
Abstract
The threats of wide-scale coral bleaching and reef demise associated with anthropogenic (global) climate change are widely known. Less well considered is the contributing role of conditions local to the reef, in particular reef water quality, in co-determining the physiological tolerance of corals to increasing sea temperatures and declining pH. Here, the modelled benefit of reduced exposure to dissolved inorganic nitrogen (DIN) in terrestrial runoff, which raises the thermal tolerance of coastal coral communities on the central Great Barrier Reef (Australia), is considered alongside alternative future warming scenarios. The simulations highlight that an 80% reduction in DIN ‘buys’ an additional ~50–60 years of reef-building capacity for No Mitigation (‘business-as-usual’) bleaching projections. Moreover, the integrated management benefits provided by: (i) local reductions of ~50% in DIN contained in river loads, and (ii) global stabilisation of atmospheric CO2 below 450 ppm can help ensure the persistence of hard-coral-dominated reefscapes beyond 2100. The simulations reinforce the message that beyond the global imperative to mitigate future atmospheric CO2 emissions there still remains the need for effective local management actions that enhance the resistance and resilience of coral reef communities to the impacts of climate change.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Anthropogenic-driven climate change is very likely to warm the surface waters of the Great Barrier Reef (GBR) at an unprecedented rate throughout the 21st Century (CSIRO and BoM 2007). The rising sea surface temperatures (SST) are predicted to cause increased coral bleaching and coral mortality, with serious consequences for the Reef’s biodiversity, ecology, appearance and dependent recreational use and economic activity (reviewed in Johnson and Marshall 2007). The severity of the problem is highlighted by predictions that coastal reefs in the central GBR area may be severely set back or even transformed to non-coral dominated states by as early as 2030 (Wooldridge et al. 2005). These risks are likely to be exacerbated by emerging ocean chemistry changes arising from the absorption of anthropogenic CO2 into surface ocean waters (Sabine et al. 2004). So-called ocean acidification may increase the susceptibility of corals and coral reef organisms to temperature stress, reduce productivity and disrupt the process by which coral produce their calcium carbonate skeletons (Kleypas et al. 2006; Anthony et al. 2008; Veron et al. 2009).
Given these unprecedented threats, a fundamental challenge for sustainable reef management is to identify policy and management interventions that might help to safeguard the reef system against long-term (and potentially irreversible) negative changes to its structure and dynamics. In principle, these could include local actions that improve the survivability of coral communities and/or aid their post-disturbance rate of recovery. However, ecological model simulations for the GBR suggest that improved recovery alone is unlikely to sustain hard-coral-dominated reefscapes beyond 2050 for anything except the most optimistic (low) rates of future warming (Wooldridge et al. 2005). Success in maintaining resilient reef communities under higher (and more likely) rates of future warming thus appears ultimately tied to also improving coral survival rates from thermal bleaching.
Present upper thermal bleaching thresholds of corals are the result of long-term adaptation of the coral-zooxanthellae symbiosis to its local thermal regime (see e.g., Fitt et al. 2001; Coles and Brown 2003). There is variability in heat resistance among strains of zooxanthellae (e.g., Baker et al. 2004), and some evidence that the relative proportions of heat-resistant strains can vary following heat stress events (Berkelmans and van Oppen 2006). However, present indications are that the speed and persistence of shifts is unlikely to compensate for the trend of increasing temperature stress predicted over the course of this century (see e.g., Coffroth et al. 2010; LaJeunesse et al. 2010). Further, there are no indications that local management interventions could be used to change the symbiotic composition at appropriate time and space scales to raise reef-wide thermal bleaching resistance.
Management interventions can, however, affect other characteristics of the system that influence thermal tolerance. Many efforts to improve the health of reef ecosystems focus on improving water quality, and recent reports suggest that poor water quality may exacerbate effects of heat stress by lowering the thermal bleaching threshold of certain coral species (Wooldridge and Done 2009; Wooldridge 2009a; Carilli et al. 2009a, b; Wagner et al. 2010; Faxneld et al. 2011). For example, the post-1980’s increase in bleaching intensity and frequency at a Mesoamerican reef was best explained by reduced water quality in the region due to a substantial increase in coastal development (Carilli et al. 2009a, b). Similarly in Jamaica, where underwater ecological observations date to the 1950s, mass bleaching went unreported until 1987 (Goreau 1992), after coastal water quality had declined, despite similar thermal stress in 1957 and 1958 to that in 1987 (Carilli et al. 2009a). Finally, corals on GBR reefs that are regularly exposed to poor water quality appear to have a higher bleaching sensitivity (per unit increase in SST) than those on less or unexposed reefs (Wooldridge and Done 2009).
Dissolved inorganic nitrogen (DIN = nitrate + nitrite + ammonium) is a key water quality attribute affecting the stability of the coral-zooxanthellae symbiosis, and it has been proposed that elevated ambient DIN concentrations exacerbate the damage to cellular processes that underpin thermal bleaching (Wooldridge 2009b; Wooldridge and Done 2009). This mechanistic explanation is consistent with results of experimental enrichment of seawater with DIN, wherein the temperature-dependent loss of zooxanthellae was increased (Schlöder and D’Croz 2004; Baohua et al. 2004; Kuntz et al. 2005; Haas et al. 2009). In these studies, DIN concentrations as low as 1–10 μM alone increased zooxanthellae loss, even at normal ambient summer temperatures. Field measurements indicate that summer DIN concentrations at specific reefs on the Florida Keys (USA) often exceed 1 μM (typically due to terrestrial waste-water discharge) and bleach more severely during periods of thermal stress (Wagner et al. 2010). Similarly, a strong quantitative relationship exists between the bleaching threshold of inshore corals on the GBR and the degree of exposure to DIN-rich (>1–10 μM; Devlin et al. 2001) flood-plume (terrestrial) waters (Fig. 1a; Wooldridge 2009a). In this case, the variable water quality regime spans ~2°C variation in bleaching ‘resistance’, with the marginal rate of increased resistance (°C) being significantly higher at the lowest DIN exposure levels; i.e., sites with good water quality (Fig. 1b).

