
Abstract
The numbers of spawning sites for Chinese sturgeon have been drastically reduced since the construction of the Gezhouba Dam across the Yangtze River. This dam has blocked migration of Chinese sturgeon to their historic spawning ground causing a significant decline of the Chinese sturgeon population. We conducted a VORTEX population viability analysis to estimate the sustainability of the population and to quantify the efficiency of current and alternative conservation procedures. The model predicted the observed decline of Chinese sturgeon, resulting from the effect of the Gezhouba Dam. These simulations demonstrated the potential interest of two conservation measures: increasing spawning area and reducing predation on sturgeon eggs. The simulations also demonstrated that the actual restocking program is not sufficient to sustain sturgeon population as the artificial reproduction program induce the loss of more wild mature adults that the recruitment expected by the artificial reproduction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Dams are known as one of the most negative human impacts on river ecosystems, as they modify physical environment and regulate flow (Allan 1996). This has significant consequences for fragmentation of habitats, blocking migration routes, and destroying spawning grounds, causing loss of biodiversity (Baxter 1977; Dudgeon 2000), as experienced on a large variety of reservoirs all around the world (World Commission on Dams 2000). Despite the negative impact of these constructions, the amount of damming projects is still increasing and dams are bigger and bigger, like the Three–Gorges Dam (TGD). This dam, is the largest dam ever built, it is 193 m high and the resulting reservoir floods over 600 km of the Yangtze course. It deeply modify the Yangtze ecosystem and will probably strongly affect both animal and vegetal communities (Wu et al. 2003). Indeed, Park et al. (2003) predicted a biodiversity decline of endemic fishes of the upper Yangtze River caused by the TGD. In addition, Xie (2003) pointed out the potential negative impact of TGD, combined with the Gezhouba Dam (located 38 km downstream from TGD) on the ancient endemic fish, such as Chinese sturgeon (Acipencer sinensis), Yangtze sturgeon (A. dabryanus), and Chinese paddlefish (Psephurus gladius).
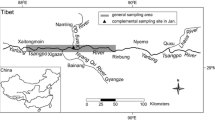
Biological information on Chinese paddlefish and Yangtze sturgeon are too meager to forecast the sustainability of these species. On the contrary, Chinese sturgeon population biology was intensively studied since the 1970s (Anonymous 1988). This species, due to its large size (more than 4 meters long and 450 kg weight, Chang and Cao 1999) but also to its ancient history (paintings representing Chinese sturgeons made more than 2000 years ago have been found in China) is considered as one of the three most emblematic fish species in the Yangtze River together with the Chinese paddlefish and the Yangtze sturgeon. This is also a vulnerable species, like most of the sturgeons, due to its late maturity (Waldman and Wirgin 1997; Ludwig 2008). Prior to construction of the Gezhouba Dam, the original spawning area was located in the upstream part of the Yangtze River, The 16 known natural spawning grounds were dispersed in a 800 km length reach, from upstream of Xinshi to downstream of Fuling (Fig. 1) (Anonymous 1988; Wei et al. 1997). The construction of the Gezhouba Dam, in 1981, blocked the migration route and all the natural spawning grounds became unavailable. Only one new spawning ground which has been recorded since 1981 is located just downstream the Gezhouba Dam and represented by a 3 km long area (Fig. 1). The area of this alternative spawning site represents less than 1% of the ancient spawning area (around 800 km), as well as the carrying capacity of this alternative spawning site is about 10% of the ancient one, leading to a decline of the population (Wei et al. 1997; Chang 1999; Yi et al. 1999). Aiming to conserve this emblematic species, the Chinese government established a series of protection measures, such as prohibiting commercial fishing since 1983, setting up an artificial propagation program (leading to release larvae and juveniles since 1984), protecting actual spawning ground, and initiating studies on the biology and the life history of Chinese sturgeon (Fu et al. 1985; Chang and Cao 1999). However the sustainability of the last population of Chinese sturgeon is still unknown. Our aim was therefore to estimate the status of the species using a population viability analysis (PVA).

