
Abstract
Large trees (>76 cm breast-height diameter) are vital components of Sierra Nevada/Cascades mixed-conifer ecosystems because of their fire resistance, ability to sequester large amounts of carbon, and role as preferred habitat for sensitive species such as the California spotted owl. To investigate the likely performance of large trees in a rapidly changing climate, we analyzed growth rings of five conifer species against 20th century climate trends from local weather stations. Over the local station period of record, there were no temporal trends in precipitation, but maximum temperatures increased by 0.10 to 0.13 °C/decade (summer and autumn), and minimum temperatures increased by 0.11 to 0.19 °C/decade in all seasons. All species responded positively to precipitation, but more variation was explained by a significant positive response to minimum winter temperatures. High maximum summer temperature adversely affected growth of two species, and maximum spring temperatures in the year prior to ring formation were negatively associated with growth of one species. The strong coherent response to increasing minimum temperatures bodes well for growth of large trees in Sierra/Cascades region mixed conifer forest under continued climatic warming, but these trees will still be under threat by the increased fire intensity that is a indirect effect of warming.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
Large trees are vital elements of the fire-adapted interior conifer forests of northern California in part because they sequester sizable amounts of carbon while resisting fire and providing nesting habitat for the California spotted owl (Bias and Gutierrez 1992; Hurteau and North 2009). California public lands managers operationally define large trees as those having breast-height diameter >76 cm (USDA Forest Service 2004, p. 5). Understanding large-tree responses to a changing climate is essential for predicting future availability of the ecosystem services they provide, and is all the more pressing given the specter of widespread drought-related declines in forest productivity already documented in the American southwest (Williams et al. 2012). Analyses of tree responses to climate fluctuations in Sierran mixed-conifer forest have not distinguished large-tree response from that of smaller trees (Yeh and Wensel 2000) even though large, old trees may have differing response to climate fluctuations than smaller, younger trees (Mérian and Lebourgeois 2011; Vieira et al. 2009). Increases in global temperature are generally predicted to increase growth of temperate-zone trees (Ryan 2010), but trees from climates with Mediterranean-type seasonality, such as that of northern California, present a special case because of coinciding low rainfall and high growing-season temperatures (Gutiérrez 1991).
Growth of Californian lower montane conifers is positively correlated with cool-season precipitation and negatively correlated with summer temperatures in the year of and prior to ring formation (Yeh and Wensel 2000). Extrapolations of growth in future climates based on these findings suggest that tree growth will decrease under all plausible greenhouse gas emissions scenarios (Battles et al. 2008). Nevertheless, application of these findings to large conifers of the Sierra Nevada is uncertain because parameters were developed from young trees during a relatively brief (15-year) census period, and because the environmental correlates examined did not include non-summer temperatures. Even though summer is expected to be the main period of growth for Mediterranean-climate conifers, the temperature of other seasons may also be important.
There are specific reasons for expecting annual climatic differences of each season to affect tree growth. Temperature influences springtime leader growth and needle expansion (Kramer et al. 2000; Royce and Barbour 2001), and high early-season temperatures combined with dry or frozen soil can damage conifer vascular systems and subsequently reduce carbon uptake (Goldstein et al. 2000). High summertime temperatures, with consequent vapor pressure deficits, may deprive trees of carbon by forcing them to close stomata to maintain leaf turgor, and temperature exerts a diverse array of effects on photosynthetic and respiratory enzymes (Sage et al. 2008). High autumn temperatures can activate an alternative photo-protective system, inhibiting carbon uptake (Busch et al. 2008). Finally, winter photosynthesis is increasingly understood to contribute a substantial portion of the annual carbon budget of conifers of Mediterranean-climate ecosystems, and higher winter temperatures may increase carbon uptake because of decreased photoinhibition (Martínez-Ferri et al. 2004; Nippert et al. 2004). These studies suggest that conifers of Mediterranean-climate ecosystems should generally exhibit decreased growth with anomalously high spring, summer, or fall temperatures, or anomalously low winter temperatures, a pattern consistent with the findings of Fritts (1974) for arid-site conifers.
