
Abstract
Temporal patterns in otolith Sr:Ca and Ba:Ca ratio values of American eels Anguilla rostrata from two sites in western Newfoundland gave insight into the use of freshwater and saline habitats. Mean Sr:Ca and Ba:Ca values at the core zone did not differ between sites, indicative of a common oceanic origin. At the otolith edge, representing continental life, both Sr:Ca and Ba:Ca values varied between sites consistent with ambient element:Ca ratio values and salinity, with typically higher Sr:Ca and lower Ba:Ca values in saline than in fresh waters. Most eels (73%) from Muddy Hole, an estuarine site, were evaluated as estuarine residents while most (70%) eels from Castors River, a freshwater site, were evaluated as freshwater residents, with the remaining eels from each site evaluated as inter-habitat migrants. An otolith element:Ca critical value appropriate for distinguishing between fresh and saline water residence is fundamental for estimating the proportion of eel residence in freshwater and their subsequent classification into habitat residence groups. Such classification is moderately robust to the critical value selected. For inter-habitat migrants, moderate otolith Sr:Ca values between the elver check and otolith edge suggestive of estuarine residence may coincide with Ba:Ca values suggestive of freshwater residence. No general critical value for separating fresh and estuarine habitats was found for otolith Ba:Ca. Otolith Ba:Ca temporal patterns may assist the use of Sr:Ca in the evaluation of historical habitat residence and inter-habitat movement but the use of otolith Ba:Ca values should be applied cautiously for American eels and perhaps of other estuarine/freshwater migratory fishes.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
American eel Anguilla rostrata stocks have declined in many regions of North America (Jessop 1997; Haro et al. 2000). The species has been designated as of special concern in Canada by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC 2006) and its status has been reviewed by the U.S. Fish and Wildlife Service (USFWS 2005) but has not been placed under protection in Canada or the United States (USFWS 2007). The complex life history and wide geographic range of the American eel makes the management of the population and of specific fisheries difficult. The facultative catadromy of the species (and of the European A. anguilla and Japanese A. japonica) means that varying portions of local and regional stocks may occupy freshwater, estuarine, or marine habitats and engage in a degree of inter-habitat migration (reviewed by Daverat et al. 2006; Jessop et al. 2008).
Chronological changes in the ratios of element:calcium composition in the otolith e.g., Sr:Ca and Ba:Ca along a growth axis have been used to reconstruct the migratory history of diadromous and estuarine fishes (reviewed by Elsdon et al. 2008). The ionic radii of divalent metals such as Sr and Ba may substitute for Ca in the orthorhombic aragonite lattice of fish otoliths and element incorporation depends on ambient element concentration (Campana 1999). Although the absolute concentration of Sr is about 100× greater in marine than in fresh waters, the approximately 4.8× greater difference in the molar ratio of Sr:Ca between salt and fresh water is more relevant to the relative degree of Sr biological uptake between salt and fresh water (Campana 1999).
Positive relations varying from linear (Elsdon and Gillanders 2005a,b; Walther and Thorrold 2006; Bradbury et al. 2008; MacDonald and Crook 2010) to curvilinear (Miller et al. 2010) have been found between otolith Sr:Ca and ambient concentrations, although no relation between otolith and ambient Sr:Ca was found by Dorval et al. (2007) over a limited ambient Sr:Ca range. For Ba:Ca, the positive relation between otolith and ambient concentrations may vary from linear (Bath et al. 2000; Elsdon and Gillanders 2005a; Bradbury et al. 2008; Miller et al. 2010) to non-linear (logarithmic or exponential) relations (Elsdon and Gillanders 2005a,b; Dorval et al. 2007; Miller 2009; Macdonald and Crook 2010; Miller et al. 2010), with little consistency between laboratory and field settings. Variable relations between ambient element:Ca occur with salinity—positive and linear for Sr:Ca (Tzeng 1996; Kawakami et al. 1998; Secor and Rooker 2000), exponential increase (Macdonald and Crook 2010), or no correlation over limited salinity ranges (Elsdon and Gillanders 2005a; Dorval et al. 2007) and negative linear for Ba:Ca (Elsdon and Gillanders 2005b; Dorval et al. 2007; Bradbury et al. 2008) or non-linear (logarithmic, exponential) (Elsdon and Gillanders 2005a; Martin and Wuenschel 2006; Macdonald and Crook 2010). The interaction of ambient element:Ca and salinity results in otolith element:Ca relations with salinity that may exponentially increase (Macdonald and Crook 2010) for Sr and decrease linearly (Dorval et al. 2007) or exponentially (Elsdon and Gillanders 2005a; Macdonald and Crook 2010) for Ba. Both otolith Sr:Ca and Ba:Ca may show no relation with salinity over a limited salinity range (Dorval et al. 2007; Martin and Wuenschel 2006). Ambient concentrations of dissolved Ba vary in a more complex manner with salinity than does Sr (Coffey et al. 1997). The non-linear relations between otolith Sr:Ca (positive) and Ba:Ca (negative) and ambient salinity makes the reconstruction of historical habitat salinities more applicable to broad classifications such as fresh, estuarine, and saltwater than to detailed salinity levels (Kraus and Secor 2004; Elsdon and Gillanders 2005a; Zimmerman 2005).
Water temperature may also influence element incorporation into otoliths, generally positively for Sr but variably for Ba, and may differ among species (Elsdon and Gillanders 2002, 2004; Elsdon et al. 2008; Miller 2009) as well as acting interactively with ambient element concentration and salinity. However, the effect of temperature is about an order of magnitude less than that due to diadromy (Campana 1999) and may reflect water chemistry more than temperature (Kraus and Secor 2004). Although Kalish (1989) found a positive relation between growth rate and otolith Sr:Ca, Bath et al. (2000) concluded that the incorporation of Sr and Ba in otoliths is uncorrelated with fish growth rate.
Inter-habitat movements by anguillid eels have typically been examined via temporal patterns in otolith Sr:Ca (reviewed by Daverat et al. 2006; Jessop et al. 2008) and, less commonly, Ba:Ca (Arai and Hirata 2006; Tabouret et al. 2010). Ideally, the use of Ba:Ca data to assess fish habitat residence and inter-habitat movement should provide categorizations consistent with the use of Sr:Ca data. This study compares life-history patterns in otolith Sr:Ca and Ba:Ca and bivariate plots of element:Ca data to assess habitat residence group (freshwater, estuarine/marine, and inter-habitat migrant), evaluates the effects of different Sr:Ca critical values on the estimation of the percentage of freshwater residence, and uses Sr:Ca data and its critical value to estimate a putative Ba:Ca critical value so as to evaluate whether Sr:Ca and Ba:Ca are complimentary for evaluating estuarine habitat residence for American eels Anguilla rostrata from a freshwater and an estuarine site.
