
Abstract
Common loons (Gavia immer) can be exposed to relatively high levels of dietary methylmercury (MeHg) through fish consumption, and several studies have documented MeHg-associated health effects in this species. To further study the neurological risks of MeHg accumulation, migrating loons dying of Type E botulism were collected opportunistically from the Lake Erie shore at Long Point (Ontario, Canada) and relationships between total mercury (THg), selenium (Se), and selected neurochemical receptors and brain enzymes were investigated. THg concentrations were 1–78 μg/g in liver; and 0.3–4 μg/g in the brain (all concentrations reported on a dry weight basis). A significant (p < 0.05) positive correlation was found between THg in liver and THg in 3 subregions of the brain (cerebral cortex: r = 0.433; cerebellum: r = 0.293; brain stem: r = 0.405). THg varied significantly among different brain regions, with the cortex having the highest concentrations. Se levels in the cortex and cerebellum were 1–29 and 1–10 μg/g, respectively, with no significant differences between regions. Se was not measured in brain stem due to insufficient tissue mass. There were molar excesses of Se over mercury (Hg) in both cortex and cerebellum at all Hg concentrations, and a significant positive relationship between THg and the Hg:Se molar ratio (cortex: r = 0.63; cerebellum: r = 0.47). No significant associations were observed between brain THg and the n-methyl-d-aspartic acid (NMDA) receptor concentration, nor between THg and muscarinic cholinergic (mACh) receptor concentration; however, brain THg levels were lower than in previous studies that reported significant Hg-associated changes in neuroreceptor densities. Together with previous studies, the current findings add to our understanding of Hg distribution in the brain of common loons, and the associations between Hg and sub-lethal neurochemical changes in fish-eating wildlife.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wild fish-eating birds are at relatively high risk for elevated dietary methylmercury (MeHg) exposure, as a result of biomagnification of MeHg through aquatic food chains (Scheuhammer et al. 2007). Numerous studies have demonstrated negative health effects of MeHg in various avian species, with impaired reproduction and behavioral changes being frequently reported. MeHg altered parental behavior in captive American kestrels (Falco sparverius) (Albers et al. 2007), resulting in an increased number of infertile eggs due to impaired mating coordination (Bennett et al. 2009). Captive great egret (Ardea alba) nestlings fed a diet containing MeHg displayed nervous system lesions and altered neurological function including tremors, posture changes, and uneven gait (Spalding et al. 2000). One of the best studied wildlife species with respect to toxic effects of environmentally realistic exposures to MeHg is the common loon (Gavia immer). In wild common loons, dietary MeHg exposure has been associated with behavioral alterations resulting in reduced productivity and altered parent–chick relationships (Barr 1986; Burgess and Meyer 2008; Evers et al. 2008), and in captive dosing studies, dietary MeHg exposure caused immunosupression (Kenow et al. 2007) and changes in loon behavior (Kenow et al. 2010) Many of these effects were observed following exposure of birds to levels of dietary MeHg that are relevant in the Great Lakes ecosystem.
Although MeHg is well known as a potent neurotoxicant, its specific effects on neurochemical pathways have only recently begun to be studied in wild avian species (Scheuhammer et al. 2008; Rutkiewicz et al. 2010, 2011). Neurochemical changes may provide early indication of future structural and functional damage to the central nervous system and thus monitoring neurochemical changes represents a potential risk assessment tool in ecotoxicology (Adams et al. 2010; Basu et al. 2005, 2007; Gagné et al. 2007; Nam et al. 2010). In addition, despite experimental evidence that selenium (Se) may afford protection against the neurotoxicity of MeHg, the possible modulating effects of Se on MeHg-induced neurochemical changes in free-ranging wildlife are poorly understood (Scheuhammer et al. 2008). Thus, we undertook the present study to assess Hg and Se concentrations, and various neurochemical parameters, in an opportunistically collected sample of common loons found dead of botulism on the north shore of Lake Erie, Ontario, Canada.
Methods
Carcasses of common loons (n = 68) dying of botulism were collected opportunistically during fall migration in November 2005, near Long Point, Ontario, Canada, shipped frozen, using storage and handling procedures as previously described (Stamler et al. 2005), to the National Wildlife Research Centre (NWRC), Environment Canada, Ottawa, Canada, and stored frozen until analysis. Livers (n = 68) were excised, and based on a range of THg concentrations analyzed in liver tissue, a subset of loons were chosen for brain excision (n = 41; 20 female and 21 male). Brains were dissected into 3 separate regions-brain stem, cerebellum, and cortex—and were stored at −80°C in separate acid washed polyethylene vials prior to preparation for Hg, Se, and neurochemical analyses. These three brain regions were chosen based on previous wildlife studies showing them to be sensitive to MeHg (Basu et al. 2007, 2010).