Our dendrological study focuses on large conifer trees at the juncture of the southern Cascades and northern Sierra Nevada mountain ranges in California USA, to determine the correlation of climate fluctuations with tree growth over a 110-year period beginning in the 20th century. We hypothesized that temperatures would influence species not just in summer but in all seasons. Our objectives were 1) to characterize the rate of change of temperature extremes and precipitation in the instrumental record for the study area, 2) to classify the response of a suite of large conifer trees to variation in seasonal temperature extremes and precipitation, and 3) to use (1) and (2) to infer likely short-term trajectories of large-tree growth in the study area.
2 Materials and methods
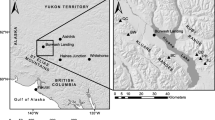
The study took place in Plumas County in northern California (Fig. 1), at the northern terminus of the Sierra Nevada range and the southern terminus of the Cascade range. The study sites consisted of Sierran mixed-conifer and ponderosa pine forest types at elevations ranging from 1,060 to 1,740 m. Characteristic tree species of these forests are white fir [Abies concolor (Gordon & Glend.) Lindl. ex Hildebr.], Douglas-fir [Pseudotsuga menziesii (Mirb.) Franco], incense-cedar [Calocedrus decurrens (Torr.) Florin], sugar pine (Pinus lambertiana Douglas), and ponderosa pine (Pinus ponderosa Douglas ex P. Lawson & C. Lawson). Prior to the era of intensive European-American settlement beginning in 1849, the forest had a disturbance regime of low-severity ground fires with a return interval of 8–22 years (Moody et al. 2006). Fire-suppression management began after 1910, and many forests were selectively harvested for large trees with extraction rates peaking from the 1960s through the 1980s. Understories are sparse in most areas; larger openings may contain shrubs such as greenleaf manzanita (Arctostaphylos patula Greene) and deerbrush (Ceanothus integerrimus Hook. & Arn.).

Location of sampled trees (black dots) and meteorological stations (crosses) within Plumas County (outline), California. Multiple trees were sampled at some sites
Although once common, large trees were heavily selectively harvested until the 1990’s and are now scarce and patchily distributed on the landscape. Owing to this scarcity we took advantage of large-tree location data from a contemporaneous study in which California spotted owls (Strix occidentalis occidentalis) were tracked to their nesting sites, which typically generally contain clusters of large trees at much higher density than the rest of the landscape (Bias and Gutierrez 1992). We selected sampling sites from among currently unoccupied nest sites that were close to access roads. Once at a site we attempted to locate a large (> 76 cm dbh) individual of each of the five study species, making sure measured trees were ≥ 20 m away from each other. Twenty or more individuals each of white fir, Douglas-fir, incense cedar, sugar pine, and ponderosa pine were selected (Fig. S1). Elevation, slope, and aspect were measured at the tree site as well as dbh. Two increment cores at right angles were extracted from each target tree at ~0.5 m above the ground. Cores were mounted in the field then prepared, measured back to the year 1900, and standardized using common dendrological techniques (see Electronic Supplementary Material).
Monthly precipitation and minimum and maximum temperature data were obtained from four weather stations within the sampled area (Fig. 1; URL: www.wrcc.dri.edu). Precipitation data were averaged at monthly intervals then summed based on a hydrologic year beginning in November. Temperature data were gap-filled, averaged at monthly intervals, extrapolated to 1,400 m elevation based on locally computed lapse rates, and averaged by season (Electronic Supplementary Material).
Climate effects on growth of each species were tested in a multiple linear regression to identify weather variables with the strongest influence on ring width index. Relative importance of climate variables was obtained by averaging across all possible orderings in the full 18-variable model (Grömping 2006), and variables that contributed ≥ 3.5 % to r 2 were retained for a regression based on a limited set (≤6) of variables. Variables with p < 0.05 in this regression were retained, and 95 % confidence intervals for them were obtained from 104 bootstrapped replicate datasets (Fox and Weisberg 2012). If confidence limits encompassed zero, variables were discarded. Diagnostics were performed by calculating the change in regression model parameters when observations were sequentially removed from the dataset; models were reviewed for continuing statistical significance (α < 0.05) of all parameters when removal of any observation caused >40 % decrease in the normalized value of any parameter estimate (Dalgaard 2002).