Materials and methods
Study sites
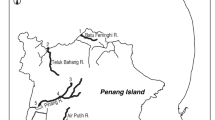
Muddy Hole (MH), a cove at the mouth of Flat Bay Brook (48° 24′ 27″ N, −58° 34′ 41″ W) and Castors River (CR) (50° 55′ 15″ N, −56° 57′ 07″ W) are on the western coast of Newfoundland (Fig. 1) and have watershed areas of 635.2 km2 and 544.2 km2, respectively. Single, surface water samples were collected in October 2005 from Muddy Hole and Castors River and later laboratory analyzed. Muddy Hole, a tidal, estuarine cove of about 17 ha in area at the mouth of Flat Bay Brook that may become entirely freshwater at high discharges from Flat Bay Brook, had a pH of 6.8 and conductivity of 435 μS·cm−1, equivalent to a salinity of 0.20 (Table 1). Castors River, at a site about 0.4 km upstream from the limit of saltwater penetration, had a pH of 7.4, conductivity of 179 μS·cm−1, and salinity of 0.09. Water samples taken in the early 1970s indicate that Flat Bay Brook had a pH of 6.5, standard conductivity of 22 μS·cm−1, and total alkalinity of 4.0 ppm while Castors River had a pH of 7.5, conductivity of 71 μS·cm−1, and alkalinity of 32.0 ppm (Porter et al. 1974). Elemental concentrations of strontium (Sr), barium (Ba), and calcium (Ca) were measured in the water samples from each study site by ICP-MS (Maxxam Analytics Inc., Halifax, NS) and expressed as element:Ca ratios expressed as mg·g−1 for Sr:Ca and μg·g−1 for Ba:Ca; Table 1) because Sr and Ba in otoliths vary relative to element:Ca ratios rather than absolute concentration (Bath et al. 2000; Kraus and Secor 2004).

Map of the mouths of the Castors River and Muddy Hole, Flat Bay Brook in western Newfoundland
Eel collection and measurement
American eels were collected from commercial fyke net fisheries at Muddy Hole (N = 57) on 28 September 2005 and at Castors River on 16 October 2005 (N = 10) and 24 October 2006 (N = 49) at a site about 11 km upstream from the river mouth. Total lengths (to 1.0 mm) and total body weights (to 0.1 g) were measured fresh. Most eels from the Castors River were silver phase while most from Muddy Hole were yellow phase. Jessop et al. (2009) provides additional details of the sampling, processing, and biological characteristics of the eels.
Otolith ageing and microchemistry analysis
Sagittal otoliths for ageing and microchemical analysis were extracted from 11 randomly selected eels from Muddy Hole and 10 eels from Castors River sampled in 2005. One otolith per pair was prepared for analysis by grinding and polishing along the sagittal plane until the primordium was exposed. Polished otoliths were decontaminated by immersion in 5% ultrapure hydrogen peroxide (H2O2) for 10 min to oxidize all remaining organic material from the otolith surface, ultrasonically cleaned three times, each for 5 min, with de-ionised water and then air-dried. Otolith elements were measured along the growth axis from the primordium to the otolith edge with a Merchantek LUV 266 Nd: YAG UV laser microprobe (New Wave Research, Inc) connected to a Finnigan MAT ELEMENT 2 high resolution inductively coupled plasma mass spectrometer (ICP-MS) (Thermo Electron corp., Bremen, Germany). The otoliths were placed in a sealed chamber and viewed through a microscope connected to a computer monitor to program the analyzing transects. The laser was pulsed at a repetition rate of 20 Hz at a scan speed of 15 μm·sec−1 with an ablation beam diameter of approximately 150 μm and a dwell time of 35 ms and power of 3.5 J·cm2. For all eels from Castors River and for three eels from Muddy Hole, each data measurement represented a progressive otolith transect scan of 1.60 s while a scan of 2.13 s was made for the remaining seven eels from Muddy Hole. The beam diameter and scan distance were set originally so as to provide adequate measurements for six otolith elements. The elements measured and reported here (44Ca, 88Sr, and 138Ba) remained constantly at least 10 times higher than background levels. Detection limits were 840.0 μg·g−1 for Ca, 1.08 μg·g−1 for Sr, and 0.21 μg·g−1 for Ba. Standards (NIST 612) were collected before each series, with each series comprising 2–3 otoliths. At the start of each otolith analysis, background counts were collected for 30 s, and the average was subtracted from sample counts to correct for background levels. The ablation chamber was purged for 60 s after sampling each otolith. Calcium was used as the internal standard with an otolith concentration of 40%wt (400 000 μg Ca g−1 otolith). All measurements were expressed as wt% ratios of element to Ca concentrations (mg·g−1, equivalent to × 10−3, for Sr:Ca and μg·g−1, equivalent to × 10−6, for Ba:Ca) by estimating the relative response factor of the instrument to the known concentration in the standard (NIST 612). After laser ablation ICP-MS analysis, the otoliths were etched in 5% EDTA to enhance the annuli for ageing (Tzeng et al. 1994).
Data analysis
The environmental history of each eel was interpreted by examining the temporal pattern of Sr:Ca and Ba:Ca values along an otolith transect from primordium to edge, with the assumption that the temporal pattern reflects habitat salinity due to the positive relation between otolith Sr:Ca value and ambient salinity (Tzeng 1996; Kawakami et al. 1998; Secor and Rooker 2000). Jessop et al. (2002) considered ratio values of Sr:Ca ≤4 × 10−3 between the elver check and the otolith edge as equivalent to freshwater residence for American eels while Thibault et al. (2007) considered the critical value to be between 3.5–4.0 × 10−3. For European eels, the freshwater critical value has ranged from <0.4 × 10−3 (Tzeng et al. 1997) to <2.0–2.24 × 10−3 (Daverat and Tomás 2006; Shiao et al. 2006; Tabouret et al. 2010) and for Japanese eels from <2.5 × 10−3 (Tsukamoto and Arai 2001; Kotake et al. 2005) to <4 × 10−3 (Tzeng et al. 2002). The grand mean (± SD) Sr:Ca values between the elver check and otolith edge for 7 eels from Castors River considered to be freshwater residents from their transect profile was 0.96 (± 0.75) × 10−3 (means range 0.48–1.74 × 10−3). A conservative Sr:Ca critical value of 3.5 × 10−3 (rounded up from 3.2 × 10−3) was chosen based upon the grand mean + 3 SD.