Map of China showing the location of historical and actual Chinese sturgeon (A. sinensis) spawning area. The two dams, Three-Gorges Dam (TGD) and Gezhouba Dam are indicated on the map
PVA methods are mathematical simulations of the dynamics of a population. These models integrate the biological characteristics of the considered population and permit us to estimate the impact of environmental or human disturbances on the sustainability of the population, expressed for example as a population growth rate. PVA methods are now extensively used to estimate extinction risk of endangered species and provide effective guidelines for establishing conservation measures (Lindenmayer and Possingham 1995; Miller and Lacy 2005). However, the precision of extinction risk prediction has been the subject of some debate (Ludwig 1999; Fieberg and Ellner 2000; Coulson et al. 2001; Brook et al. 2002; Ellner et al. 2002; Reed et al. 2002; McCarthy et al. 2003). Despite uncertainties on the accuracy of the predictions, due to incomplete biological information, stochastic environmental events and the complexity of feedback mechanisms, PVA are recognized as reliable methods to estimate species sustainability (Brook et al. 2000) and useful tools to manage threatened species (Mathews and Macdonald 2001; Lunney et al. 2002).
Based on previous studies on the population biology of the Chinese sturgeon, our aim was to build a population viability analysis (PVA) model to estimate the sustainability of this species in the Yangtze River. Then, we used this model to quantify the efficiency of actual and alternative management procedures, leading to discuss their interest for Chinese sturgeon conservation.
Material and methods
Chinese sturgeon
Chinese sturgeon is an anadromous species, which historically reproduced in the upstream parts of two large rivers: Yangtze and Pearl rivers (Fig. 1). However, due to human impact, the Chinese sturgeon is now rarely found in the Pearl River, and the Yangtze River is probably its last refuge (Wei et al. 1997). Spawning stocks were estimated as well as biological characteristics of the species were recorded since 1981 by the Hydrobiology Institute of the Chinese Academy of Sciences in Wuhan, the Chinese Sturgeon Institute in Yichang and Yangtze River Fisheries Research Institute in Shashi. Since 1981, sampling has been done each year during the reproduction period. Fish spawning stock were estimated using hydroacoustic methods and biological characteristics (see Table 1) were measured by fish captures. Indeed, each year, mature adults were captured for artificial reproduction purposes. For more detail on the biological characteristics and population structure of Chinese sturgeon, see Deng et al. (1985), Chang (1999) and Wei (2003).
PVA model and scenarios
Population sustainability of Chinese sturgeon was estimated using the VORTEX (Lacy 1993) PVA simulation method. The choice of VORTEX was guided by the ability of this model to incorporate available details on sturgeon life history. Moreover, this modeling procedure is commonly used to forecast sustainability of endangered species, and it has been recognized as an efficient and flexible method, providing similar predictions to the other sustainability analysis programs (Brook et al. 2000). The VORTEX model is a Monte Carlo simulation of the effects of deterministic forces as well as demographic, environmental and genetic stochasticity on the dynamics of a population (Lacy 2000; Miller and Lacy 2005). This program models population processes as discrete sequential events with probabilistic outcomes. It is an individual-based system that keeps track of each member of a population. Life history processes are therefore simulated by generating random numbers to determine whether each individual live or dies and to determine the reproductive output of each female at each reproduction. Full details of VORTEX functioning are given by Lacy (1993).
The VORTEX model of Chinese sturgeon population dynamics was built using actual knowledge on the species and these parameters are given in Table 1. Although the VORTEX model can be used on any kind of animals, it can not deal with total initial population sizes larger than 30 000 (Miller and Lacy 2005). This is easily exceeded for high fecundity species (such as fish); in that case, Lacy (1993) proposed to restrict the population considered in the model to sub-adult and adult individuals to avoid biases in the simulations. The size and age structure of Chinese sturgeon population has been roughly estimated to c.a. 90 000 fish older than one year (Gao 2007). The estimation was based on 1981 biological survey of the species conducted by the Chinese Academy of Sciences and all available information on the species. As this fish number exceeded the VORTEX program capabilities, we followed the recommendation of Miller and Lacy (2005) to restrict the population size to adults and sub-adults. We hence restricted our analysis to the fish older than 6 years, and the initial population size was then set to 26 930 individuals (i.e. total estimated number of Chinese sturgeon older than 6 years old; Table 1). This age also correspond to the beginning of sub-adult fish migration from the sea to the Yangtze River. It was then possible to record these fish in the Yangtze River. Indeed, it is almost impossible to estimate sturgeon survival during the first years, as the individuals can not be counted in a large river such as the Yangtze. This procedure proposed by Miller and Lacy (2005) to deal with large population sizes, was efficiently employed for population viability analysis of trout using VORTEX (Brook et al. 2000; Rieman and Allendorf 2001) (See Appendix). Due to the large population size, we assumed there was no inbreeding depression, as demonstrated by Zhang et al. (2003). After construction of the Gezhouba Dam, the new spawning ground below the dam was not sufficient to permit all the mature fish to spawn (Qiao et al. 2006). Therefore, we considered that the dam induced a density-dependent reproduction due to the loss of natural spawning habitats (Table 1). We expressed the percentage (P F ) of females breeding density-dependently as
where S was the carrying capacity of the spawning ground, namely the number of spawners (Chang 1999). A (time) was the number of spawning activity each year (Wei 2003). F was the number of adult females considered in the VORTEX model and P i was the percentage of adult females breeding (Gao 2007).