Hg and Se analysis
Liver and brain samples were prepared and analyzed for THg and Se in an accredited NWRC laboratory for metals analyses using established protocols as described previously (Scheuhammer and Bond 1991; Scheuhammer et al. 1998; Neugebauer et al. 2000; Weech et al. 2004). THg concentrations were measured in dried tissue homogenates using a Direct Mercury Analyzer (DMA-80, Milestone Inc., CT). All elemental residue concentrations are reported on a dry weight basis unless otherwise indicated. Analytical accuracy and precision were monitored through the use of Standard Reference Materials (SRMs), and intermittent analysis of duplicate samples. SRMs included National Research Council of Canada (NRCC) DOLT-3 (dogfish liver), DORM-2 (dogfish muscle), and TORT-2 (lobster hepatopancreas). Average recovery of Hg and Se was within 10% of certified values for all analyses; similarly, analytical precision (% Relative Standard Deviation of replicate samples) averaged 10% for all analyses.
Neurotransmitter receptor assays
Concentrations of glutamate (N-methyl-d-aspartic acid-NMDA) and muscarinic acetylcholine (mACh) neuroreceptors were determined in each brain region. Cellular membranes were prepared from brain tissue using protocols described in detail elsewhere (Scheuhammer et al. 2008; Nam et al. 2010). For mACh receptor assays, a Na/K buffer was used (50 mm NaH2PO4, 5 mm KCl, 120 mm NaCl, pH 7.4) and for NMDA receptors, a Tris buffer was employed (50 mm Tris, 100 μm glycine, 100 μm l-glutamic acid, pH 7.4). 30 μg of prepared membrane was re-suspended in the appropriate buffer and added to microplate wells containing a 1.0 μm GF/B glass filter (Millipore, Boston, MA, USA). For mACh receptor binding, samples were incubated with 1 nm [3H]-QNB (42 Ci mmol−1; NEN/Perkin Elmer, Boston, MA, USA) for 60 min. For NMDA receptor binding, samples were incubated with 5 nm [3H]-MK-801 (22 Ci mmol−1; NEN/Perkin Elmer) for 120 min. All assays were carried out on a shaking platform at room temperature. Binding reactions were terminated by vacuum filtration. The filters were rinsed three times with buffer and then allowed to soak for 96 h in 25 μl of OptiPhase Supermix Cocktail (Perkin Elmer). Radioactivity retained by the filter was quantified by liquid scintillation counting in a microplate detector (Wallac Microbeta, Perkin Elmer) having a counting efficiency of approximately 35%. Specific binding to both receptors was defined as the difference in radioligand bound in the presence and absence of 100 μM unlabelled atropine (for mACh receptors) and MK-801 (for NMDA receptors). Binding was reported as fmol of radioisotope bound per mg of membrane protein (fmol/mg). All samples were assayed in quadruplicate for total and non-specific binding. Intra- and inter-plate variation in binding was less than 10% as determined by use of internal, pooled controls.
Brain enzyme analysis
The activities of cholinesterase (ChE) and monamine oxidase (MAO) were determined as previously described (Scheuhammer et al. 2008; Nam et al. 2010). Briefly, tissues were sonicated for 30 s in cold Na/K buffer including 0.5% (v/v) Triton X-100. Following a 10 min centrifugation at 15,000g (4°C), the supernatant was removed. For ChE activity, 0.5 μg of supernatant protein was mixed with 100 μM 10-acetyl-3,7-dihydroxyphenoxazine, 200 mU horseradish peroxidase, 20 mU choline oxidase, and 100 μM acetylcholine. For MAO activity, 5 μg of protein was mixed with 100 μM 10-acetyl-3,7-dihydroxyphenoxazine, 200 mU horseradish peroxidase, and 100 mM tyramine. Following a 30 min incubation period for assays, the reaction end-product, resorufin (ex = 540, em = 590), was monitored between 30 and 60 min (CytoFluor 2350, Millipore, Bedford, MA, USA). Specific activities of ChE and MAO were expressed as nmol resorufin formed per min per unit (μg or mg) protein. Each sample was assayed in triplicate. Intra- and inter-plate variation in binding was less than 7% as determined by use of internal standards.