3 Results
Mean breast-height diameter of the 102 trees sampled ranged from 102 to 122 cm by species, and most were on southerly aspects with mean slopes of 32 to 38 % (Table 1).
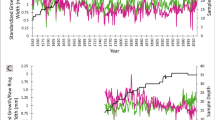
The seasonality of temperature and precipitation (composite record from 4 stations with modeled gap filling and extrapolation to 1,400 m elevation) was typical for Mediterranean-type climates, with average maximum summer temperatures of 27.5 °C and average minimum winter temperatures of −6.2 °C. Mean cool-season precipitation was 801 mm, mean warm season precipitation was 148 mm, and there were no linear trends in annual precipitation. Maximum air temperatures increased by 0.13 °C per decade in summer and 0.10 °C in winter but did not increase in other seasons, whereas minimum air temperatures increased linearly in all seasons. The rate of minimum temperature increase for winter (Fig. 2), spring, summer, and autumn all ranged from 0.10 °C to 0.19 °C per decade, but these rates did not differ statistically as shown by overlapping 95 % confidence intervals (Table 2).

Annual precipitation (Nov 1-Oct 31) and minimum winter air temperatures for 1900–2009, upper American River valley (composite of weather stations in Quincy, Chester, Greenville, and Canyon Dam)
Final regression equations for the main climate influences on growth contained two to four climatic factors with multiple r 2 of 17 %–40 % (Table 3, Fig. 3). The most striking pattern was a positive correlation between minimum winter temperatures in the year of ring formation and growth. All five species displayed this correlation, which explained between 7.6 % and 21.9 % of variation in ring width index. Surprisingly, white fir growth was negatively correlated with minimum winter temperatures in the year prior to ring formation. As expected, growth of all species was positively correlated with log-transformed precipitation in the year of and/or prior to ring formation. Growth of the two pines and Douglas fir was correlated with current-year precipitation, and growth of Douglas-fir, white fir, and incense cedar was correlated with prior-year precipitation. Growth of the latter two species was also weakly inhibited by high summer temperatures (weakness is inferred from small relative importance values). Only one species, ponderosa pine, had growth inhibited by high maximum spring temperatures in the year prior to ring formation.

Ring-width indices for five tree species of Sierran mixed-conifer forest in northern California for 1900 to 2009. Solid black line gives population means (residuals after removal of auto-correlation) and dotted line is prediction from multiple linear regression models incorporating only significant terms (coefficients in Table 3)
4 Discussion
4.1 Response to environmental factors
Strong correlation of tree growth with current- and/or previous-year precipitation is a general feature of Mediterranean-type climates (Gutiérrez 1991; Vieira et al. 2009), and therefore it was not surprising to detect a significant role of precipitation in the growth of the large trees in the present study. Our results were consistent with other studies of the same tree community in this finding (Sprague 2009; Yeh and Wensel 2000) with the minor difference that our data suggest that ponderosa pine, sugar pine, and Douglas-fir are mainly dependent on current-year precipitation, whereas white fir and incense cedar growth is correlated with precipitation in the year prior to ring formation. It is unclear whether there may be some functional linkage between lagged response to precipitation and growth inhibition by high summer temperatures, which coincidentally occurred only in white fir and incense cedar. Ponderosa pine is functionally classified as an isohydric species because its stomata respond rapidly to water deficits, allowing it to tightly regulate water stress (McDowell et al. 2008). White fir is more anisohydric than ponderosa pine, experiencing greater internal water stress under similar conditions (Barker 1973) because of lower stomatal sensitivity and greater ability to tolerate water stress without cavitation of root vessels (but see Grulke 2010; Lanini and Radosevich 1986; Martinez-Vilalta et al. 2004). Rapid stomatal response of ponderosa pine to water deficits is consistent with the observation that growth is correlated with current year precipitation, and conceivably the isohydric/anisohydric dichotomy may prove useful in predicting and interpreting growth in response to environmental factors over dendrological time scales. However, this must remain conjectural with respect to the current study, because of the paucity of ecophysiological information regarding sugar pine and incense cedar, and the high ecotypic variability of Douglas-fir.