Male sex ratio at birth was estimated as the proportion of 6 years old males to 6 years old individuals. According to the previous studies on sturgeon biology and the calculated population structure, the average fecundity was estimated to 10.5 six-year old offspring per female (Gao 2007). Males in breeding pool were considered as the proportion of mature males to males older than 13 years. Natural mortality rate was estimated as described by Chen and Watanabe (1989). Since fishing would be involved as an input parameter, the fishing mortality was neglected here to avoid “double dipping” (Brook 2000). The actual carrying capacity is unknown, but the population size was stable before damming due to a constant recruitment (Chang 1999). We considered that the initial population size approached carrying capacity which was assumed equal to 30 000, according to the VORTEX model prescriptions (Miller and Lacy 2005). One “catastrophe” was considered, it accounted for 238 individuals killed by ships propellers from 1984 to 1998 (Chang 1999). This occurs each year, so the “catastrophe” frequency was 100% and its severity in reproduction and survival rate was 0.999 and 0.987, respectively. Since 1984 spawners were captured to perform artificial reproduction and restocking (i.e. supplementation). In the model, supplementation and harvest were considered as artificial impacts on the population.
The VORTEX parameters were calculated using species-specific and population–specific data obtained from field surveys (Table 1). The population was modeled over 100 years with each simulation repeated for 1000 iterations. The model output provided population growth rate, extinction probability and population size dynamics. Differences between models simulations were tested using Student t-test. We first predicted population dynamics using the parameters of sturgeon biology given in Table 1. This accounted for the actual situation, including impact of the dam and conservation measures set since the Gezhouba Dam construction (no commercial fishing, restocking…). Restocking is the current conservation strategy. This program has been set since 1984 by the Institute of Chinese Sturgeon. From 1984 to 1999, up to 100 mature fishes were permitted to be captured each year. Mature fish were captured and used to produce juveniles by artificial reproduction. Since 1999, the catches were reduced to less than 50 fish per year (Chang and Cao 1999). Each year since 1984, juveniles were produced by artificial reproduction and released to the Yangtze. However, the number of juveniles released did not exceed 20 000 fish per year. In 1998, about 20 000 juveniles were released to the Yangtze (Xiao et al. 1999). Gao (2007) estimated that this represents about 4% of the juvenile population in the estuary based on the mark-recapture and genetic data (Chang 1999; Zhu et al. 2002). Morover, the long term biological survey of Chinese sturgeon population dynamics in place since 1984, allowed us to estimate among the 20 000 released juveniles only 34 will survive up to 7 years of age (Gao 2007). To provide a detailed understanding of the drawbacks of restocking we used our simulations to test the negative effect of adult capture for artificial reproduction purposes and the positive effect of restocking (only 20 000 juveniles per year was tested as this corresponds to the actual restocking level). However, due to both individual variability and uncertainties in the biology of sturgeon, we provided simulations ranging from best to worst situation (Table 1). In that way, the outcome of the sturgeon population is comprised between these two extreme values. Then, based on parameters estimated by the average value of the biological data, we constructed a basic simulation of sturgeon population size dynamics. This basic simulation comprised no restocking and no harvest, to provide a situation without human impact except the impact of the dam. The basic simulation was then used to run a series of simulations to simulate the effect of different human actions in terms of conservation or disturbances for the species. These scenarios were modeled to help identify the impact of some human activities as well as the interest of some conservation measures to the sustainability of Chinese sturgeon population.
The 6 scenarios tested were the following:
-
Scenario 1: Impact of the dams
-
In this scenario we considered that the removal of the dams (TGD and Gezhouba) would allow recovery of the original spawning grounds surface and therefore improve spawning. In the basic simulations, the impact of the dams was considered as density-dependent reproduction due to the small size of the spawning ground. The assumption without density-dependent reproduction, which represented the status without impact of the dam, was simulated and compared with the basic assumption.