a Quantitative linkage between upper thermal bleaching limits (°C) and the degree of exposure to nutrient enriched terrestrial waters (after Wooldridge 2009a). Coastal reef waters with high DIN-enriching impact are characterised by a higher annual exceedence probability (AEP) of [Chl a] > 0.9 μg.L−1; a threshold (trigger) value that correlates with localised reductions in hard coral species richness on the GBR (De’ath and Fabricius 2010). Implicit with the bleaching limits is the exceedence of a short (<5 day) exposure period b Marginal (bar) and cumulative (line) increase in coral bleaching threshold (°C) across the (normalised) inshore DIN-enrichment gradient. The relationship demonstrates that water quality improvements at relatively clean sites will attract the greatest initial improvements in bleaching threshold
Based on the evidence detailed in the above studies, this study assumes that a quantitative causal relationship between terrestrial DIN loading and thermal bleaching thresholds exists. This highlights the particularly crucial role of management activities that improve water quality as a way of increasing the probability that corals will tolerate predicted temperature increases. In anticipation of increasing interest in management of water quality as a way of building reef resilience to climate change, we have developed a modelling framework to map coral bleaching risks at terrestrially impacted (i.e. coastal) reefs as a function of two key drivers: local land management and global warming. Here, we develop and apply this modelling framework to support management decision making that aims to build climate change resilience of reefs through spatially targeted water quality management. Specifically, we: (i) simulate the potential improvement in bleaching tolerance (°C) of locally-impacted corals due to 20, 40, 60 and 80% reductions in the end-of-river DIN loads for the major basins that drain the GBR catchment, and, given a range of future warming scenarios, (ii) project the benefit of this improved thermal tolerance on coral survival rates to bleaching.
The central GBR represents an ideal study area to highlight important aspects of the modelling framework. Flood-plume sampling within the central GBR lagoon demonstrates that DIN is well mixed (= conservatively diluted) throughout the plume volume (Devlin et al. 2001). The DIN-enriching footprint of terrestrial runoff intrusions into the GBR therefore increases considerably as end-of-river DIN concentrations rise. Current modelling indicates that the estimated 3–5 times increase in end-of-river DIN concentrations since European settlement (Furnas 2003) has increased the area of DIN-enriched waters in the central GBR by a factor of ~10–20 (Wooldridge et al. 2006). This has extended nutrient-enriched coastal waters typically bathing near-shore reefs (with characteristic corals and macro-algae zones—Done 1982; DeVantier et al. 1998) to mid- and even outer-shelf reef locations (characterized by other corals and coralline algae—Wooldridge et al. 2006; Maughan and Brodie 2009).
2 Methods
2.1 Water quality-dependent coral bleaching thresholds
The decision support tool ‘ChloroSim’ (Wooldridge et al. 2006) was used to model the beneficial effects of riverine DIN reductions, relative to present day, on the size and intensity of the enriching footprint of flood plumes within the GBR lagoon. ChloroSim is based on a regionally-calibrated relationship that links: (i) end-of-river DIN concentrations, (ii) runoff: seawater dilution (a measure of flood intensity), and (iii) the ensuing flood plume intensity (‘bloom’) of phytoplankton biomass (as indicated by the concentration of photosynthetic pigment, chlorophyll a (Chl a)). The general rationale behind ChloroSim is that for a given runoff: seawater dilution ratio, any broad-scale differences in the plume concentration of Chl a can be largely attributed to the initial concentration of DIN in the discharging runoff. In this way, simulated reductions (20%, 40%, 60% and 80%) in end-of-river DIN concentration were processed through a 30-year (1969–1998) runoff: seawater flood plume archive for the GBR (King et al. 2002); thereby enabling the annual exceedence probability (AEP) for [Chl a > 0.9 μg/L] to be calculated for every 2 km pixel in the coastal impact model domain. The potential degree of improvement in thermal tolerance (°C) of coastal coral communities due to the specified (uniform%) reductions in the end-of-river DIN loads was computed for each pixel by substituting this AEP value within the water quality-dependent bleaching threshold relationship developed by Wooldridge (2009a) (see Fig. 1).
2.2 Projected SST warming patterns till 2100
The future SST warming pattern on the GBR (till 2100) was considered for two alternative global CO2 emission scenarios: (a) No mitigation (‘business-as-usual’), and (ii) CO 2 mitigation leading to atmospheric stabilisation of CO2 concentrations at 450 ppm early in the 22nd century (after Garnaut 2008). The translation of global CO2 scenarios into regional-scale SST estimates was facilitated by the ‘ReefClim’ software package. ReefClim adopts a downscaling methodology based on analysing the results of a number of global climate models to obtain regional patterns of maximum summer SST change expressed in terms of local warming per degree of global warming (Mitchell 2003; Whetton et al. 2007). The methodology attempts to incorporate the modifying impact of local weather and ameliorating oceanographic processes.