High summer temperatures are often implicated in decreases in tree growth in Mediterranean climates (Gutiérrez 1991; Vieira et al. 2009), but we found only modest evidence of summer temperature influence on growth of large trees in the lower montane zone in the northern Sierra Nevada. Even in the two species with significantly decreased growth with higher summer temperatures, this correlation explained little variation in growth (<7 %) compared to other factors. A previous study of the same species over a larger area but shorter period (15 years) found a more pervasive influence of high summer temperatures, although like the present study they found that ponderosa and sugar pine were less sensitive to high temperatures than the more shade-tolerant white fir, Douglas-fir, and incense cedar (Yeh and Wensel 2000). Some of the difference in findings could be due to differences in the size and age of individuals sampled in the two studies, i.e., young vs. old trees in the present study. We conjecture that the roots of large trees may have reached deeper sources of water than the roots of the young stands studied by Yeh and Wensel (2000), allowing continued transpiration and carbon uptake in the face of high vapor pressure deficits caused by high temperatures (Kerhoulas et al. 2013). Nevertheless, larger and older individuals of these species may be more sensitive to climate than younger individuals (Sprague 2009).
High maximum spring temperatures in the year preceding ring formation may have limited growth of ponderosa pine. Early spring drying events (vapor pressure deficits from high temperatures combined with low atmospheric humidity) in soil that is frozen or dry can lead to vascular embolism and subsequent reduction in carbon fixation throughout the summer/autumn growing season (Goldstein et al. 2000). Such reductions in carbon fixation may then be expressed in unusually low stem growth the following spring. Meteorological inputs for our analysis did not include vapor pressure deficit because we lacked local measurements of dew point (an indicator of atmospheric humidity) for the early decades of the 110-year meteorological record. Nevertheless vapor pressure deficit has proved highly correlated with arid-site tree performance (Williams et al. 2012), and incorporating vapor pressure deficit would be a logical step for modeling growth in recent decades at our study, particularly because several large growth deviations at decadal time scales were not effectively captured by our modeling approach (Fig. 3).
A coherent positive response to higher winter minimum temperatures among all five tree species is the most striking finding of the present study (negative correlation of white fir growth to higher minimum temperatures the year after ring formation was an exception to the coherence of the response). Indeed, the tree community responds even more strongly to minimum winter temperatures than to precipitation, as judged by summing relative importance values for these factors across species (Table 3). The importance of minimum winter temperatures for growth of large individuals of Sierran lower montane conifers has not been highlighted in previous studies, although Fritts (1974) observed that a direct relationship between growth and winter temperature is frequently seen for arid-site conifers. Winter usually is viewed as a quiescent time for conifers and physiological research has focused on protective mechanisms that guard against photo-oxidative damage to green needles after down-regulation of photosynthesis. Nevertheless, winter is increasingly recognized as an important time for photosynthesis in conifers, particularly in Mediterranean-climate regions (Martínez-Ferri et al. 2004; Misson et al. 2006; Nippert et al. 2004). Recent findings have emphasized the importance of transient photoinhibition/up-regulation involving the xanthophyll cycle in conifers in climates that experience rapid change in temperature. The duration of photo-inhibition subsequent to exposure to freezing temperatures is an active area of research. Given that mean monthly winter maximum and minimum temperatures at our study site are approximately symmetrical around 0 °C, transient responses to freezing temperatures may play a large role in winter carbon balance.