-
Scenario 2: Utility of supplementation
-
The supplementation comes from restocking permitted by artificial reproduction. The supplementation range tested, equivalent of 20, 30 and 40 seven year old individuals, was consistent with actual restocking. This allowed us to test if the artificial supplementation constitutes a useful conservation strategy.
-
Scenario 3: Impact of fish harvest
-
Commercial fishing is forbidden since 1983, but adults are still captured for scientific purposes and artificial reproduction. We therefore tested the impact of the adults harvest on population sustainability. Three harvest levels were tested: 20, 50 and 100 individuals per year (50% male and 50% female).
-
Scenario 4: Modifying spawning ground size
-
The remaining spawning site is less than 1% of the historic spawning area. In this scenario we tested the effect of increasing spawning ground size, for example by building artificial spawning grounds. Therefore, we simulated increases of 50%, 100% and 200% in spawning ground size. We also simulated a decrease of 50% of the carrying capacity of the spawning ground. This simulates the effect of the TGD, as a 41.6 % reduction of the flow downstream Gerzhouba dam is expected due to TGD functioning.
-
Scenario 5: Increasing fecundity
-
A large proportion of Chinese sturgeon eggs are eaten by benthic fish (mainly largemouth bronze gudgeon (Coreius guichenoti), bronze gudgeon (C. heterodon) and dark barbel (Pelteobagrus vachelli)). This predation pressure drastically reduces recruitment and affects population dynamics (Hu et al. 1992; Chang 1999; Yu et al. 2002). In this scenario, we simulated a reduction of benthic fish predation pressure on sturgeon eggs, leading to increased sturgeon fecundity. The impact of a set of fecundity improvements were modeled as 50%, 100% and 200% increase of offspring numbers.
-
Scenario 6: combination of spawning ground and fecundity
-
In this scenario, the effects of simultaneous implementation of two conservation measures suggested above were tested. These combinations of simultaneous increases of 50%, 100%, and 200% in both carrying capacity of spawning ground and offspring numbers were simulated.
Results
The best and worst cases for Chinese sturgeon predicted a decline of the sturgeon (Fig. 2), with a growth rate of between 1.1% and −17.2% per year and a population size comprised between 24 598 (±3 343) and 0 individuals after 100 years simulation. The best status predicted a sustainable population whereas the worst status predicted extinction in 35.1 years (Table 2).

Population size evolution on a 100 years time scale simulated using actual characteristics of the Chinese sturgeon population. The number of individuals is given as a range corresponding to minimal and maximal values of life histories traits introduced in the model (i.e. best and worst status)
Considering the 6 sets of scenarios used to simulate the effects of human management or disturbances on Chinese sturgeon sustainability, population size significantly differed (P < 0.001) between the basic assumption and all the scenarios, showing that the human activities influence population size. However, this result is balanced by the analysis of population growth rate, indicating that increasing supplementation (scenario 2) did not affected significantly (P < 0.05) population growth rate (Table 3). In that way, for this scenario, the population showed a similar decreasing dynamic as the simulation of the basic scenario (Fig. 3d and e).

Population size evolution on a 100 years time scale simulated according to 5 conservation scenarios simulating the effect of different anthropic actions in terms of conservation or disturbances for the species. (a) Scenario 1- impact of the dams; (b) Scenario 2- increasing supplementation; (c) Scenario 3- increasing adult harvest; (d) Scenario 4- modifying spawning ground size; (e) Scenario 5- increasing fecundity; (f) Scenario 6- combination of increasing spawning ground and fecundity
The four scenarios, namely: impact of the dams (scenario 1), increasing spawning ground size (scenario 4), increasing fecundity by protecting eggs against predation (scenario 5) and the impact of increasing fecundity and spawning ground (scenario 6) had a significant influence on both population size and growth rate (Table 3). Removal of the dams was the only scenario which predicted a stabilization of the population size (Fig. 3a) and a positive population growth rate (Table 3). In scenario 3, the increase of adults harvest could have a dramatic effect on the sustainability of the species. A fishing strength of 100 adults per year could induce a steep decline of the population. This simulation was the only case where the extinction probability (PE) was not zero (PE = 1) and the mean extinction time (Mean TE) predicted was 65.5 years (Fig. 3c and Table 3), showing that the population can decline rapidly in the case of intensive harvests. Even if decreasing the fishing pressure by 50%, there was still a significant impact resulting from harvest of adults (Fig. 3c and Table 3). In the same way, the increase in fecundity and spawning ground could provide efficient means of conservation, as population survivals were clearly higher than for the basic assumption (Fig. 3d and e). Comparing with the basic scenario, even a small (e.g. 50%) increase in fecundity and spawning ground induced a significant increase of both population size (P < 0.001) and growth rate (P < 0.05). In Scenario 4, reducing the size of spawning ground to 50% of its actual surface caused a significant decrease in both population growth rate and population size (P < 0.001) (Fig. 3d and Table 3). In scenario 6, combination of two conservation strategies described above had a significant positive effect on both population size (P < 0.001) and growth rate (P < 0.001). Particularly, an increase of both spawning ground surface and early survival rate would permit stop the decline of the species and acquire a population growth rate up to 5% (Fig. 3f; Table 3). Moreover, the effect of simultaneous improvements in carrying capacity of spawning grounds and early survival rate significantly (P < 0.001) exceeded that of a single conservation measure.