For the present study, eight different climate models were used to develop an envelope (distribution) of maximum summer month SST estimates, and included representations from the Hadley Centre (two models), Max Planck Institute (two models), Canadian Climate Centre (two models), and CSIRO (two models). Additional details of the models are provided in supplemental Table S1. The use of a number of climate models means that uncertainty due to model differences in the pattern of SST warming is allowed for. As there was no way to assess which model warming pattern was most likely, the range achieved for any 10-km grid cell location was considered representative of possible changes.
2.3 Coral mortality (LD50) thresholds
The SST projections were combined with coral mortality thresholds under different ambient DIN to project the course of temperature driven coral mortality (LD50) until 2100. Mortality threshold curves based on 50% mortality of thermally sensitive coral taxa have been proposed by Berkelmans (2009). An analysis of these curves indicates that most thermally sensitive taxa die at temperatures ~0.5–1.5°C above their bleaching threshold. For the present study, a modelled trigger value of 3°C above the water quality dependent bleaching threshold was chosen to represent 50% mortality events (LD50). The higher triggering point endeavours to represent the raised ‘GBR-wide’ average thermal tolerance due to the contribution of more thermally-tolerant coral taxa (e.g. massive Porites spp; Marshall and Baird 2000). This reef-average mortality threshold compares favourably with the ~2–3°C difference between field-measured bleaching (BD50) and mortality (LD50) observations during the 1998 and 2002 mass bleaching events on the GBR (Wooldridge 2008).
Calculation of the AEP of the LD50 threshold (for each level of DIN improvement) was achieved with a standard normal (z-score) methodology. In this case, at specific time intervals (2010, 2030, 2050, 2070, 2100) the downscaled SST projections were used to develop a normal distribution of daily summer-averaged SST projections for each 10-km grid cell that constitutes the riverine (flood) impacted GBR model domain. With the z-score approach, the likelihood of exceeding the spatially explicit LD50 threshold can be conceptualised as the area under the curve in excess of the threshold, z (supplementary Fig. A1). The standard deviation (SD) of this distribution for each grid cell was calculated based on a 12-year remotely-sensed climatology (AIMS SST web atlas; Skirving et al. 2002) of maximum SST during the summer (Dec-Mar) period. These SD estimates were temporally updated based on the assumption that the coefficient of variation of SST was stationary in time. Thus for each grid cell, the mean SST increases over time but the coefficient of variation around this value is assumed temporally uniform.
3 Results
3.1 Simulated improvement in local coral bleaching thresholds
The potential degree of improvement in thermal tolerance (°C) of coastal coral communities due to the specified (uniform%) reductions in the end-of-river DIN loads is presented in Fig. 2. These simulations indicate that the potential ~2°C improvement in bleaching threshold for the most impacted reef sites typically require large (>50%) reductions in DIN. Some care is needed in the practical interpretation of Fig. 2 since the uniform reductions (%) in end-of-river DIN concentration are based on pre-existing river (flood) loads. For example, to achieve a 40% reduction in the end-of-river DIN concentration from a ‘nutrient-rich’ river system requires a substantially larger absolute reduction in DIN (in terms of μg.L−1) than a 40% reduction from a ‘nutrient-poor’ river system. Nevertheless, standardising the model output does help to demonstrate the higher marginal rate of improvement (°C) at progressively cleaner DIN sites. This is observable at two scales in Fig. 2: (i) the north to south gradient of increasing end-of-river DIN concentrations (Furnas 2003), and (ii) the transitions from the more diluted plume-edges to individual river mouths. Such spatial patterns suggest that it will be hardest to achieve DIN-related improvements in thermal bleaching tolerance at reef sites closest to the most enriched river mouths.

Simulated increased in the upper thermal bleaching limits (°C) of impacted corals due to specified (uniform%) reductions in end-of-river DIN loading for the numerous basins that drain the GBR catchment
3.2 Simulated warming of local coral reef waters
Figure 3a illustrates the projected rate of future global warming (2000–2100) based on: (i) no CO2 emission mitigation (‘business-as-usual’), and (ii) global CO2 emission mitigation leading to stabilisation of atmospheric CO2 at 450 ppm (after Garnaut 2008). The linked change in regional SST for the GBR, expressed as change per degree of global warming, is highlighted in Fig. 3b. Beyond the inshore (shallow) to offshore (deep) enhancement, the dominant feature of the projected SST warming on the GBR is the proportionately higher rate of warming in the central-southern GBR. In considering the robustness of these predicted patterns, it is important to note that despite our best efforts to incorporate local variability at the scale ~10’s km (e.g., due to major currents, eddies and basic bathymetry) the warming projections are unlikely to capture the potential modifying influence of reef scale (~1 km) bathymetry and hydrodynamics. The choice of a conservative reef-scale mortality threshold attempts to offset this potential uncertainty.

a Projected global warming (2000–2100) based on: (i) no CO2 emission mitigation (‘business-as-usual’), and (ii) global CO2 emission mitigation leading to stabilisation of atmospheric CO2 at 450 ppm (after Garnaut 2008) b Regional SST changes for the GBR expressed as change per degree of global warming based on the average of eight climate models
3.3 Projected coral mortality (LD50) till 2100
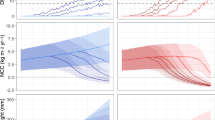
Output for a representative inshore reef area of the central GBR is shown in Fig. 4, highlighting the potential levels of improvement in the AEP of LD50 events given the simulated improvement (20%, 40%, 60% and 80%) in end-of-river DIN loads relative to present day values. To aid interpretation, a 10-yr average return interval (ARI) is highlighted (grey line), which provides an estimate of the lowest possible ARI estimate for maintaining a viable, hard-coral-dominated reef condition (see Discussion). Importantly, the model projections demonstrate that for this representative area, an 80% reduction in DIN permits the maintenance of the coral-dominated reef state for an additional ~50 years beyond the current LD50 mortality projections for the No mitigation (‘business-as-usual’) warming scenario. Moreover, when coupled with the CO 2 mitigation (‘Stabilisation at 450 ppm’) warming scenario, the enforcement of ~50% reduction in DIN is sufficient to ensure the long-term persistence of the impacted reefs.