Based upon this critical value, eels were classified into fresh and salt (estuarine/marine) water residence groups and an inter-habitat migrant group when Sr:Ca values along the otolith transect varied across the critical value, with freshwater being ≤3.5 × 10−3 and saline water being >3.5 × 10−3. Coastal and marine, or simply seawater residence, with no defining salinity, has been defined for European eels at otolith Sr:Ca values >6.0 × 10−3 (Daverat et al. 2005, 2006; Daverat and Tomás 2006), and as >4.9 × 10−3 for a salinity >20 (Tabouret et al. 2010) and as >5.0 or 6.0 × 10−3 for Japanese eels (Tsukamoto and Arai 2001; Kotake et al. 2005; Arai and Hirata 2006), with values intermediate between freshwater and seawater being termed estuarine. If the freshwater zone is defined as having a salinity ≤0.5, estuarine zone as >0.5 to 30, and the marine zone as >30 (Venice System; Bulger et al. 1993), then defining coastal/marine residence as being represented by otolith Sr:Ca values >6.0 × 10−3 is plausible. Estuarine habitat residence has thus been inferred from otolith Sr:Ca values >3.5 and <6.0 × 10−3. A more biologically relevant salinity boundary for coastal/marine residence might be >24 (Bulger et al. 1993), with consequent reduction in the critical otolith Sr:Ca value for the estuarine to marine transition. Otolith Sr:Ca critical values for the broad classifications of freshwater, estuarine, and marine clearly vary among species and sites.
Otolith Ba:Ca values were assumed to decrease from fresh to brackish to seawater because of the decrease in otolith Ba:Ca value with increasing environmental salinity (Elsdon and Gillanders 2005a, b; Dorval et al. 2007; Macdonald and Crook 2010), because salinity tends to increase with distance downstream in an estuary, and because Ba is of terrestrial origin (Coffey et al. 1997).
The effects of habitat residence (freshwater, estuarine, migrant) on otolith element:Ca values between the elver check and otolith edge was examined by a linear mixed effects model using package lme4 (R Core Development Team 2010), with habitat group as a fixed effect and fish as a random effect, followed by a Tukey multiple comparison test of specific contrasts. Autocorrelation among adjacent element:Ca values was accounted for by adjusting for a first order autocorrelation within each fish.
The effects of ontogeny and site on otolith element:Ca values were examined by a linear mixed effects model, with site and otolith zone considered as fixed effects and fish and fish × otolith zone interaction considered as random effects followed by a Tukey multiple comparison test of specific contrasts. The mean element:Ca values were based on the first three or four ablation spots, depending upon whether the spot scan length was 32 or 24 μm (96 μm length in total), beginning at the core (primordium) and elver check and before the otolith edge (core, elver check, and edge zones). Autocorrelation was low and of no concern. The larval phase of the eel life history is represented by the core zone, the newly-arrived glass eel/elver in continental waters by the elver check zone, and the continental juvenile stage at capture by the edge zone. The 3–4 element:Ca spots comprising 96 μm on the otolith transect represent about 1 year in the elver check zone (mean distance between elver check and first annulus = 106 μm) and about 1 year at the otolith edge zone for eels of age 13–14 and 2 years for ages 17–28 years. Logarithmic (base 10) transformation of the element:Ca data was required to satisfactorily meet the normality of distribution and homogeneity of variance requirements of parametric statistics.
The mean Sr:Ca and Ba:Ca values in the zone between the elver check and the otolith edge and the mean values for the elver check zone and otolith edge zone were plotted to examine the relationship between these two parameters for insights into the habitat use and inter-habitat movements of American eels. Correlations between the mean Sr:Ca values between the elver check and otolith edge and fish age and their significance were adjusted for low sample size bias (Olkin and Pratt 1958; Zar 1984).
The percentage (proportion) of residence in freshwater (%fwr) was estimated for each fish based on the percentage of Sr:Ca values between the elver check and otolith edge based on an Sr:Ca critical value ≤3.5 × 10−3 separating freshwater from estuarine/marine residence and is effectively equivalent to the proportion of time resident in fresh water (Jessop et al. 2007). The effect of different (2.5, 3.0, 3.5, 4.0, 4.5 × 10−3) Sr:Ca critical values on the estimation of %fwr and the categorization of fish into habitat residence group was evaluated by analysis of the mean difference in %fwr estimated at each Sr:Ca critical value (with 3.5 × 10−3 as the standard) by paired sample t-test of the arcsine transform of the square root of p estimates. The Hedges’ g effect size statistic and associated 95% CI was used as a measure of the magnitude of the effect on %fwr estimates of changes in the critical value, where: \( g = {t_{{paired}}}\sqrt {{\frac{{2(1 - {r_{{12}}})}}{n}}} \) and t paired is the t value from a paired t-test, r 12 is the correlation coefficient between the two groups and n = n 1 = n 2 (Nakagawa and Cuthill 2007). Effect sizes (g) of 0.2 are considered small, 0.5 are medium, and 0.8 are large (Cohen 1988).
A review of the transect data plots and an iterative fitting process was used to establish site- and habitat group-specific Ba:Ca critical values for American eels under the assumptions that an Sr:Ca critical value of 3.5 × 10−3 reliably separated freshwater from saltwater residence and that each element:Ca relation should produce similar estimates of %fwr. The relation between Ba:Ca critical value and the estimation of %fwr for each fish from each site (MH, CR) and group (freshwater, inter-habitat migrant) was evaluated by varying the presumed Ba:Ca critical value over the range 1, 2, 3, 5, 10, 20, 30, 40, 50 × 10−6 and examining the similarity of the estimation of %fwr estimated from Ba:Ca relative to that estimated from Sr:Ca for each fish. The best Ba:Ca critical value for each fish and site × group combination was determined by the smallest difference between the resultant estimates of %fwr by Sr:Ca and Ba:Ca.
Results
Water and otolith element relations
The Sr:Ca ratio of the water sample from Muddy Hole in the estuary of Flat Bay Brook was about 5.0× greater than for the freshwater site in Castors River while the Ba:Ca rato was 3.4× greater in Castors River than in Muddy Hole (Table 1, Fig. 1). This pattern is consistent with that for the element:Ca ratios in the otolith edge, although the magnitudes differed—the otolith Sr:Ca was 2.6× higher in Muddy Hole eels than in Castors River eels while Ba:Ca was 9.7× higher in Castors River than in Muddy Hole. However, the conductivity/salinity values indicate that the surface waters at both sites were fresh (salinity <0.5) when sampled. The mean lengths, weights, and ages of the subsamples of eels used for otolith element analysis did not differ significantly (t-tests, all p > 0.50) from that of the total samples for each site (Table 2).
Habitat classification and otolith Sr:Ca and Ba:Ca
Of the 11 eels from Muddy Hole, 8 eels had mean transect values of Sr:Ca between elver check and otolith edge that averaged 5.07 × 10−3 and mean Ba:Ca values that averaged 2.17 × 10−6 (Figs. 2A, B and 4), which indicates primarily estuarine residence. Three eels showed patterns of switching from relatively high Sr:Ca and low Ba:Ca to low Sr:Ca and high Ba:Ca values, suggestive of movement between saline and fresh water habitats (Fig. 2C, D). Thus, the eel of Fig. 3C spent a short time in estuarine waters before entering freshwater and later returning to saline waters while the eel of Fig. 3D briefly entered freshwater before returning to the estuary for awhile before finally returning to freshwater.