Discussion
The long lifespan and the late maturity of the Chinese sturgeon makes field experimentations almost impossible and experimental and hazardous management strategies could endanger the entire species. In that context, the simulation of “virtual populations” offered the only feasible approach to estimate the impact of different conservation or management strategies. However, the results of PVA models should be considered with caution, as according to Taylor (1995) and McCarthy et al. (2003), simulations can only provide trends on the likely responses of a population to changes in its environment, but not exact values on the future of the considered species. To reduce those potential biases in the calculation, we therefore ran the model on a 100 years time span, although this duration was quite short compared to the longevity of the species (100 years represents less than 5 generations). However, there was still some heterogeneity in the predictions, due to the high natural variability of some biological characteristics of Chinese sturgeon. We therefore preferred to provide a range of predictions between upper and lower limits (see Fig. 2), than providing a-priori selection of the model parameters.
The VORTEX model used here predicted a decline of the population (Fig. 2 and Table 2). Such population trends were consistent with the results of the field survey showing a steep decline of fish recruitment (Chang 1999; Yi et al. 1999). This means that the conservation strategies, such as prohibiting commercial fisheries and restocking, are probably not sufficient to sustain the population. The main cause of the decline is primarily the loss of the natural spawning grounds due to the dam construction (Chang 1999; Wei 2003), which totally blocks the fish migration route (Xie 2003). These field observations were consistent with the model prediction showing a stabilization of the population in the case of the removal of the dams (Table 2). However, the consideration of dams removal for conservation purpose, although proposed by some authors (Kareiva et al. 2000; Dambacher 2001) is not a realistic issue in this case. On the contrary, the Chinese government policy favors construction of dams, such as the newly built TGD, located 38 km upstream from the Gerzhouba dam.
The actual main conservation strategy is represented by restocking using artificial reproduction techniques. Since 1999, more than 20000 young-of-the-year juveniles have been released each year to the Yangtze. The number of the released juveniles reached 60000 in 1999 (Zhu et al. 2002). Despite the important effort of the Chinese government, this conservation strategy hardly affects the decline of the population, as shown by Fig. 3b. This is in accordance with field works showing a low survival rate of juvenile sturgeons and therefore suggesting increasing supplementation (Chang 1999; Zhu et al. 2008; Wei 2003), as efficiently done for other sturgeon species. For example, millions of juveniles of Russian sturgeon (A. gueldenstaedti), stellate sturgeon (A. stellatus) and beluga sturgeon (Huso huso) were released each year in the Caspian Sea to sustain these species and compensate for population loss due to reduced spawning grounds, pollution, and commercial fishing (Secor et al. 2000; Ludwig 2008). However, such intensive supplementation requires capturing mature fish and the loss of adult fish used for artificial reproduction could not be compensated by juveniles restocking. Such artificial propagation using wild mature individuals should therefore be stopped immediately, as this conservation measure accelerates population declines. The best way to avoid this negative impact would be to stock juveniles and rear them in cages or ponds up to sexual maturity, and then use them for artificial reproduction, as proposed by Secor et al. (2000) and Wei et al. (2004) to conserve Caspian Sea sturgeons. This measure would have a lower impact than capturing adults, but previous experiments failed to induce gonads maturation of reared individuals. Another solution may be to improve artificial reproduction techniques to obtain more juveniles per female, as each female produce 645 000 eggs. This could permit to reduce the amount of mature fish capture and increase restocking. However, even if the artificial reproduction could be optimized, in that case, there will be a risk of genetic diversity loss due to outbreeding depressions (Ludwig 2006).