Projected annual exceedence probability (AEP) for LD50 mortality events given a 20%, 40%, 60% and 80% reduction in end-of-river DIN load. The projections are run for the No mitigation (‘business-as-usual’) and CO 2 mitigation (‘Stabilisation at 450 ppm’) global warming scenarios. The 10 year ARI (grey line) is indicative of the maximum bleaching disturbance frequency for the maintenance of a viable hard-coral-dominated reefscape. The results highlight that as an absolute minimum, local reductions in DIN load of ~50% and global CO2 stabilisation at ~450 ppm are needed to ensure the long-term persistence of coastal reefs on the central GBR. Note: (ARI ≈ 1/AEP)
3.4 Timeframes for the loss of viable reef communities
Extrapolation of the modelling framework across the entire flood-impacted GBR domain allows the likely timeframe (yr) for the loss of viable hard-coral-dominated reef communities to be projected for No mitigation (Fig. 5) and CO 2 mitigation (Fig. 6) warming scenarios. The key findings are: (i) without any DIN reduction the long-term viability of a large proportion of inshore reef area is already marginal given the existing bleaching risk profile (2010), and (ii) local reductions in DIN loads of ~50% and global atmospheric CO2 stabilisation below 450 ppm allows impacted reefs to persist beyond 2100, thus providing a potential integrated management target.

Projected timeframes (yr) for the loss of viable reef communities based on the No Mitigation (‘business-as-usual’) warming scenario and alternative levels of (uniform%) reductions in end-of-river DIN loads. Projections are based on the suggested minimum 10 year ARI between catastrophic LD50 mortality events

Projected timeframes (yr) for the loss of viable reef communities based on a CO 2 Mitigation (‘Stabilisation at 450 ppm’) warming scenario and alternative levels of (uniform%) reductions in end-of-river DIN loads. Projections are based on the suggested minimum 10 year ARI between catastrophic LD50 mortality events
4 Discussion
Maintenance of reef framework and reef building capacity are important attributes of healthy functioning coral reefs (Davies 1983; Done et al. 1996). An important parameter that helps to define reef-building capacity is the average return interval (ARI) between events that cause catastrophic mortality in reef-building taxa (see e.g., Connell 1997; Done 1999). Long-term field studies on the GBR highlight that even for sites with few local stressors, an ARI between major disturbance events of at least 10 years is critical for maintaining hard-coral-dominated reefscapes (see e.g., Wakeford et al. 2008). Worryingly, our modelling shows the present day (2010) bleaching frequency experienced by many coastal reef communities on the central GBR is already likely to exceed their long-term capacity to maintain reef-building capacity; i.e., the viability of many reefs is already marginal before considering scenarios for future coral bleaching and ocean acidification. This finding is supported by a recent GBR-wide meta-analysis of inshore benthic community change data covering 1985–2007, which concludes that thermal bleaching frequency and severity over the last two decades have caused a rapid decline in inshore coral cover (Thompson and Dolman 2010). The authors explain that whilst significant cover of live coral presently remains on the inshore reefs, and that recovery is observed during inter-disturbance periods, the system will not be resilient to an equivalent or more severe disturbance regime over the long-term. Done et al. (2007) reach a similar conclusion based on repeat analysis of photogrammetric surveys (1981–2005) taken from a coastal reef on the central GBR.
Significant to the present study is the considered impact of post-European declines in local water quality, specifically via terrestrial DIN-enrichment, as a contributing influence in lowering the bleaching tolerance (i.e. enhancing the bleaching risk profile) of impacted corals (sensu Wooldridge and Done 2009; Wooldridge 2009a). Previously, the role of water quality in altering the resilience of coral communities under climate change has primarily focused on recovery rates of coral from mortality events (see review by McCook et al. 2007). For example, reef waters characterised by low sediment and nutrient loads are judged favourably in terms of: (i) promoting the re-establishment of disturbed reef sites with new coral recruits by improving success rates for larval arrival, settlement, post-settlement survival, and growth; and (ii) limiting the potential for faster growing seaweeds to out-compete surviving (remnant) corals and new coral recruits. Moreover, since high levels of turbidity and nutrients can impede the skeletal growth rate and reproductive output of adult corals (reviewed by Fabricius 2005), good water quality is an essential ingredient for enhancing reef-building capacity between disturbance events. Whilst these recovery-side benefits are a fundamental driver for investment in water quality improvement, the present study highlights that enhanced bleaching survival is also an important consideration.
Indeed, our simulations suggest that reducing end-of-river DIN loads can considerably benefit the future survival prospects of locally-impacted reefs on the GBR. For example, a simulated 80% reduction in DIN ‘buys’ an additional ~50–60 years of reef-building capacity beyond current (No mitigation) bleaching projections (Fig. 5), whilst a ~50% reduction in DIN sustains long-term (>2100) reef-building capacity under a global CO 2 mitigation (‘Stabilisation at 450 ppm’) future warming scenario (Fig. 6). Importantly, a simplifying assumption within the present analysis is that the inter-disturbance recovery rate is independent of DIN loads; thereby enabling a consistent interpretation to the 10-yr ARI (viable coral reef) threshold. The central findings of this study are thus strengthened by the intuitive likelihood that reef waters with lowered DIN levels not only assist coral survivorship (during thermal stress events) but also promote recovery (through successful settlement, recruitment and growth) following thermal stress events.