4.2 Future climate and tree growth
The identification of relationships between seasonal variation in climate variables and annual growth raises implications of projected future climate on tree growth. McKenzie et al. (2001) advocate against extrapolation to long-term climatic forcing because climate may influence growth over multiple scales and because climate-growth relationships can change as trees adapt phenotypically over time. Circumspection is clearly required in making extrapolations, yet it is reasonable to at least qualitatively examine future climate predictions in light of short-term tree growth responses. Most global climate models predict a poleward expansion of the subtropical dry zones during the current century, but at 40° latitude the upper American River valley, the site of the this study, is expected to lie between a zone of greater drying to the south and increased wetness to the north and is expected to escape major changes in precipitation quantity (Seager and Vecchi 2010). Our results therefore do not augur precipitation-driven changes in growth rates. In contrast to precipitation, minimum winter temperatures have risen in northern California over the 20th century (Hamlet and Lettenmaier 2007; this paper), and mean winter and summer temperatures are predicted to continue to rise through the current century (Cayan et al. 2006). Our results suggest that this may stimulate growth of large conifers, although the growth of two of the five species we studied may be partly counterbalanced by negative responses to increasing summer maximum temperatures. The findings do not support projections of uniformly downward growth trends under foreseeable climate scenarios (Battles et al. 2008).
4.3 Conclusions
Minimum winter temperatures have been a major determinant of growth of large trees in the lower montane mixed-conifer forest in northern California over the 20th century, eclipsing even precipitation in importance. The five species studied show increased growth with higher minimum winter temperatures, a pattern that was stronger and more prevalent than decreasing growth with higher summer maximum temperatures. Large trees are an iconic and functionally irreplaceable element of the lower montane mixed-conifer forest, and the steady rise in surface temperature in California throughout the 20th century has rightfully engendered concern about their performance, raising the specter of a positive feedback loop in which elevated temperatures due to anthropogenic carbon emissions inhibit the ability of trees to take up carbon. Our findings may help to allay such fears as regards large trees. Vigilant forest management will nonetheless be required to protect and recruit large trees, which despite their exceptional fire resistance are still vulnerable to the increasingly large, intense, and frequent fires characteristic of present-day western conifer forests.
References
Barker JE (1973) Diurnal patterns of water potential in Abies concolor and Pinus ponderosa. Can J For Res 3:556–564
Battles JJ, Robards J, Das AJ, Waring K, Gilless JK, Biging GS, Schurr F (2008) Climate change impacts of forest growth and tree mortality: a data-driven modeling study in mixed-conifer forest of the Sierra Nevada, California. Clim Chang 87:193–213
Bias MA, Gutierrez RJ (1992) Habitat associations of California spotted owls in the central Sierra Nevada. J Wildl Manag 56:584–595
Busch F, Hüner NPA, Ensminger I (2008) Increased air temperature during simulated autumn conditions impairs photosynthetic electron transport between photosystem II and photosystem I. Plant Physiol 147:402–414
Cayan DR, Maurer EP, Dettinger MD, Tyree M, Hayhoe K (2006) Climate change scenarios for the California region. Clim Chang 87:21–42
Dalgaard P (2002) Introductory statistics with R. Springer, New York
Fox J, Weisberg S (2012) Bootstrapping regression models in R. An R companion to applied regression, 2nd edn. Sage, Thousand Oaks, p 17
Fritts HC (1974) Relationships of ring widths in arid-site conifers to variations in monthly temperature and precipitation. Ecol Monogr 44:411–440
Goldstein AH, Hultman NE, Fracheboud JM, Bauer MR, Panek JA, Xu M, Qi Y, Guenther AB, Baugh W (2000) Effects of climate variability on the carbon dioxide, water, and sensible heat fluxes above a ponderosa pine plantation in the Sierra Nevada (CA). Agric For Meteorol 101:113–129
Grömping U (2006) Relative importance for linear regression in R: the package relaimpo. J Stat Softw 17