Statistical analyses
For all statistical comparisons, the critical level of significance was set at α = 0.05. THg results are reported as mean ± standard deviation. Prior to statistical analysis, brain and liver THg levels were log-transformed to achieve a normal distribution (determined using the method of D’Agostino et al. (1990). One-way ANOVAs were used to test whether concentrations of THg, Se, or neurochemical biomarkers varied significantly across the three brain regions, except for NMDA and MAO for which the non parametric Kruskal–Wallis test was used as log transformation was unable to normalize these neurochemical data. Standard linear correlation analyses were used to test whether there were significant associations between THg and any of the neurochemical variables, Standard linear correlation analyses were also used to test whether THg in liver was significantly correlated with THg in any of the three brain regions; and also to test if THg and Se were significantly correlated in any of the three brain regions. All statistical analyses were performed using SAS (Version 9.1, Institute, Cary, NC, USA).
Results
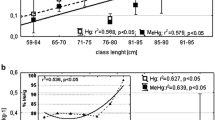
THg in common loon liver tissue ranged from 1.17 to 77.2 μg/g, with a mean of 22.8 μg/g (Table 1). The brain regions revealed substantially lower THg concentrations than the liver, ranging from 0.27 to 3.43 μg/g in the brain stem, 0.32–2.83 μg/g in the cerebellum, and 0.37–3.65 μg/g in the cortex, with overall means of 1.16, 1.17, and 1.38 μg/g, respectively (Table 1). There were significant differences in THg between the three regions (Table 1), determined by ANOVAs performed on log-transformed data, with THg levels in the cortex significantly higher than the two other regions. Concentrations of liver THg were positively correlated with THg in each of the brain regions, with the strongest association found between liver and cortex (Fig. 1).

Relationships between THg in liver and THg in three brain regions [Cortex (black circle); Cerebellum (white circle); Brain stem (black triangle)] from common loons found dead of botulism. Dashed lines represent best-fit linear regressions. Linear correlation coefficients (r), slopes, and y-intercepts for log-transformed data are similar for the 3 brain regions. Cortex: r = 0.704 (p < 0.05), slope = 0.32, intercept = −0.28; Cerebellum: r = 0.657 (p < 0.05), slope = 0.28, intercept = −0.30; brain stem: r = 0.668 (p < 0.05); slope = 0.34; intercept = −0.40
Se levels ranged from 1.47 to 28.5 μg/g in the cortex; values were lower in the cerebellum but not to a level of statistical significance (Table 1). When compared to THg values from the same brain region, there were no significant correlations between Hg and Se. The molar ratio of Hg:Se ranged from 0.02 to 0.54 in the cerebellum, and 0.02–0.58 in the cortex, with overall mean Hg:Se molar ratio of 0.14 and 0.15, respectively (Table 1), indicating a molar excess of Se over Hg in all samples. Significant positive correlations were found in both regions between the Hg:Se molar ratios and THg (p > 0.05; Table 2 and Fig. 2), with a stronger association found in the cortex.

Relationships between THg and Hg:Se molar ratio in Cortex (black circle) and Cerebellum (white circle) from common loons found dead of botulism. Dashed lines represent best-fit linear regressions. Linear correlation coefficients (r), slopes, and y-intercepts for log-transformed data are similar for both brain regions. Cortex: r = 0.781 (p < 0.05), slope = 1.14, intercept = −1.05; Cerebellum: r = 0.632 (p < 0.05), slope = 1.09, intercept = −0.98
Four neurochemical biomarkers were measured in each of the three brain regions. For each of the biomarkers, significant differences in mean values were found across brain regions with levels generally greatest in the cerebral cortex (Table 1). However, no significant correlations were found between brain THg and neurochemical biomarkers in any of the three brain regions (Table 2). For the NMDA receptor, negative trends were found with THg in each of the three brain regions, but not to a level of statistical significance.