For the rivers of the central GBR, the degree of DIN enrichment is directly related to the percentage of fertiliser-additive land use (typically sugarcane and banana plantations) in the upstream catchment area (see e.g., Mitchell et al. 2009; Bainbridge et al. 2009). For the most enriched river systems (e.g., Tully-Murray catchment near Cardwell) modelling and monitoring show that ~80–85% of the DIN exported to the river mouths is derived from fertiliser loss (Armour et al. 2009; Bainbridge et al. 2009). Thus, a focus on reducing these fertiliser losses is likely to provide the greatest scope for large reductions in end-of-river DIN discharge in the future. This contrasts with the case for suspended sediments. Suspended sediment delivery into the GBR lagoon is dominated by soil erosion processes exacerbated by intensive rangeland beef cattle grazing (Brodie et al. 2003). Low rates of suspended sediment delivery are thus best achieved through reduction in grazing area (especially in areas prone to gullying) in combination with maintenance of high pasture cover (which may involve reduced stocking rates) (Brodie et al. 2003).
Landscape nutrient-budget models show that nitrogen-efficient management practices such as no over–application of fertilisers, reduced tillage, split fertiliser application, and removing production from the least productive soil types can eliminate over 80% of agricultural DIN exports (Roebeling et al. 2009). Linked economic analysis concludes that optimal adoption of these best management practices could reduce DIN delivery by up to ~50% with no additional cost, or even benefit, to the local agricultural industry (Roebeling et al. 2009). In contrast, the reductions in production area and fertiliser application rates needed to attain the more stringent DIN reduction targets (~80%) are only possible at a significant cost to the local agricultural industry (Roebeling et al. 2009). However, the economic cost imposed by such measures is likely to be more than offset by the long-term economic benefits of safeguarding reef ecosystem goods and services, such as those that support the local reef tourism industry (Thomas et al. 2009). The political economy of setting two important regional sectors at odds with each other makes such decisions extremely problematic.
It is important to emphasize that any land management benefit will be most strongly observed in inner- to mid-shelf reef areas where terrestrial nutrient sources are the dominating influence (Furnas 2003). However, a better understanding of the spatio-temporal dynamics of DIN loading across the entire GBR will help identify those areas most vulnerable to heat stress. Within the central GBR, DIN loading is typically highest at the coastal locations most exposed to terrestrial runoff, lowest at mid-shelf locations, and moderate at offshore locations (e.g., see Sammarco et al. 1999). All things being equal, it is thus predicted that the mid-shelf reefs of the central GBR should display the highest resistance to heat stress. This prediction is currently the focus of new research and has important implications for the design of a marine protected area (MPA) network that aims to spread the risk of future bleaching impacts (sensu Done 2001; Game et al. 2008).
In terms of an integrated management target, the present analysis suggests that reduced DIN river loads (~50%) combined with stabilisation of atmosphere CO2 at ~450 ppm can help ensure the long-term viability of hard coral communities within the terrestrial flood zone of the central GBR (see Figs. 4 and 6). The likelihood of achieving these specific local and global management targets depends on the outcomes of political, environmental, economic and social initiatives spanning local to global scales. Policy actions aiming to halt declining water quality on the GBR are being implemented by Australian state and federal governments, including a target of 50% reductions in ambient DIN loads by 2014 (Anon 2007, 2009). To date however, no global climate policy agreement exists between the major CO2-emitting countries that will ensure atmospheric CO2 concentrations stabilise below 450 ppm (Garnaut 2008), although a 2°C warming target was adopted at the United Nations Framework Convention on Climate Change 15th Council of Parties meeting in Copenhagen in late 2009. Achieving this target would most likely require an overshoot trajectory for greenhouse gases peaking at above 450 ppm CO2-equivalent with subsequent reductions to an undetermined goal (Sheehan et al. 2008). Yet, some researchers still consider the achievement of 450 ppm CO2 technically achievable (e.g., van Vuuren et al. 2007). The next decade is critical; if delay of meaningful action stretches beyond 2020, exceedance of 550 ppm becomes all-but unavoidable (Vaughan et al. 2009). Based on the present study, at 550 ppm vast tracts of GBR reef ecosystem would pass the tipping point for maintenance of viable coral reef (10-yr ARI), unless more aggressive reductions (~80%) in end-of-river DIN loads were achieved. This shows the importance of hedging local management strategies against global uncertainty when both have a critical bearing on management outcomes. Early local action will hedge against global uncertainties, particularly if an overshoot situation is reached, where temperatures peak but ultimately decline.
The complementary benefit of global policies that stabilise atmospheric CO2 below 450 ppm is also found in the reduced severity of future ocean acidification impacts (see e.g., Guinotte et al. 2003). Studies into the sub-lethal impacts on tropical reefs due to ocean acidification are still in their infancy, as are considerations into possible interactions with temperature stress (Kleypas et al. 2006; Reynaud et al. 2003; Anthony et al. 2008). Encouragingly, the negative impact of elevated CO2 concentrations on skeletal growth rates is also lessened by good quality water, particularly reduced DIN (Renegar and Riegl 2005). Whether good water quality in combination with atmospheric CO2 stabilisation ~450 ppm is sufficient to avoid the most serious impacts from ocean acidification is the subject of on-going research and will be reported elsewhere.