As actual conservation measures are not efficient, we tested two alternative scenarios. First, we tested the influence of the increase of spawning ground size, leading to increase the carrying capacity of spawning grounds, as it was efficiently used for other species in Russia and Canada (Vlasenko 1974; Provost et al. 1984). This measure, recommended for Chinese sturgeon by Chang and Cao (1999), did improve population size and population growth rate significantly in our simulations (Fig. 3d and Table 3). However, the actual spawning grounds are now endangered by the construction of the TGD. This dam will regulate Yangtze discharge, which will be reduced up to 41.6% during sturgeon spawning period (Chang and Cao 1999). Given the importance of flow fluctuations in sturgeon spawning, the TGD will strongly impact Chinese sturgeon reproduction. It is also likely to reduce the size of the spawning grounds due to decreasing water surface area and pebbles size, as experienced downstream of many dams (Baxter 1977; Allan 1996). The effect from TGD was modeled here by decreasing spawning ground area by 50%, which resulted in a significant decline of population growth rate and population size (Table 3). The second alternative scenario tested the effect of increasing sturgeon fecundity by reducing eggs predation by benthic fish. From a theoretical point of view, this could constitute a valuable conservation way, as more than 90% of the eggs are eaten by benthic fish predators directly in the spawning ground (Hu et al. 1992; Chang 1999). The model provided a highly significant positive response to reasonable decrease of predation pressure on eggs. An increase of 200% of the remaining eggs would stabilize the population to 13 000 individuals. From a practical point of view, it might consist of laying a fine meshed metallic grid on the spawning ground just after the end of spawning to reduce predation on eggs. Although hypothetical, such conservation measures should be tested.
Although PVA is just a predictive tool, with its inherent shortcomings, it clearly indicated a decline of the Chinese sturgeon population due to the loss of the spawning grounds. The results also suggested that the actual restocking strategy probably has a negative impact on the population. In addition, the model demonstrated that the efficient conservation strategies would be to build the spawning ground and protect sturgeon eggs against predation. However, the real impact of the newly built TGD has probably been underestimated in this model, as it may strongly modify the Yangtze ecosystem by regulating flow and reducing small particulate organic matter transport which will sink in the newly built reservoir. This may modify the aquatic ecosystem processes and therefore also affect fish diversity (Dudgeon 2000). In that way, the precise impact of the TGD on Chinese sturgeon is still difficult to predict. According to both field studies (Wei et al. 1997; Chang 1999; Yi et al. 1999) and to the model predictions, we can believe that the Chinese sturgeon urgently needs efficient conservation measures to avoid its extinction.
Although this work focuses on only one emblematic species, for which extended data is available, it should be noticed that Gerzhouba dam and the recently built TGD will also threaten many endemic species such as Yangtze sturgeon, Chinese paddlefish (Xie 2003), as well as more endemic fish species (Park et al. 2003). Although Wu et al. (2003) considered that the Gerzhouba and TGD dams could be viewed as an invaluable opportunity to study river fragmentation, it should be noticed that this “experiment” will cause a huge biodiversity decline in the Yangtze basin, as well as the extinction of several endemic fish species, among which, the Chinese sturgeon is only an example.