Anguilla rostrata. Element:Ca values along a transect from core to edge for selected otoliths of American eels from Muddy Hole, western Newfoundland representing notable life history patterns. The arrow indicates the position of the elver check

Anguilla rostrata. Element:Ca values along a transect from core to edge for selected otoliths of American eels from Castors River, western Newfoundland representing notable life history patterns. The arrow indicates the position of the elver check. Fw = fresh water
Of the 10 eels from Castors River, 7 eels had mean Sr:Ca values between the elver check and otolith edge indicative of freshwater residence and higher Ba:Ca values than did estuarine residents from Muddy Hole (Figs. 3A, B and 4). Three eels had sections of otolith transect with Sr:Ca values >3.5 × 10−3, suggesting a movement from fresh to saline waters, but with Ba:Ca values mostly >15 × 10−6 even in the putatively saline portions of the transect (Fig. 3C, D) and were categorized as inter-habitat migrants. Although the Castors River eels were collected in fresh water well upstream of the river mouth, two of the inter-habitat migrants had Sr:Ca values at the otolith edge (4.2–4.7 × 10−3) suggestive of estuarine residence (Fig. 3C).

Anguilla rostrata. Mean element:Ca ratios (±SD) from elver check to otolith edge, by habitat group (Est = estuarine (n = 8 fish), Est migr = estuarine migrant (n = 3), Fw = freshwater (n = 7), Fw migr = freshwater migrant (n = 3)) and site (Muddy Hole represented by Est and Est migr; Castors River by Fw and Fw migr) for otoliths of American eels from western Newfoundland. Near each mean symbol is the mean value
Otolith element composition by site and habitat residence
Mean element:Ca values between the elver check and otolith edge differed significantly among habitat groups (Muddy Hole estuarine and inter-habitat migrants, Castors River freshwater and inter-habitat migrants: Sr:Ca—F = 58.1, df = 3, 17, p < 0.001; Ba:Ca—F = 33.9, df = 3, 17, p < 0.001) (Fig. 4). The number of element:Ca values for individual eels ranged from 32 to 62, resulting in autocorrelation estimates of 0.80 for Sr:Ca and 0.60 for Ba:Ca. Estuarine resident eels from Muddy Hole had significantly (p < 0.001) higher mean Sr:Ca values and lower (p < 0.001) Ba:Ca values than did freshwater resident eels from Castors River. Inter-habitat migrants from Muddy Hole had significantly (p = 0.033) lower Sr:Ca and higher (p < 0.001) Ba:Ca than did estuarine residents while inter-habitat migrants from Castors River had significantly (p < 0.001) higher Sr:Ca and Ba:Ca (p = 0.003) than did freshwater residents. Inter-habitat migrants from Castors River had a mean Sr:Ca that did not differ significantly (p = 0.62) from that of estuarine residents from Muddy Hole but had a higher (p < 0.001) mean Ba:Ca. Inter-habitat migrants from Muddy Hole had a mean otolith Sr:Ca significantly higher (p < 0.001) than did freshwater residents from Castors River but Ba:Ca means did not differ significantly (p = 0.63). The Akaike Information Criterion values for the models used (−1317.0 for Sr:Ca data, 304.5 for Ba:Ca data) indicated good support relative to null models that omitted the random effects and correlation structure (−698.2 for Sr:Ca data, 668.6 for Ba:Ca data).
Otolith transect (elver check to edge) mean Sr:Ca was not significantly correlated with age for freshwater resident eels from Castors River (N = 7, age range = 13–28 years, r = 0.31, p = 0.50) or for estuarine resident eels from Muddy Hole (N = 8, age range = 4–8 years, r = 0.56, p = 0.15).
Otolith element composition by site and otolith position
The mean otolith element:Ca ratios differed significantly by site and otolith position for eels from Muddy Hole and Castors River (Fig. 5). The significant interaction effect between site and otolith position (Sr:Ca: F = 49.3, df = 2, 143, p < 0.001; Ba:Ca: F = 55.4, df = 2, 143, p < 0.001) indicated that main effects should only be considered in relation to each other. Mean Sr:Ca values decreased at each site from the core to the elver check to the edge (multiple comparison tests, all p < 0.001) except for Sr:Ca values between elver check and edge for eels from Muddy Hole (p = 0.13) (Fig. 5). Mean Ba:Ca ratios varied in pattern among otolith zones, increasing significantly from core to edge for Castors River and decreasing for Muddy Hole (multiple comparison tests, all p ≤ 0.026). Neither Sr:Ca nor Ba:Ca ratios in the otolith core differed between sites (Sr:Ca: p = 0.98; Ba:Ca: p = 0.50). However, Sr:Ca values at the elver check were higher at Muddy Hole than at Castors River (p < 0.001) and Ba:Ca values were higher at Castors River than at Muddy Hole (p = 0.005). At both sites, Sr:Ca values were higher at the core than at the elver check or edge while Ba:Ca values differed in pattern among otolith zones at each site. At Muddy Hole, otolith edge Sr:Ca values were higher (p < 0.001) and Ba:Ca values were lower (p < 0.001) than at Castors River. The Akaike Information Criterion values for the models used (−144.5 for Sr:Ca data, −7.5 for Ba:Ca data) indicated good support relative to null models that omitted the random effects (0.87 for Sr:Ca data, 91.9 for Ba:Ca data).

Anguilla rostrata. Mean element:Ca ratios (+SD), by site and otolith zone, for otoliths of American eels from western Newfoundland. For each otolith zone, N = 22 at Castors River based on 7 freshwater resident eels 3 or 4 values per otolith position and N = 32 at Muddy Hole based on 8 estuarine resident eels and 4 values per otolith position
Relation between otolith Sr:Ca and Ba:Ca values
Bivariate plots of the mean otolith Sr:Ca and Ba:Ca values from the transects of 21 eels between the elver check and edge revealed variable clustering that could be related to eel residence by site, salinity of habitat (freshwater, estuarine), and stage of life cycle (early continental arrival, capture). For the elver check zone (Fig. 6A), otolith edge zone (Fig. 6B), and total transect (Fig. 6C), the element:Ca values for freshwater and estuarine resident groups were characteristically clustered but the distribution of inter-habitat migrant values varied according to their life cycle stage and particular life history e.g., short or long estuarine or freshwater residence before a habitat change. For the brackish Muddy Hole site, the transect element:Ca plot tended to form a negative curvilinear relation while for the freshwater Castors River the relation was positive curvilinear, with the inter-habitat migrant fish driving the relationship (Fig. 6C). Freshwater residents clustered at low (<2 × 10−3) mean Sr:Ca and mostly intermediate (5–15 × 10−6) Ba:Ca values, estuarine residents clustered at high (>4.5 × 10−3) Sr:Ca and low (<5 × 10−6) Ba:Ca, and inter-habitat migrants with intermediate (about 2–4 × 10−3) Sr:Ca and a wide range (3–63 × 10−6) of Ba:Ca values (Figs. 4, 6C).