5 Conclusions
This study reinforces the proposition that there are no ‘silver bullet’ management solutions for safeguarding shallow water coral reefs against the risks from anthropogenic climate change during the 21st century (sensu Marshall and Schuttenberg 2006). Declines in productivity and reef-building capacity are already likely for many coastal reef areas on the central GBR; climate change left unmitigated could see them terminally degraded before 2050. Yet this outcome can still be avoided through a realistic combination of local and global management actions. This study has identified a minimum threshold for local reductions in river DIN loads of ~50% and global stabilisation of atmospheric CO2 below 450 ppm. Although the resulting near-term reefscapes may still fall short of the iconic panoramas enjoyed today, the key ecological function of symbiotic corals as reef-builders can potentially be maintained. From this most basic functional state it must be hoped that natural selective processes, for example endosymbiont changes towards more stress resistance zooxanthellae types (Baker et al. 2004; Berkelmans and van Oppen 2006) and/or community composition shifts towards more tolerant host species (Done 1999; Loya et al. 2001), can lead to a maintenance of provision of essential ecosystem goods and services. Swift action will ensure that the goal of maintaining viable coral reef communities and their attendant benefits remains possible, leaving little room for delay. To date, global efforts to reduce CO2 emissions have been limited in magnitude and weakly implemented; sustained cuts are needed to stabilise atmospheric CO2 concentration below 450 ppm. For the GBR, comprehensive government policies partnered with on-ground implementation programs have been introduced to halt and reverse the decline of water quality entering coastal waters. This study highlights the significant potential of such actions for enhancing reef survivability during thermal stress in addition to aiding its recovery after stress. This capacity is an important hedging mechanism for managing global risks to coral reef systems.
References
Anon (2007) Labour’s reef rescue plan. Policy Document, Australian Labour Government, Canberra, p. 13. http://www.alp.org.au/download/071028___labors-reef-rescue-plan.pdf
Anon (2009) Reef water quality protection plan 2009 for the Great Barrier Reef world heritage area and adjacent catchment. Policy Document, Queensland State Labour Government, Brisbane, p. 32. http://www.reefplan.qld.gov.au/library/pdf/reef-plan-2009.pdf
Anthony KRN, Kline DI, Diaz-Pulido G, Dove S, Hoegh-Guldberg O (2008) Ocean acidification causes bleaching and productivity loss in coral reef builders. Proc Natl Acad Sci U S A 105:17442–17446
Armour JD, Hateley LR, Pitt GL (2009) Catchment modelling of sediment, nitrogen and phosphorus nutrient loads with SedNet/ANNEX in the Tully-Murray basin. Mar Freshw Res 60:1091–1096
Bainbridge ZT, Brodie JE, Faithfull JW, Sydes DA, Lewis SE (2009) Identifying the land-based sources of suspended sediments, nutrients and pesticides discharged to the Great Barrier Reef from the Tully-Murray Basin, Queensland. Aust Mar Freshwat Res 60:1081–1090
Baker AC, Starger CJ, McClanahan TR, Glynn PW (2004) Corals’ adaptive response to climate change. Nature 430:741
Baohua Z, Guangce W, Bo H, Tseng CK (2004) Effects of temperature, hypoxia, ammonia and nitrate on the bleaching among three coral species. Chin Sci Bull 49:1923–1928
Berkelmans R (2009) Bleaching and mortality thresholds: how much is too much? In: van Oppen MJH, Lough JM (eds) Coral bleaching: patterns, processes, causes and consequences. Ecological Studies, Springer, pp 103–119
Berkelmans R, van Oppen MJH (2006) The role of zooxanthellae in the thermal tolerance of corals: a ‘nugget of hope’ for coral reefs in an era of climate change. Proc R Soc London B273:2305–2312
Brodie JE, McKergow LA, Prosser IP, Furnas MJ, Hughes AO, Hunter H (2003) Sources of sediment and nutrient exports to the Great Barrier Reef World Heritage Area. ACTFR Report No. 03/11, Australian Centre for Tropical Freshwater Research, James Cook University, Townsville. http://www.actfr.jcu.edu.au/Publications/index.html
Carilli JE, Norris RD, Black B, Walsh SM, McField M (2009a) Century-scale records of coral growth rates indicate that local stressors reduce coral thermal tolerance threshold. Global Change Biol. doi:10.1111/j.1365-2486.2009.02043.x
Carilli JE, Norris RD, Black BA, Walsh SM, McField M (2009b) Local stressors reduce coral resilience to bleaching. PloS One 4(7):e6324. doi:10.1371/journal.pone.0006324
Coffroth MA, Poland DM, Petrou EL, Brazeau DA, Holmberg JC (2010) Environmental symbiont acquisition may not be the solution to warming seas for reef-building corals. Plos One 5:e13258
Coles SL, Brown BE (2003) Coral bleaching—capacity for acclimatization and adaptation. Adv Mar Biol 46:183–223
Connell JH (1997) Disturbance and recovery of coral assemblages. Coral Reefs 16:101–113
CSIRO, BoM (ed) (2007) Climate change in Australia. CSIRO, Melbourne
Davies PJ (1983) Reef growth. In: Barnes DJ (ed) Perspectives on coral reefs. Australian Institute of Marine Science, Townsville, pp 69–95
