-5
Pennak RW (1978) Freshwater invertebrates of the United States. Wiley-Interscience, New York. 2nd edn, 803 pp
Prepas EE, Kotak BG, Campbell LM, Evans JC, Hrudey SE, Holmes CF (1997) Accumulation and elimination of cyanobacterial hepatotoxins by the freshwater clam Anodonta grandis simpsonia. Can J Fish Aquat Sci 54:41–46
Ressom R, Soong FS, Fitzgerald J, Turczynowicz L, El Saadi O, Roder D, Maynard T, Falconer IR (1994) Health effects of toxic cyanobacteria (blue-green algae). National Health and Medical Council, Australian Government Publishing Service, Canberra
Rohrlack T, Dittman E, Borner T, Christoffersen K (2001) Effects of cell-bound microcystins on survival and feeding of Daphnia spp. Appl Environ Microbiol 67(8):3523–3529
Rohrlack T, Christoffersen K, Dittmann E, Nogueira I, Vasconcelos V, Borner T (2005) Ingestion of microcystins by Daphnia: intestinal uptake and toxic effects. Limnol Oceanogr 50:440–448
Romo S, Francisca F, Youness O, Ángel B-S (2011) Assessment of microcystins in lake water and fish (Mugilidae, Liza sp.) in the largest Spanish coastal lake. Environ Monit Assess 184:939–949
Sarnelle O, Wilson AE (2005) Local adaptation of Daphnia pulicaria to toxic cyanobacteria. Limnol Oceanogr 50:1565–1570
Sipia V, Lahti K, Kankaanpaa H, Vuorinen P, Meriluoto J (2002) Screening for cyanobacterial hepatotoxins in herring and salmon from the Baltic Sea. Aquat Ecosyst Health Manag 5(4):451–456
Sipiä VO, Karlsson KM, Meriluoto JAO, Kankaanpää HT (2004) Eiders (Somateria mollissima) obtain nodularin, a cyanobacterial hepatotoxin, in Baltic Sea food web. Environ Toxicol Chem 23:1256–1260
Tencalla F, Dietrich D, Schlatter C (1994) Toxicity of Microcystis aeruginosa peptide toxin to yearling raibowtrout (Oncorhynchus mykiss). Aquat Toxicol 30:215–224
Thostrup L, Christoffersen K (1999) Accumulation of microcystin in Daphnia magna feeding on toxic Microcystis. Archiv fūr Hydrobiologie 145:447–467
Vardaka E, Moustaka-Gouni M, Cook CM, Lanaras T (2005) Cyanobacterial blooms and water quality in Greek waterbodies. J Appl Phycol 17:391–401
Vareli K, Pilidis G, Mavrogiorgou MC, Briasoulis E, Sainis I (2009) Molecular characterization of cyanobacterial diversity and yearly fluctuations of microcystin loads in a suburban Mediterranean lake (Lake Pamvotis, Greece). J Environ Monit 11:1506–1512
Vasconcelos VM (2006) Eutrophication, toxic cyanobacteria and cyanotoxins: when ecosystems cry for help. Limnetica 25(1–2):425–432
White SH, Duivenvoorden LJ, Fabbro LD, Eaglesham GK (2007) Mortality and toxin bioaccumulation in Bufo marinus following exposure to Cylindrospermopsis raciborskii cell extracts and live cultures. Environ Pollut 147:158–167
WHO (1998) Cyanobacterial toxins: microcystin-LR. In: Guidelines for drinking water quality (ed) Addendum to vol. 2. Health criteria and other supporting information, 2nd edn. World Health Organization, Geneva, pp. 95–110
Wiegand C, Pflugmacher S (2005) Ecotoxicological effects of selected cyanobacterial secondary metabolites: a short review. Toxicol Appl Pharmacol 203:201–218
Xie LQ, Xie P, Ozawa K, Honma T, Yokoyama A, Park HD (2004) Dynamics of microcystins-LR and -RR in the phytoplanktivorous silver carp in a sub-chronic toxicity experiment. Environ Pollut 127:431–439
Xie L, Yokoyama A, Nakamura K, Park H (2007) Accumulation of microcystins in various organs of the freshwater snail Sinotaia histrica and three fishes in a temperate lake, the eutrophic Lake Suwa, Japan. Toxicon 49:646–652
Yokoyama A, Park HD (2003) Depuration kinetics and persistence of the cyanobacterial toxin microcystin-LR in the freshwater bivalve Unio douglasiae. Environ Toxicol 18:61–67
Yu SZ (1995) Primary prevention of hepatocellular carcinoma. J Gastroenterol Hepatol 10:674–682
Zhang DW, Xie P, Liu YQ, Qiu T (2009) Transfer, distribution and bioaccumulation of microcystins in the aquatic food web in Lake Taihu, China, with potential risks to human health. Sci Total Environ 407:2191–2199
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Papadimitriou, T., Kagalou, I., Stalikas, C. et al. Assessment of microcystin distribution and biomagnification in tissues of aquatic food web compartments from a shallow lake and evaluation of potential risks to public health. Ecotoxicology 21, 1155–1166 (2012). https://doi.org/10.1007/s10646-012-0870-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-012-0870-y