Anguilla rostrata. Bivariate plots of Sr:Ca and Ba:Ca mean ± SE values, based on 3–4 element:Ca values, in the (a) elver check zone, (b) the otolith edge zone, and (c) 32–63 element:Ca values in the total transect, categorized by habitat residence group (freshwater, estuarine, migrant) for American eels (n = 21) from Muddy Hole and Castors River, western Newfoundland. SE values are not shown for panels A and B because they were essentially subsumed within the mean symbol due to small sample sizes
Effect of critical value selection on estimation of percent freshwater residence
For a given otolith Sr:Ca ratio, increasing the critical value increased the estimated %fwr and vice versa (Fig. 7A). For example, increasing the otolith Sr:Ca critical value from 3.5 to 4.0 × 10−3 increased the calculated %fwr values for all habitat groups except the freshwater group, where the %fwr was already 100% at the 3.5 critical value, and when the saline water group has no Sr:Ca values less than either critical value e.g., wholly estuarine/marine residents. The rate of change in estimated %fwr as the critical value changed was lowest at the extremes of the Sr:Ca range (freshwater and high salinity residents) and greatest for intermediate Sr:Ca values (low salinity residents and inter-habitat migrants). Thus, the single freshwater resident from Castors River not at 100%fwr went from 91.5 to 95.7 %fwr while inter-habitat migrants increased by 20.2 %fwr (range 15.4–29.1%fwr). For estuarine residents from Muddy Hole, the mean increase in %fwr with the increased critical value was 10.4 (range 2.3–18.9), to 13.1 (range 4.6–25.7) for estuarine migrants. For a critical value of 3.5 × 10−3, the non-linear fit of a polynomial of degree three (adjusted R 2 = 0.962) was little better than for a linear model (adjusted R 2 = 0.955).

Anguilla rostrata. Percent freshwater residence (%fwr) in relation to otolith Sr:Ca values at (A) a range (2.5–4.5) of freshwater to saline water critical values (CV) and (B) by site (Castors River (CR), Muddy Hole (MH)) and habitat residence group (freshwater, estuary, inter-habitat migrant) at a CV of 3.5 × 10−3, for American eels (n = 21) from western Newfoundland
The number of fish reclassified as to habitat group varied with the degree of error in the critical value used. Increasing the critical value from 3.5 to 4.0 × 10−3 produced a grand mean increase in %fwr of 8.9 (13.1 excluding freshwater residents) that was highly significant (t = 5.68, df = 20, p < 0.0001) but the increase resulted in reclassifying only one fish from estuarine resident to estuarine migrant. Decreasing the critical value to 3.0 from 3.5 decreased significantly (t = 5.46, df = 20, p < 0.0001) the grand mean %fwr by 5.5 resulting in two fish being reclassified, one each from freshwater resident to inter-habitat migrant and from inter-habitat migrant to estuarine resident. Hedges’ g effect size values were small for both the CV 3.0–3.5 (g = 0.16, 95% CI = 0.081–0.241) and CV 3.0–4.0 (g = 0.12, 95%CI = 0.060–0.181) comparisons and small-to-medium (g = 0.27, 95% CI = 0.124–0.398) for the CV 3.0–4.0 comparison. At an Sr:Ca critical value of 3.5 × 10−3, all eels classified as freshwater resident had a %fwr value >90, estuarine residents had a value <11, and inter-habitat migrants had values >20 and <85 (Fig. 7B). If a Sr:Ca critical value of 3.0 or 4.0 rather than 3.5 × 10−3 had been used in conjunction with a habitat group classification of freshwater as ≥88 %fwr, inter-habitat migrant as <88 to >12 %fwr, and estuarine as ≤12 %fwr, few (5–10%) fish would have been reclassified as to habitat residence group.
Most (90% of 21 cases) estimates of %fwr by Sr:Ca and, after the iterative process to select the best critical value, Ba:Ca, were close (<10 %fwr difference), as would be expected with a reasonable Ba:Ca critical value selection (Table 3). Acceptance of the Sr:Ca values and a critical value of 3.5 × 10−3 as the primary standard for estimating %fwr for freshwater resident eels required a Ba:Ca critical value of 1, 2, or 3 (1–3) × 10−6 for estimating %fwr at Castors River and 3, 5, or 10 × 10−6 at Muddy Hole. Inter-habitat migrants from each site required different Ba:Ca critical values, with 40 or 50 × 10−6 applicable at Castors River and 3–10 × 10−6 at Muddy Hole (Table 3).
Discussion
Habitat effects on otolith element:Ca values
The similar patterns in the Sr:Ca and Ba:Ca values in the edges of American eel otoliths as in the ambient water of the habitat of their collection site (freshwater at Castors River, estuarine at Muddy Hole) and the estimated %fwr values for each habitat residence group were consistent with the known relations between otolith element:Ca values and ambient water concentrations and salinity for Sr:Ca (higher at higher salinity) and Ba:Ca (lower at higher salinity) (Campana 1999; Bath et al. 2000; Elsdon and Gillanders 2005a, b; Walther and Thorrold 2006). Freshwater concentrations of both Sr and Ba depend largely on geomorphological conditions such as catchment geology, weathering rates, and river discharge and consequently may vary greatly among river systems while estuarine concentrations depend on river discharge, tidal stage, and the mixing pattern in the estuary (Coffey et al. 1997; Kraus and Secor 2004) and, for Ba, the pH of river discharge and extent of estuarine salt marsh (Coffey et al. 1997). Unlike Sr, Ba is adsorbed onto riverborne suspended particulate matter e.g. clays and released by ion-exchange processes in exchange for more abundant cations in estuarine and marine waters (Coffey et al. 1997) and the Ba is then removed to varying extent by biological processes (Guay and Falkner 1998) and, in some estuaries, by chemical processes (Coffey et al. 1997). Thus, the magnitude and location of peak dissolved Ba concentrations depend on the density of riverine particulate matter and hydrologic conditions.
Changes in otolith element:Ca values of sufficient magnitude may reflect a change in habitat salinity achieved either by eel movement between habitats or changes in habitat salinity about a resident eel. However, the approximately 30 days required to incorporate an Sr signal into an American eel otolith (Jessop et al. 2002), and perhaps a similar period for Ba, and the relatively large analytical “spot” size means that short-term fluctuations in ambient element:Ca values are less relevant than seasonal or annual patterns in ambient water chemistry, which are often less variable. Otolith element:Ca value changes of longer duration, say more than 2–3 values, depending on “spot” size and interval, suggest a movement between habitats because changes in habitat salinity about a single location are rarely of extended duration in open tidal estuaries such as the study sites. The issue is not whether water chemistry varies temporally or geographically but whether by doing so it creates conditions, subsequently reflected in otolith chemistry, that may make otolith element:Ca data difficult to interpret when categorizing fish habitat residence and inter-habitat movements into broad categories such as freshwater, estuarine, and marine residence and determining movements between these categories.