De’ath G, Fabricius K (2010) Water quality as a regional driver of coral biodiversity and macroalgae on the Great Barrier Reef. Ecol Appl 20:840–850
Devantier LM, De’ath G, Done TJ, Turak E (1998) Ecological assessment of a complex natural system: a case study from the Great Barrier Reef. Ecol Appl 8:480–496
Devlin M, Waterhouse J, Taylor J, Brodie J (2001) Flood plumes in the Great Barrier Reef: spatial and temporal patterns in composition and distribution. Research publication 68, Great Barrier Reef Marine Park Authority, Townsville, Australia
Done T (1982) Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1:75–90
Done TJ (1999) Coral community adaptability to environmental changes at scales of regions, reefs and reef zones. Am Zool 39:66–79
Done T (2001) Scientific principles for establishing MPAs to alleviate coral bleaching and promote recovery. In: Salm R, Coles SL (eds) Coral bleaching and marine protected areas. Proceedings of the workshop on mitigating coral bleaching impact through MPA design. Bishop Museum, Honolulu, 29–31 May, 2001. Asia Pacific Coastal Marine Program Report No. 0102, The Nature Conservancy, Honolulu, Hawaii, pp. 53–59
Done TJ, Ogden JC, Wiebe WJ, Rosen BR (1996) Biodiversity and ecosystem function of coral reefs. In: Mooney HA, Cushman JH, Medina E, Sala OE, Schultze ED (eds) Functional roles of biodiversity: a global perspective. John Wiley, Chichester, pp 393–429
Done TJ, Turak E, Wakeford M, DeVantier L, McDonald A, Fisk D (2007) Decadal changes in turbid-water coral communities at Pandora Reef: loss of resilience or too soon to tell? Coral Reefs 26:789–805
Fabricius KE (2005) Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Mar Poll Bull 50:125–146
Faxneld S, Jörgensen TL, Nguyen ND, Nyström M, Tedengren M (2011) Differences in physiological response to increased seawater temperature in nearshore and offshore corals in northern Vietnam. Mar Environ Res 71:225–233
Fitt WK, Brown BE, Warner ME, Dunne RP (2001) Coral bleaching: interpretation of thermal tolerance limits and thermal thresholds in tropical corals. Coral Reefs 20:51–65
Furnas MJ (2003) Catchments and corals: terrestrial Runoff to the Great Barrier Reef. Australian Institute of Marine Science and CRC Reef Research Centre, Townsville
Game E, Watts M, Wooldridge SA, Possingham H (2008) Planning for persistence in marine reserves: a question of catastrophic importance. Ecol Appl 18:670–680
Garnaut R (2008) The Garnaut climate change review. Cambridge University Press, Melbourne
Goreau TJ (1992) Bleaching and reef community change in Jamaica: 1951–1991. Am Zool 32:683–696
Guinotte JM, Buddemeier RW, Kleypas JA (2003) Future coral reef habitat marginality: temporal and spatial effects of climate change in the Pacific basin. Coral Reefs 22:551–558
Haas A, Al-Zibdah M, Wild C (2009) Effect of inorganic and organic nutrient addition on coral-algae assemblages from the Northern Red Sea. J Exp Mar Biol Ecol 380:99–105
Johnson JE, Marshall PA (2007) Climate change and the Great Barrier Reef: a vulnerability assessment. Great Barrier Reef Marine Park Authority and Australian Greenhouse Office, Australia
King B, McAllister F, Done T (2002) Modelling the impact of the Burdekin, Herbert, Tully and Johnstone River plumes on the central Great Barrier Reef. CRC Reef Research Centre Technical Report No. 44, CRC Research Centre, Townsville
Kleypas JA, Feely RA, Fabry VJ et al (2006) Impacts of ocean acidification on coral reefs and other marine calcifiers: a guide for future research. Report of a Workshop held 18–20 April 2005, St. Petersburg Florida. Sponsored by NSF, NOAA, and the US Geological Survey, p. 88
Kuntz N, Kline DI, Sandin SA, Rohwer F (2005) Pathologies and mortality rates caused by organic carbon and nutrient stressors in three Caribbean coral species. Mar Ecol Prog Ser 294:173–180
LaJeunesse TC, Smith R, Walther M et al (2010) Host-symbiont recombination versus natural selection in the response of coral-dinoflagellate symbioses to environmental disturbance. Proc R Soc London B277:2925–2934
Loya Y, Sakai K, Yamazato K, Nakano Y, Sambali H, van Woesik R (2001) Coral bleaching: the winners and the losers. Ecol Lett 4:122–131
Marshall PA, Baird AH (2000) Bleaching of corals on the Great Barrier Reef: differential susceptibilities among taxa. Coral Reefs 19:155–163
Marshall PA, Schuttenberg HZ (2006) Adapting coral reef management in the face of climate change. In: Phinney JT, Strong A, Skrving W, Kleypas J, Hoegh-Guldberg O (eds) Coral reefs and climate change: science and management. American Geophysical Union Coastal and Estuarine Series, volume 61, pp. 244
Maughan M, Brodie J (2009) Reef exposure to river-borne contaminants: a spatial model. Mar Freshw Res 60:1132–1140
McCook LJ, Folke C, Hughes TP, Nyström M, Obura D, Salm R (2007) Ecological resilience, climate change and the Great Barrier Reef. In: Johnson JE, Marshall PA (eds) Climate change and the Great Barrier Reef: a vulnerability assessment. Great Barrier Reef Marine Park Authority, Townsville, pp 75–96
Mitchell TD (2003) Pattern scaling: an examination of the accuracy of the technique for describing future climates. Clim Change 60:217–242