Discussion
As a piscivorous species with a wide geographic range, the common loon has been proposed as a useful bioindicator of dietary MeHg availability in aquatic environments across North America (Evers 2006). Studies on wild loons have documented associations between MeHg exposure and behavioral alterations resulting in reduced reproductive capacity and altered parent–chick relationships (Nocera and Taylor 1998; Evers et al. 2008). A study by Evers et al. (2008), based on an extensive 18-years dataset of over 5,000 measurements, documented lethargy, altered reproductive behavior, and wing area asymmetry in loons related to MeHg exposure. Laboratory studies have confirmed some of these field observations, and have documented MeHg-associated oxidative stress (Kenow et al. 2008), immunosupression (Kenow et al. 2007), and changes in loon behavior (Kenow et al. 2010). Collectively, studies on wild and captive loons suggest that loons may be at particularly high risk for MeHg toxicity. Barr (1986) and Burgess and Meyer (2008) concluded that reproductive success in free-living loons was reduced by 50% when birds feed on fish with 0.2–0.3 μg/g Hg (wet wt.), and virtually no reproductive success was observed when birds feed on fish with 0.3–0.4 μg/g Hg (wet wt.), concentrations that are frequently observed in several fish species across eastern North America (Kamman et al. 2005).
Because common loons can be exposed to relatively elevated dietary concentrations of MeHg, methods are needed to determine levels of exposure that are associated with early and sub-clinical health effects (i.e., effects that precede overt gross toxicity). Neurochemical biomarkers may serve this purpose. Studies on fish and aquatic invertebrates (Gagné et al. 2007; Adams et al. 2010; Nam et al. 2010) and some fish-eating mammals (Basu et al. 2005; Basu et al. 2007) have shown that contaminant-associated neurochemical changes precede functional and structural damage to the brain. Similarly, Scheuhammer et al. (2008) documented Hg-associated neurochemical changes in wild common loons and bald eagles found dead in various locations across Canada; specifically, brain Hg levels were positively correlated with mACh receptors and negatively correlated with NMDA receptors. One limitation of the Scheuhammer et al. (2008) study was that Hg accumulation and neurochemical changes were not characterized in distinct brain regions, rather they were measured in the whole brains. The brain is perhaps the most heterogeneous organ in higher vertebrates and resolving Hg accumulation and associated changes in distinct anatomical regions may increase our understanding of the causative linkages between Hg concentrations, neurochemical changes, and observed behavioral changes. Accordingly, a major goal of the present study was to characterize THg levels and selected neurochemical parameters in three discrete brain regions previously shown to be affected by Hg in wildlife (Basu et al. 2007, 2010). In the current study, Hg concentrations varied across three common loon brain regions and were highest in the cerebral cortex. The observed range of brain Hg concentrations (0.3–3.7 μg/g across three brain regions) was much narrower than noted in a previous study on common loons found dead in Canada, in which whole brain Hg values ranged from 0.2 to 68 μg/g (Scheuhammer et al. 2008). The values reported in the present study, however, were similar to values reported for Hg in brains of Great Lakes herring gulls, and for some other aquatic bird species from non-contaminated sites in North America (Rutkiewicz et al. 2010).
In the current study, no significant Hg-associated changes in neurochemical biomarkers were observed. Similarly, Rutkiewicz et al. (2010) noted no significant associations between brain Hg and neurochemical markers in herring gulls; nor in bald eagles with relatively low concentrations of brain Hg (Rutkiewicz et al. 2011). We hypothesize that brain THg levels in these studies were probably too low to be associated with significant neurochemical change. The maximum loon brain Hg value measured in the current study is nearly 20-times lower than the maximum value reported by Scheuhammer et al. (2008) in which significant Hg-related neurochemical changes were found in loons. Taken together, these studies suggest that brain Hg concentrations <3 μg/g (dry wt.) may not be sufficiently high to produce consistent changes in brain neurochemistry in free-living avian species. In addition, loon brain samples analyzed in the current study for both THg and Se showed molar excesses of Se over Hg, and also exhibited significant positive correlations between Hg:Se and THg (Fig. 2). Excess molar concentrations of Se are generally thought to be protective against Hg toxicity (Ralston et al. 2008). The relatively low brain Hg concentrations that we observed coupled with molar excesses of Se in the same brain samples are consistent with a lack of Hg-related neurochemical effects.
Though a Hg-associated neurochemical effect threshold has not been established for wildlife, previous studies fish-eating mammals (Basu et al. 2006, 2007) and birds (Scheuhammer et al. 2008) have suggested that neurochemical changes occur in the 3–5 ppm brain Hg range. Also, Hoffman et al. (2011) reported that brain THg concentrations averaging about 3 μg/g (dry wt.) in breeding adult Forster’s terns (Sterna forsteri) were associated with evidence of increased lipid peroxidation, loss of protein bound thiols, and decreased activity of antioxidant enzymes. Effects thresholds may vary between bird species, and also between mammals and birds. In a study on polar bear brain stem tissues, Hg-associated decreases in NMDA receptors were reported despite brain Hg levels being less than 1 μg/g (Basu et al. 2009). In a study of laboratory mink fed MeHg, a Hg-associated decrease in NMDA receptor levels was found in the lowest exposure group (0.1 μg/g in diet resulting in brain Hg levels of 1–2.2 μg/g) (Basu et al. 2007). Although significant associations were not observed in the current study, a weak negative trend was observed between brain Hg levels and NMDA receptor levels. Results of the present study are valuable to help determine threshold tissue-Hg concentrations that are associated with significant changes in important neurochemical signaling pathways in MeHg-exposed wildlife.