References
Allan JD (1996) Stream Ecology: structure and function of running waters. Chapman and Hall, London
Anonymous (1988) The biology of the sturgeon in Yangtze and their artificial propagation. The Yangtze Aquatic Resources Survey Group, Sichuan Scientific and Technical Publishing House, Chengdu (in Chinese)
Baxter RM (1977) Environmental effect of dams and impoundments. Annual Reviews of Ecology and Systematics 8:255–283
Brook BW (2000) Pessimistic and optimistic bias in population viability analysis. Conservation Biology 14:564–566
Brook BW, O'Grady JJ, Chapman AP, Burgman MA, Akcakaya HR, Frankham R (2000) Predictive accuracy of population viability analysis in conservation biology. Nature 404:385–387
Brook BW, Burgman MA, Akcakaya HR, O'Grady JJ, Frankham R (2002) Critiques of PVA ask the wrong questions: Throwing the heuristic baby out with the numerical bath water. Conservation Biology 16:262–263
Chang J (1999) Structure and dynamics of the spawning stock of Chinese sturgeon, Acipenser sinensis, in the Yangtze River. PhD Thesis. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan (in Chinese)
Chang J, Cao W (1999) History and prospect of conservation on Chinese sturgeon in the Yangtze River. Acta Hydrobiologica Sinica 23:712–720 (in Chinese)
Chen S, Watanabe S (1989) Age dependence of natural mortality coefficient in fish population dynamics. Nippon Suisan Gakkaishi 55:205–208
Coulson T, Mace GM, Hudson E, Possingham H (2001) The use and abuse of population viability analysis. Trends in Ecology and Evolution 16:219–221
Dambacher J (2001) Dam Breaching and Chinook Salmon Recovery. Science 291:939a
Deng Z, Yu Z, Xu Y, Zhou C (1985) Age determination and population structure of spawning Chinese sturgeon. Acta Hydrobiologica Sinica 9:99–110 (in Chinese)
Dudgeon D (2000) Large-Scale Hydrological Changes in Tropical Asia: Prospects for Riverine Biodiversity. BioScience 50:793–806
Ellner SP, Fieberg J, Ludwing D, Wilcox C (2002) Precision of population viability analysis. Conservation Biology 16:258–261
Fieberg J, Ellner SP (2000) When is it meaningful to estimate an extinction probability? Ecology 81:2040–2047
Fu Z, Hu D, Chen S (1985) Artificial propagation of the Chinese sturgeon below the Gezhouba Dam. Freshwater Fisheries 1:1–5 (in Chinese)
Gao X (2007) Conservation biology of rare and endemic fishes of the Yangtze River. PhD Thesis. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan (in Chinese)
Hu D, Ke F, Zhang G, Liu J (1992) Investigation on the spawning ground of Chinese sturgeon down Gezhouba Dam. Freshwater Fisheries 5:6–10 (in Chinese)
Kareiva P, Marvier M, McClure M (2000) Recovery and Management Options for Spring/Summer Chinook Salmon in the Columbia River Basin. Science 290:977–979
Lacy RC (1993) VORTEX: a computer-simulation model for population viability analysis. Wildlife Research 20:45–65
Lacy RC (2000) Structure of the VORTEX simulation model for population viability analysis. Ecological Bulletins 48:191–203
Lindenmayer DB, Possingham HP (1995) The conservation of arboreal marsupials in the Montame Ash Forests of the Central Highlands of Victoria. South-Eastern Australia. Modelling the sustainability of Leadbeater's Possum in response to modified timber harvesting practices. Biological Conservation 73:239–257
Ludwig D (1999) Is it meaningful to estimate a probability of extinction? Ecology 80:298–310
Ludwig A (2006) A sturgeon view on conservation genetics. European Journal of Wildlife Research 52:3–8
Ludwig A (2008) Identification of Acipenseriformes species in trade. J. Appl. Ichthyol. 24:2–19
Lunney D, O'Neill L, Matthews A, Sherwin WB (2002) Modelling mammalian extinction and forecasting recovery: koalas at Iluka (NSW, Australia). Biological Conservation 106:101–113
Mathews F, Macdonald DW (2001) The sustainability of the common crane (Grus grus) flock breeding in Norfolk: insights from simulation modelling. Biological Conservation 100:323–333
McCarthy MA, Andelman SJ, Possingham HP (2003) Reliability of relative predictions in population viability analysis. Conservation Biology 17:982–989
Miller P, Lacy RC (2005) VORTEX: a stochastic simulation of the extinction process. Version 9.50 user's manual. Conservation Breeding Specialist Group (SSC/IUCN), Apple Valley MN, USA. http://www.vortex9.org/vortex.html
Park YS, Chang J, Lek S, Cao W, Brosse S (2003) Conservation strategies for endemic fish species threatened by the Three Gorges Dam. Conservation Biology 17:1748–1758
Provost J, Verret L, Dumont P (1984) L'alose savoureuse au Quebec: synthese des connaissances biologiques et perspectives d'amenagement d'habitats. Rapport Manusrit Canadien des Sciences Halieutiques et Aquatiques 1793:86–87