The significant interaction between site and otolith transect zone indicated that element:Ca values at different otolith zones varied by site, confounded by salinity, as would be expected for the use of element:Ca values to be used as geographic tags. The absence of any significant difference between sites in the Sr:Ca and Ba:Ca values in the otolith core of American eels was expected given the general uniformity of oceanic element:Ca values, relative to estuarine or freshwater values (Campana 1999), over their oceanic migration as larvae and arrival in continental waters. Marine concentrations of Sr are relatively constant at about 8 mg·L−1 (Angino et al. 1966) while Ba ranges between 8 and 13 μg·L−1 in the Atlantic Ocean and increases in concentration with depth (Neff 2002). High Sr:Ca ratios in the otolith core area are an ontogenetic effect typical of anguillid eels due to biological processes during the oceanic leptocephalus stage, with values decreasing after metamorphosis to the glass eel stage (Otake et al. 1994; Tzeng and Tsai 1994; Arai et al. 1997).
Ontogenetic effects on otolith Ba:Ca values
Whether Ba:Ca values in the otolith core zone are influenced by ontogeny in a manner similar to that for Sr:Ca is unknown. However, the significant decline in otolith Ba:Ca between the core and edge for eels from Muddy Hole is contrary to the expectation of an increase as salinity decreases from marine to estuarine values. Otolith element:Ca ratios are most influenced by the molar ratios of the ambient elements not their concentration ratios (Campana 1999). Consequently, the occurrence of a molar element:Ca ratio in the core zone that is 1.9 times the ambient oceanic molar ratio for Ba and only 0.6 times for Ca suggests that Ba is also influenced by ontogeny during the larval phase, as is Sr (Otake et al. 1994). Unlike this study, Arai and Hirata (2006) found higher Ba:Ca values before the elver check (corresponding to the oceanic life stage) for eels characterized as freshwater residents than for seawater residents, leading them to hypothesize that such different elemental signatures might originate from different parental groups and that populations that spawn at different times exist for Japanese eels. Otolith Sr:Ca values are not influenced by ontogeny in the juvenile stage (Otake et al. 1994) nor, apparently, are Ba:Ca values.
Inter-habitat migration effects on otolith element:Ca transect patterns
Mean element:Ca values over an otolith transect from elver check to edge may obscure the highly variable patterns that can occur over segments of the transect, particularly for inter-habitat migrants that transition from one habitat to another, for example, early or later in life. Inter-habitat movements are common by a portion of the anguillid eel population in coastal zones (reviewed by Daverat et al. 2006; Jessop et al. 2008) and the three temperate-zone species are regarded as facultatively catadromous. A high otolith mean Sr:Ca and low Ba:Ca corresponded to estuarine residence while low Sr:Ca and high Ba:Ca corresponded to freshwater residence, consistent with patterns found for other species and may be considered a habitat effect (Arai and Hirata 2006; Crook et al. 2006; 2008; Tabouret et al. 2010). The variable relations between Ba:Ca and Sr:Ca for inter-habitat migrants moving from fresh to estuarine (intermediate Sr:Ca, high Ba:Ca) and estuarine to fresh waters (intermediate Sr:Ca, intermediate Ba:Ca) demonstrate that Ba:Ca values may be an inconsistent indicator of estuarine residence. Consequently, the migratory history of anguillid eels should be considered when evaluating otolith element:Ca values with respect to historical habitat residence.
Habitat reconstructions from otolith Sr:Ca and Ba:Ca patterns may differ
The occurrence of high Ba:Ca values in conjunction with high Sr:Ca values for portions of the life history of some inter-habitat migrant American eels requires caution in concluding that Ba:Ca values alone, without reference to Sr:Ca values, can be generally used to determine residence in saline or fresh waters, as was done, for example, for black bream (Acanthopagrus butcheri) by Elsdon and Gillanders (2005b) based on the occurrence of ambient Ba:Ca ratios that were higher in fresh water than in salt water. Element incorporation into otoliths is species specific (Zimmerman 2005; Hamer and Jenkins 2007), illustrated by the difference in Ba:Ca critical values for Japanese eels (≥6 × 10−6) (Arai and Hirata 2006), black bream (Elsdon and Gillanders 2005b), and American eels. The variability in ambient Ba:Ca concentrations between river systems due to differences in local geology, river discharge and level of suspended particulate matter, estuarine salinity gradients, and temperature (Coffey et al. 1997; Elsdon and Gillanders 2005b; Dorval et al. 2007) may not permit development of a general Ba:Ca critical value for the freshwater to estuarine transition.
The occurrence of high Ba:Ca values in the estuarine-resident portions of inter-habitat migrants from both Muddy Hole and Castors River suggests that these eels resided close to the river mouth in the estuarine mixing zone. Barium is released from river-borne suspended particulate matter during estuarine mixing (Coffey et al. 1997). The desorption of Ba is low at salinities <1 but is strong and rapid at salinities of 1–2. The location of peak desorption is controlled by hydrodynamic conditions, occurring at higher salinities where river flows are low (Coffey et al. 1997). Thus, the greatest variation in ambient Ba:Ca appears to occur at low salinities (0–10%) (Coffey et al. 1997; Tabouret et al. 2010), as does the variation in otolith Ba:Ca (Elsdon and Gillanders 2005b). Elsdon and Gillanders (2004) caution that ambient Ba level may have a greater influence on otolith concentration than does salinity. Otolith Sr:Ca evidently has a more consistent relation with ambient salinity than does Ba:Ca, particularly at estuarine-level salinities.
Residence in and movement between fresh and marine waters may be determined (Elsdon and Gillanders 2005b; Macdonald and Crook 2010) if only because the ambient level and variability of Ba:Ca in marine waters is much less than in estuarine and fresh waters. Otolith Ba:Ca values <3.2 × 10−6 at salinities >20 were considered to represent salt water residence for European eels (Tabouret et al. 2010) while Arai and Hirata (2006) considered sea eels to have a mean ± SD of less than about 3.0 × 10−6 and a similar value might be accepted as an estuarine-marine transition critical value.
The presence of downstream migrant eels with otolith edge Sr:Ca values indicative of estuarine habitat as opposed to the freshwater of their capture has been noted in several studies but no clear explanation is available (Jessop et al. 2008). Explanations include edge artefact and a brief, unregistered in the otolith, return to freshwater before migrating downstream. The occurrence in Castors River of eels with low and high mean lifetime Ba:Ca values in freshwater suggests that they may have come from regional habitats within the river system that differ in levels of suspended particulate matter or geological content of Ba (Coffey et al. 1997; Crook et al. 2008).