References
Adams DH, Sonne C, Basu N, Dietz R, Nam DH, Leifsson PS, Jensen AL (2010) Mercury contamination in spotted seatrout, Cynoscion nebulosus: an assessment of liver, kidney, blood, and nervous system health. Sci Total Environ 408:5808–5816
Albers PH, Koterba MT, Rossman R, Link WA, French JB, Bennett R, Bauer WC (2007) Effects of methylmercury on reproduction in American kestrels. Environ Toxicol and Chem 26:1856–1866
Barr JF (1986) Population dynamics of the common loon (Gavia immer) associated with mercury-contaminated waters in north-western Ontario. Can. Wildl Ser Occasional Paper 56:1–25
Basu N, Klenavic K, Gamberg M, O’Brien M, Evans RD, Scheuhammer AM, Chan HM (2005) Effects of mercury on neurochemical receptor binding characteristics in wild mink. Environ Toxicol Chem 24:1444–1450
Basu N, Scheuhammer AM, Rouvinen-Watt K, Grochowina N, Klenavic K, Evans RD, Chan HM (2006) Methylmercury impairs components of the cholinergic system in captive mink (Mustela vison). Toxicol Sci 91:202–209
Basu N, Scheuhammer AM, Rouvinen-Watt K, Grochowina NM, Evans RD, O’Brien M, Chan HM (2007) Decreased N-methyl-d-aspartic acid (NMDA) receptor levels are associated with mercury exposure in wild and captive mink. Neurotoxicology 28:587–593
Basu N, Scheuhammer AM, Sonne C, Letcher RJ, Born EW, Dietz R (2009) Is dietary mercury of neurotoxicological concern to polar bears (Ursus maritimus)? Environ Toxicol Chem 28:133–140
Basu N, Scheuhammer AM, Rouvinen-Watt K, Evans RD, Trudeau VL, Chan HM (2010) In vitro and whole animal evidence that methylmercury disrupts GABAergic systems in discrete brain regions in captive mink. Comp Biochem Physiol Part C 151:379–385
Bennett RS, French JB, Rossmann R, Haebler R (2009) Dietary toxicity and accumulation of methylmercury in American kestrels. Arch Environ Contam Toxicol 56:149–156
Burgess NM, Meyer MW (2008) Methylmercury exposure associated with reduced productivity in common loons. Ecotoxicology 17:83–91
D’Agostino RB, Belanger A, D’Agostino RB Jr (1990) A suggestion for using powerful and informative tests of normality. Am Stat 44:316–321
Evers DC (2006) Loons as biosentinels of aquatic integrity. Environ Bioindicators 1:18–21
Evers DC, Savoy LJ, DeSorbo CR, Yates DE, Hanson W, Taylor KM, Siegel LS, Cooley JH Jr, Bank MS, Major A, Munney K, Mower BF, Vogel HS, Schoch N, Pokras M, Goodale MW, Fair J (2008) Adverse effects from environmental mercury loads on breeding common loons. Ecotoxicology 17:69–81
Gagné F, Cejka P, André C, Hausler R, Blaise C (2007) Neurotoxicological effects of a primary and ozonated treated wastewater on freshwater mussels exposed to an experimental flow-through system. Comp Biochem Physiol C Toxicol Pharmacol 146:460–470
Hoffman DJ, Eagles-Smith CA, Ackerman JT, Adelsbach TL, Stebbins KR (2011) Oxidative stress response of Forster’s terns (Sterna forsteri) and Caspian terns (Hydroprogne caspia) to mercury and selenium bioaccumulation in liver, kidney, and brain. Environ Toxicol Chem 30:920–929
Kamman NC, Burgess NM, Driscoll CT, Simonin HA, Goodale W, Linehan J, Estabrook R, Hutcheson M, Major A, Scheuhammer AM, Scruton DA (2005) Mercury in freshwater fish of northeast North America: A geographic perspective based on fish tissue monitoring databases. Ecotoxicology 14(1–2):163–180
Kenow KP, Grasman KA, Hines RK, Meyer MW, Gendron-Fitzpatrick A, Spalding MG, Gray BR (2007) Effects of methylmercury exposure on the immune function of juvenile common loons (Gavia immer). Environ Toxicol Chem 26:1460–1469