Qiao Y, Tang X, Brosse S, Chang J (2006) Chinese sturgeon (Acipenser sinensis) in the Yangtze River: a hydroacoustic assessment of fish location and abundance on the last spawning groud. J. Appl. Ichthyol. 22(Suppl. 1):140–144
Reed JM, Mills LS, Dunning JB, Menges ES, McKelvey KS, Frye R, Beissinger SR, Anstett MC, Miller P (2002) Emerging issues in population viability analysis. Conservation Biology 16:7–19
Rieman BE, Allendorf FW (2001) Effective population size and genetic conservation criteria for bull trout. North American Journal of Fisheries Management 21:756–764
Secor DH, Arefjev V, Nikolaev A, Sharov A (2000) Restoration of sturgeons: lessons from the Caspian Sea Sturgeon Ranching Programme. Fish and Fisheries 1:215–230
Taylor BL (1995) The reliability of using population viability analysis for risk classification of species. Conservation Biology 9:551–558
Vlasenko AD (1974) On artificial spawning grounds for Kuban's sturgeons. Trudy Vniro. Moskva C II: 3–29
Waldman JR, Wirgin II (1997) Status and restoration options for Atlantic Sturgeon in North America. Conservation Biology 12:631–638
Wei Q (2003) Reproduction behavioral ecology of Chinese sturgeon (Acipenser sinensis Gray) with its stock assessment. PhD Thesis. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan (in Chinese)
Wei Q, Ke F, Zhang J, Zhuang P, Luo J, Zhou R, Yang W (1997) Biology, fisheries, and conservation of sturgeons and paddlefish in China. Environmental Biology of Fishes 48:241–255
Wei Q, He J, Yang D, Zheng W, Li L (2004) Status of sturgeon aquaculture and sturgeon trade in China: a review based on two recent nationwide surveys. J. Appl. Ichthyol. 20:321–332
World Commission on Dams (WCD) (2000) Dams and development: a new framework for decision-making. Earthscan, London
Wu J, Huang J, Han X, Xie Z, Gao X (2003) Three-Gorges Dam-experiment in habitat fragmentation? Science 300:1239–1240
Xiao H, Liu Y, Chang J (1999) Evaluation on status of artificial propagation and releasing of Chinese sturgeon in the Yangtze River. Acta Hydrobiologica Sinica 23:573–576 (in Chinese)
Xie P (2003) Three-Gorges Dam: risk to ancient fish. Science 302:1149
Yi J, Chang J, Tang D, Liu D, Ge B (1999) Present status of spawning stock of Chinese sturgeon, Acipenser sinensis, in the Yangtze River. Acta Hydrobiologica Sinica 23:554–559 (in Chinese)
Yu G, Liu J, Xu Y, Chang J (2002) Estimation on abundance of benthic fishes preying on eggs of Chinese sturgeon in reach below the Gezhouba Dam in the Yangtze River. Acta Hydrobiologica Sinica 26:591–599 (in Chinese)
Zhang S, Wang D, Zhang Y (2003) Mitochondrial DNA variation, effective female population size and population history of the endangered Chinese sturgeon, Acipenser sinensis. Conservation Genetics 4:673–683
Zhu B, Zhou F, Cao H, Shao Z, Zhao N, May B, Chang J (2002) Analysis of genetic variation in the Chinese sturgeon, Acipenser sinensis: estimating the contribution of artificially produced larvae in a wild population. Journal of Applied Ichthyology 18:301–306
Zhu B, Que Y, Yang Z, Chang J (2008) A review on genetic studies and their trade control in China. J. Appl. Ichthyol. 24(Suppl. 1):29–35
Acknowledgments
This study was supported in part by the key project of CAS (KZCX1-YW-08-01), the French-Chinese advanced research program (PRA E-02/3) and Executive Office of the State Council Three Gorges Project Construction Committee (SX [2001]-011). We thank R.C. Lacy for expert help on VORTEX modeling.
Author information
Authors and Affiliations
Corresponding author
Additional information
Note: Since 2006, Jianbo Chang has been transfered from Institute of Hydrobiology, Chinese Academy of Sciences to Institute of Hydroecology, Ministry of Water Resources and Chinese Academy of Sciences.
Appendix
Appendix
Comparison between two PVA models considering only fish older than 6 years and only fish older than 7 years. Models were run with 1 000 iterations and predicted population parameters in 10 years, 50 years, and 100 years, respectively. Population growth rate between simulations were compared using Student t-test. Differences in population growth rates between start age 6 and start age 7 were not significant (P > 0.05). Proportions of output population size to initial population size were similar between simulations. These results show that considering fish older than 6 years or fish older than 7 years did not significantly affect the results.
Rights and permissions
About this article
Cite this article
Gao, X., Brosse, S., Chen, Y. et al. Effects of damming on population sustainability of Chinese sturgeon, Acipenser sinensis: evaluation of optimal conservation measures. Environ Biol Fish 86, 325–336 (2009). https://doi.org/10.1007/s10641-009-9521-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-009-9521-4