Effect of Sr:Ca critical value selection
Appropriate selection of the critical value separating fresh from saline water residence for element:Ca data is important because it may significantly affect the estimation of %fwr and result in incorrect categorizations of habitat residence/migratory group. A higher than justified critical value may push estuarine residents into the low %fwr inter-habitat migrant group but does not greatly affect those categorized as fully freshwater while a lower than justified critical value little affects fish already categorized as estuarine residents and pushes freshwater residents towards the high %fwr inter-habitat migrant group, high %fwr inter-habitat migrants towards low %fwr, and low %fwr inter-habitat migrants to the fully estuarine resident group. The broad categorization of habitat residence group may be fairly robust to some error in the selection of Sr:Ca critical value. The biological importance is uncertain of mean differences of about 10% in %fwr in response to a change in critical value of about 0.5×10−3 and resulting in a small effect size. Medium to large effect sizes have been classified as biologically important (Arnqvist and Wooster 1995) and effect size may be positively associated with biological importance but the biological importance of an effect size may ultimately be determined by the nature of the study and the researcher’s judgement (Cohen 1988). The %fwr boundaries for each primary habitat residence group considered here (freshwater, estuarine) are based on observed patterns, with the objective that a residence group, say freshwater, should maximize the freshwater characteristic and minimize the estuarine characteristic so commonly observed for eels resident in near-coastal fresh waters (Jessop et al. 2008).
Optimal management requires a better understanding of the proportions of an eel stock occupying a given habitat type and of the frequency and duration of inter-habitat movements. Such estimates depend upon the correct classification of individual eels into habitat residence groups, usually via the inspection of otolith element:Ca transect patterns and specification of a critical value for otolith element:Ca values signifying the boundary between habitat residence groups. The use of Ba can assist the use of Sr in the evaluation of the environmental history of habitat residence and inter-habitat movement of anguillid eels via temporal patterns in otoliths but the complexity of eel life history and the wide range of habitats occupied can present difficulties of interpretation for the sole use of Ba, particularly for inter-habitat migrants.
References
Angino EE, Billings GK, Andersen N (1966) Observed variations in the strontium concentration of sea water. Chem Geol 1:145–153
Arai T, Hirata T (2006) Differences in the trace element deposition in otoliths between marine- and freshwater-resident Japanese eels, Anguilla japonica, as determined by laser ablation ICPMS. Env Biol Fish 75:173–182
Arai T, Otake T, Tsukamoto K (1997) Drastic changes in otolith microstructure and microchemistry accompanying the onset of metamorphosis in the Japanese eel Anguilla japonica. Mar Ecol Prog Ser 161:17–22
Arnqvist G, Wooster D (1995) Meta-analysis: synthesizing research findings in ecology and evolution. Tree 10:236–240
Bath GE, Thorrold SR, Jones CM, Campana SE, McLaren JW, Lam JWH (2000) Strontium and barium uptake in aragonitic otoliths of marine fish. Geochim Cosmochim Acta 64(10):1705–1714
Bradbury IR, Campana SE, Bentzen P (2008) Otolith elemental composition and adult tagging reveal spawning site fidelity and estuarine dependency in rainbow smelt. Mar Ecol Prog Ser 368:255–268
Bulger AJ, Haden BP, Monaco ME, Nelson DM, McCormick-Day MG (1993) Biologically-based estuarine salinity zones derived from a multivariate analysis. Estuaries 16(2):311–322
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297
Coffey M, Dehairs F, Collette O, Luther G, Church T, Jickells T (1997) The behaviour of dissolved barium in estuaries. Estuar Coast Shelf Sci 45:113–121
Cohen J (1988) Statistical power analysis for the behavioral sciences, 2nd edn. Erlbaum, Hillsdale
COSEWIC (Committee on Status of Endangered Wildlife in Canada) (2006) Status Report – American Eel. COSEWIC. Available: www.sararegistry.gc.ca/species/speciesDetails_e.cfm?sid=891. (June 2008)
Crook DA, Macdonald JI, O’Connor JP, Barry B (2006) Use of otolith chemistry to examine patterns of diadromy in the threatened Australian grayling Prototroctes maraena. J Fish Biol 69:1330–1344
Crook DA, Macdonald JI, Raadik TA (2008) Evidence of diadromous movements in a coastal population of southern smelts (Retropinninae: Retropina) from Victoria, Australia. Mar Freshw Res 59:638–646
Daverat F, Tomás J (2006) Tactics and demographic attributes in the European eel Anguilla anguilla in the Gironde watershed, SW France. Mar Ecol Prog Ser 307:247–257
Daverat F, Tomas J, Lahaye M, Palmer M, Elie P (2005) Tracking continental habitat shifts of eels using otolith Sr/Ca ratios: validation and application to the coastal, estuarine and riverine eels of the Gironde-Garonne-Dordogne watershed. Mar Freshw Res 56:619–627
Daverat F, Limburg KE, Thibault I, Shiao J-C, Dodson JJ, Caron F, Tzeng W-N, Iizuka Y, Wickström H (2006) Phenotypic plasticity of habitat use by three temperate eel species, Anguilla anguilla, A. japonica and A. rostrata. Mar Ecol Prog Ser 308:231–241
Dorval E, Jones CM, Hannigan R, van Montfrans J (2007) Relating otolith chemistry to surface water chemistry in a coastal plain estuary. Can J Fish Aquat Sci 64:411–424
Elsdon TS, Gillanders BM (2002) Interactive effects of temperature and salinity on otolith chemistry: challenges for determining environmental histories of fish. Can J Fish Aquat Sci 59:1796–1808
Elsdon TS, Gillanders BM (2004) Fish otolith chemistry influenced by exposure to multiple environmental variables. J Exp Mar Biol Ecol 313:269–284
Elsdon TS, Gillanders BM (2005a) Consistency of patterns between laboratory experiments and field collected fish in otolith chemistry: an example and applications for salinity reconstructions. Mar Freshw Res 56:609–617
Elsdon TS, Gillanders BM (2005b) Alternative life-history patterns of estuarine fish: barium in otoliths elucidates freshwater residency. Can J Fish Aquat Sci 62:1143–1152
Elsdon TS, Wells BK, Campana SE, Gillanders BM, Jones CM, Limburg KE, Secor DH, Thorrold SR, Walther BD (2008) Otolith chemistry to describe movements and life-history parameters of fishes: hypotheses, assumptions, limitations and inferences. Ocean Mar Biol: An Ann Rev 46:297–330
Guay CK, Falkner KK (1998) Barium as a tracer of Arctic halocline and river waters. Deep-sea Res II 44(8):1543–1569
Haro A, Richkus W, Whalen K, Hoar A, Busch W-D, Lary S, Brush T, Dixon D (2000) Population decline of the American eel. Fisheries 25(9):7–16