Kenow KP, Hoffman DJ, Hines RK, Meyer MW, Bickham JW, Matson CW, Stebbins KR, Montagna P, Elfessi A (2008) Effects of methylmercury exposure on glutathione metabolism, oxidative stress, and chromosomal damage in captive-reared common loon (Gavia immer) chicks. Environ Pollut 156:732–738
Kenow KP, Hines RK, Meyer MW, Suarez SA, Gray BR (2010) Effects of methylmercury exposure on the behavior of captive-reared common loon (Gavia immer) chicks. Ecotoxicology 19:933–944
Nam DH, Adams DH, Flewelling LJ, Basu N (2010) Neurochemical alterations in lemon shark (Negaprion brevirostris) brains in association with brevetoxin exposure. Aquat Toxicol 99:351–359
Neugebauer EA, Sans Cartier GL, Wakeford BJ (2000) Methods for the determination of metals in wildlife tissues using various atomic absorption spectrophotometry techniques. Technical Report Series No. 337E. Canadian Wildlife Service, Environment Canada, Ottawa, Canada
Nocera J, Taylor P (1998) In situ behavioural response of common loons associated with elevated mercury exposure. Conserv Ecol 2:10
Ralston NV, Ralston CR, Blackwell JL, Raymond LJ (2008) Dietary and tissue selenium in relation to methylmercury toxicity. Neurotoxicology 29:802–811
Rutkiewicz J, Crump D, Scheuhammer AM, Jagla M, Basu N (2010) Investigation of spatial trends and neurochemical impacts of mercury in herring gulls across the laurentian great lakes. Environ Pollut 158:2733–2737
Rutkiewicz J, Nam D-H, Cooley T, Neumann K, Padilla IB, Route W, Strom S, Basu N (2011) Mercury exposure and neurochemical impacts in bald eagles across several Great Lakes states. Ecotoxicology. doi:10.1007/s10646-011-0730-1
Scheuhammer AM, Bond D (1991) Factors affecting the determination of total mercury in biological samples by continuous-flow cold vapour atomic absorption spectrophotmetry. Biol Trace Elem Res 31:119–129
Scheuhammer AM, Wong AHK, Bond D (1998) Mercury and selenium accumulation in common loons (Gavia immer) and common mergansers (Mergus merganser) from Eastern Canada. Environ Toxicol Chem 17:197–201
Scheuhammer AM, Meyer MW, Sandheinrich MB, Murray MW (2007) Effects of environmental methylmercury on the health of wild birds, mammals, and fish. Ambio 36:12–18
Scheuhammer AM, Basu N, Burgess N, Elliott JE, Campbell GD, Wayland M, Champoux L, Rodrigue J (2008) Relationships among mercury, selenium, and neurochemical parameters in common loons (Gavia immer) and bald eagles (Haliaeetus leucocephalus). Ecotoxicology 17:93–101
Spalding GM, Frederick PC, McGill HC, Bouton SN, Richey LJ, Schumacher IM, Blackmore CGM, Harrison J (2000) Histologic, neurologic, and immunologic effects of methylmercury in captive great egrets. J Wild Dis 36:423–435
Stamler CJ, Basu N, Chan HM (2005) Biochemical markers of neurotoxicity in wildlife and human populations: considerations for method development. J Toxicol Environ Health A 68(16):1413–1429
Weech SA, Scheuhammer AM, Elliot JE, Cheng KM (2004) Mercury in fish from the Pinchi Lake region, British Columbia, Canada. Environ Pollut 131:275–286
Acknowledgments
The authors thank Della Bond and Emily Porter for their excellent technical assistance. Danny Bernard, Paul Ashley, Jeff Robinson, Kerrie Wilcox, Matt Timpf and Craig Hebert contributed to the collection and dissection of the loons used in this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hamilton, M., Scheuhammer, A. & Basu, N. Mercury, selenium and neurochemical biomarkers in different brain regions of migrating common loons from Lake Erie, Canada. Ecotoxicology 20, 1677–1683 (2011). https://doi.org/10.1007/s10646-011-0754-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-011-0754-6