Grulke NE (2010) Plasticity in physiological traits for conifers: implications for response to climate change in the Western U.S. Environ Pollut 158:2032–2042
Gutiérrez E (1991) Climate tree-growth relationships for Pinus uncinata Ram. in the Spanish pre-Pyrenees. Acta Oecol 12:213–225
Hamlet AF, Lettenmaier DP (2007) Effects of 20th century warming and climate variability on flood risk in the western U.S. Water Resour Res 43
Hurteau M, North M (2009) Fuel treatment effects on tree-based forest carbon storage and emissions under modeled wildfire scenarios. Front Ecol Environ 7:409–414
Kerhoulas LP, Kolb TE, Koch GW (2013) Tree size, stand density, and the source of water used across seasons by ponderosa pine in northern Arizona. For Ecol Manag 289:425–433
Kramer K, Leinonen I, Loustau D (2000) The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems: an overview. Int J Biometeorol 44:67–75
Lanini WT, Radosevich SR (1986) Responses of three conifer species to site preparation and shrub control. For Sci 32:61–77
Martínez-Ferri E, Manrique E, Valladares F, Balaguer L (2004) Winter photoinhibition in the field involves different processes in four co-occurring Mediterranean tree species. Tree Physiol 24:981–990
Martinez-Vilalta J, Sala A, Pinol J (2004) The hydraulic architecture of Pinaceae - a review. Plant Ecol 171:3–13
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb TE, Plaut JA, Sperry JS, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178:719–739
McKenzie D, Hessl AE, Peterson DL (2001) Recent growth of conifer species of western North America: assessing spatial patterns of radial growth trends. Can J For Res 31:526–538
Mérian P, Lebourgeois F (2011) Size-mediated climate–growth relationships in temperate forests: a multi-species analysis. For Ecol Manag 261:1382–1391
Misson L, Gershenson A, Tang J, McKay M, Cheng W, Goldstein A (2006) Influences of canopy photosynthesis and summer rain pulses on root dynamics and soil respiration in a young ponderosa pine forest. Tree Physiol 26:833–844
Moody TJ, Fites-Kaufman J, Stephens SL (2006) Fire history and climate influences from forests in the northern Sierra Nevada, USA. Fire Ecol 2:115–141
Nippert JB, Duursma RA, Marshall JD (2004) Seasonal variation in photosynthetic capacity of montane conifers. Funct Ecol 18:876–888
Royce EB, Barbour MG (2001) Mediterranean climate effects. II. Conifer growth phenology across a Sierra Nevada ecotone. Am J Bot 88:919–932
Ryan MG (2010) Temperature and tree growth. Tree Physiol 30:667–668
Sage RF, Way DA, Kubien DS (2008) Rubisco, rubisco activase, and global change. J Exp Bot 59:1581–1595
Seager R, Vecchi GA (2010) Greenhouse warming and the 21st century hydroclimate of southwestern North America. Proc Natl Acad Sci 107:21277–21282
Sprague A (2009) Evaluating the effect of climate in a Sierra mixed conifer forest. Berkeley Sci J 13:44–53
USDA Forest Service (2004) Record of decision: Sierra Nevada forest plan amendment final supplemental environmental impact statement. Management Bulletin. U.S. Forest Service, Pacific Southwest Region, Vallejo, California
Vieira J, Campelo F, Nabais C (2009) Age-dependent responses of tree-ring growth and intra-annual density fluctuations of Pinus pinaster to Mediterranean climate. Trees Struct Funct 23:257–265
Williams AP, Allen CD, Macalady AK, Griffin D, Woodhouse CA, Meko DM, Swetnam TW, Rauscher SA, Seager R, Grissino-Mayer HD, Dean JS, Cook ER, Gangodagamage C, Cai M, McDowell NG (2012) Temperature as a potent driver of regional forest drought stress and tree mortality. Nat Clim Chang 3:292–297
Yeh H-Y, Wensel LC (2000) The relationship between tree diameter growth and climate for coniferous species in northern California. Can J For Res 30:1463–1471
Acknowledgments
Kaline Le Coq and J. Hulbert did field work, and S. Livingston supervised and did lab work. Paula Shaklee, C. Gallagher, and J. Keane assisted with finding large trees, and P. Stine and US Forest Service International Programs provided administrative assistance. Three reviewers provided input that greatly improved the paper. We thank A. Lloyd for discussion and US Forest Service Region 5 for funding.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 881 kb)
Rights and permissions
About this article
Cite this article
Bigelow, S.W., Papaik, M.J., Caum, C. et al. Faster growth in warmer winters for large trees in a Mediterranean-climate ecosystem. Climatic Change 123, 215–224 (2014). https://doi.org/10.1007/s10584-014-1060-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-014-1060-0