Hamer PA, Jenkins GP (2007) Comparison of spatial variation in otolith chemistry of two fish species and relationships with water chemistry and otolith growth. J Fish Biol 71:1035–1055
Jessop BM (1997) An overview of European and American eel stocks, fisheries, and management issues. In Peterson, R. H. (ed.) The American eel in Eastern Canada: stock status and management strategies. Proceedings of Eel Management Workshop, January 13–14, 1997, Quebec City, Q.C. Can Tech Rep Fish Aquat Sci No 1296:6–20
Jessop BM, Shiao JC, Iizuka Y, Tzeng WN (2002) Migratory behaviour and habitat use by American eels Anguilla rostrata as revealed by otolith microchemistry. Mar Ecol Prog Ser 233:217–229
Jessop BM, Shiao JC, Iizuka Y, Tzeng WN (2007) Effects of inter-habitat migration on the evaluation of growth rate and habitat residence of American eels Anguilla rostrata. Mar Ecol Prog Ser 342:255–263
Jessop BM, Cairns DK, Thibault I, Tzeng WN (2008) Life history of American eel Anguilla rostrata: new insights from otolith microchemistry. Aquat Biol 1:205–216
Jessop BM, Shiao JC, Iizuka Y (2009) Life history of American eels from western Newfoundland. Trans Am Fish Soc 138:861–871
Kalish JM (1989) Otolith microchemistry: Validation of the effects of physiology, age, and environment on otolith composition. J Exp Mar Biol Ecol 132:151–178
Kawakami Y, Mochioka N, Morishita K, Tajima T, Nakagawa H, Toh H, Nakazono A (1998) Factors influencing otolith strontium/calcium ratios in Anguilla japonica elvers. Envir Biol Fish 52:299–303
Kotake A, Okamura A, Yamada Y, Utoh T, Arai T, Miller MJ, Oka HP, Tsukamoto K (2005) Seasonal variation in the migratory history of the Japanese eel Anguilla japonica in Mikawa Bay, Japan. Mar Ecol Prog Ser 293:213–221
Kraus RT, Secor DH (2004) Incorporation of strontium into otoliths of an estuarine fish. J Exp Mar Biol Ecol 302:85–106
Macdonald JI, Crook DA (2010) Variability in Sr:Ca and Ba:Ca ratios in water and fish otoliths across an estuarine salinity gradient. Mar Ecol Prog Ser 413:147–161
Martin GB, Wuenschel MJ (2006) Effect of temperature and salinity on otolith element incorporation in juvenile gray snapper Lutjanus griseus. Mar Ecol Prog Ser 324:229–239
Miller JA (2009) The effects of temperature and water concentration on the otolith incorporation of barium and manganese in black rockfish Sebastes melanops. J Fish Biol 75:39–60
Miller JA, Gray A, Merz J (2010) Quantifying the contribution of juvenile migratory phenotypes in a population of Chinook salmon Oncorhynchus tshawytscha. Mar Ecol Prog Ser 408:227–240
Nakagawa S, Cuthill IC (2007) Effect size, confidence interval and statistical significance: a practical guide for biologists. Biol Rev 82:591–605
Neff JM (2002) Bioaccumulation in marine organisms: effects of contaminants from oil well produced water. Elsevier, Oxford
Olkin I, Pratt JW (1958) Unbiased estimation of certain correlation coefficients. Ann Math Stat 29:201–211
Otake T, Ishii T, Nakahara M, Nakamura R (1994) Drastic changes in otolith strontium/calcium ratios in leptocephali and glass eels of Japanese eel Anguilla japonica. Mar Ecol Prog Ser 112:189–193
Porter TR, Riche LG,Traverse GR (1974) Catalogue of rivers in insular Newfoundland. Resource Development Branch, Newfoundland Region. Data Rep Ser No. New/D-74-9 vol. C
R Development Core Team (2010) R: A language and environment for statistical computing, reference index version 2.12.1. R Foundation for statistical computing, Vienna, Austria. URL http://www.R-project.org
Secor DH, Rooker JR (2000) Is otolith strontium a useful scalar for life-cycles in estuarine fishes? Fish Res 46:359–371
Shiao JC, Ložys L, Iizuka Y, Tzeng WN (2006) Migratory patterns and contribution of stocking to the population of European eels in Lithuanian waters as indicated by otolith Sr:Ca ratios. J Fish Biol 69:749–769
Tabouret H, Bareille G, Claverie F, Pécheyran C, Prouzet P, Donard OFX (2010) Simultaneous use of strontium:calcium and barium:calcium ratios in otoliths as markers of habitat: Application to the European eel (Anguilla anguilla) in the Adour basin, South West France. Mar Envir Res 70:35–45
Thibault I, Dodson JJ, Caron F, Tzeng W-N, Iizuka Y, Shiao J-C (2007) Facultative catadromy in American eels: testing the conditional strategy hypothesis Mar Ecol Prog Ser 344:219–229
Tsukamoto K, Arai T (2001) Facultative catadromy of the eel Anguilla japonica between fresh water and seawater habitats. Mar Ecol Prog Ser 220:265–276
Tzeng W-N (1996) Effects of salinity and ontogenetic movements on strontium:calcium ratios in the otoliths of the Japanese eel, Anguilla japonica Temminck and Schlegel. J Exp Mar Biol Ecol 199:111–122
Tzeng WN, Tsai YC (1994) Changes in otolith microchemistry of the Japanese eel, Anguilla japonica, during its migration from the ocean to the rivers of Taiwan. J Fish Biol 45:671–683
Tzeng WN, Wu HF, Wickström H (1994) Scanning electron microscope analysis of annulus microstructure in otolith of European eel, Anguilla anguilla. J Fish Biol 45:479–492
Tzeng W-N, Severin KP, Wickström H (1997) Use of otolith microchemistry to investigate the environmental history of European eel Anguilla anguilla. Mar Ecol Prog Ser 149:73–81
Tzeng WN, Shiao JC, Iizuka Y (2002) Use of otolith Sr:Ca ratios to study the riverine migratory behaviors of the Japanese eel Anguilla japonica. Mar Ecol Prog Ser 245:213–221
USFWS (United States Fish & Wildlife Service) (2005) Progress Report: The American Eel Status Review. USFWS. Available: www.fws.gov/northeast/ameel/statusreprog.html. (June 2008)
USFWS (United States Fish & Wildlife Service) (2007) Endangered and threatened wildlife and plants; 12-month finding on a petition to list the American eel as threatened or endangered. Federal Register 72:22(2 February 2007):4967–4997
Walther BD, Thorrold SR (2006) Water, not food, contributes the majority of strontium and barium deposited in the otoliths of a marine fish. Mar Ecol Prog Ser 311:125–130
Zar JH (1984) Biostatistical analysis, 2nd edn. Prentice-Hall, Englewood Cliffs
Zimmerman CE (2005) Relationship of otolith strontium-to-calcium ratios and salinity: experimental validation for juvenile salmonids. Can J Fish Aquat Sci 62:88–97
Acknowledgements
We thank M. Feigenbaum and M. Campbell for providing the eel samples and for use of the sample processing facilities at South Shore Trading Company Ltd, G. Sneddon and S. Stewart for assistance with mixed effects models, the National Science Council of the Republic of China for funding the otolith microchemistry analysis, and members of the Earth Dynamic System Research Center, National Cheng Kung University for assisting the ICP-MS analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jessop, B.M., Wang, CH., Tzeng, WN. et al. Otolith Sr:Ca and Ba:Ca may give inconsistent indications of estuarine habitat use for American eels (Anguilla rostrata). Environ Biol Fish 93, 193–207 (2012). https://doi.org/10.1007/s10641-011-9905-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-011-9905-0