Mitchell A, Reghenzani J, Faithful J, Furnas M, Brodie J (2009) Relationships between land use and nutrient concentrations in streams draining a ‘wet-tropics’ catchment in northern Australia. Mar Freshw Res 60:1097–1108
Renegar DA, Riegl BM (2005) Effect of nutrient enrichment and elevated CO2 partial pressure on growth rate of Atlantic scleractinian coral Acropora cervicornis. Mar Ecol Prog Ser 293:69–76
Reynaud S, Leclercq N, Romaine-Lioud S, Ferrier-Pagès C, Jaubert J, Gattuso JP (2003) Interacting effects of CO2 partial pressure and temperature on photosynthesis and calcification in a scleractinian coral. Global Change Biol 9:1660–1668
Roebeling PC, van Grieken ME, Webster AJ, Biggs J, Thorburn P (2009) Cost-effective water quality improvement in linked terrestrial and marine ecosystems: a spatial environmental-economic modelling approach. Mar Freshw Res 60:1150–1158
Sabine CL, Feely RA, Gruber N et al (2004) The oceanic sink for anthropogenic CO2. Science 305:367–371
Sammarco PW, Risk MJ, Schwarcz HP, Heikoop JM (1999) Cross-continental shelf trends in coral 15N on the Great Barrier Reef: further consideration of the reef nutrient paradox. Mar Ecol Prog Ser 180:131–138
Schlöder C, D’Croz L (2004) Responses of massive and branching coral species to the combined effects of water temperature and nitrate enrichment. J Exp Mar Biol Ecol 313:255–268
Sheehan P, Jones RN, Jolley A, Preston BL, Clarke M, Durack PJ, Islam SMN, Whetton PH (2008) Climate change and the new world economy: implications for the nature and timing of policy responses. Global Environ Change—Hum Pol Dimens 18:380–396. doi:10.1016/j.gloenvcha.2008.04.008
Skirving W, Mahoney M, Steinberg C (2002) Sea surface temperature atlas of the Great Barrier Reef. 1990–2002. Available on CD-ROM from the Australian Institute of Marine Science, Townsville, Australia
Thomas CR, Gordon IJ, Wooldridge SA, van Grieken M, Marshall P (2009) The development of an integrated systems model for balancing coral reef health, land management and tourism risks on the Great Barrier Reef. Proceeding of the 18th World IMACS/MODSIM Congress, Cairns, Australia 13–17 July 2009, pp. 4346–4352. (http://www.mssanz.org.au/modsim09/J3/thomas_cr.pdf)
Thompson AA, Dolman AM (2010) Coral bleaching: one disturbance too many for near-shore reefs of the Great Barrier Reef. Coral Reefs 29:637–648
van Vuuren DP, den Elzen MG, Lucas PL et al (2007) Stabilizing greenhouse gas concentrations at low levels: and assessment of reduction strategies and costs. Clim Change 81:119–159
Vaughan NE, Lenton TM, Shepherd JG (2009) Climate change mitigation: trade-offs between delay and strength of action required. Clim Change 96:29–43
Veron JEN, Hoegh-Guldberg O, Lenton TM et al (2009) The coral reef crisis: the critical importance of <350 ppm CO2. Mar Poll Bull 58:1428–1436
Wagner DE, Kramer P, van Woesik R (2010) Species composition, habitat, and water quality influence coral bleaching in south-eastern Florida. Mar Ecol Prog Ser 408:65–78
Wakeford M, Done TJ, Johnson CR (2008) Decadal trends in a coral community and evidence of changed disturbance regime. Coral Reefs 27:1–13
Whetton P, Macadam I, Bathols J, O’Grady J (2007) Assessment of the use of current climate patterns to evaluate regional enhanced greenhouse response patterns of climate models. Geophys Res Lett 35:L14701. doi:10.1029/2007GL030025
Wooldridge SA (2008) Modelling the improved resilience of inshore coral reefs to climate change due to terrestrial water quality improvements: A case study from the Tully-Murray River catchment, North Queensland. Report to the Marine and Tropical Sciences Research Facility. Reef and Rainforest Research Centre Limited, Cairns, p 19
Wooldridge SA (2009a) Water quality and coral bleaching thresholds: formalising the linkage for the inshore reefs of the Great Barrier Reef, Australia. Mar Poll Bull 58:745–751
Wooldridge SA (2009b) A new conceptual model for the warm-water breakdown of the coral-algae endosymbiosis. Mar Freshw Res 60:483–496
Wooldridge SA, Done TJ (2009) Improved water quality can ameliorate effects of climate change on corals. Ecol Appl 19:1492–1499
Wooldridge SA, Done TJ, Berkelmans R, Jones R, Marshall PA (2005) Precursors for resilience in coral communities in a warming climate: a belief network approach. Mar Ecol Prog Ser 295:157–169
Wooldridge SA, Brodie J, Furnas M (2006) Exposure of inner-shelf reefs to nutrient enriched runoff entering the Great Barrier Reef Lagoon: post-European changes and the design of water quality targets. Mar Poll Bull 52:1467–1479
Acknowledgments
This study was supported by the Australian Government’s Marine and Tropical Science Research Facility (MTSRF). We acknowledge the helpful suggestions of two anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Table A1
Global circulation models used to predict future sea surface temperature patterns (DOCX 15 kb)
Supplementary Fig. A1
Conceptual representation of the z-score approach for modelling the AEP of LD50 mortality events on the GBR. With the z-score approach, the likelihood of exceeding the spatially explicit LD50 threshold can be conceptualised as the area under the curve in excess of the threshold, Z (DOCX 49 kb)
Rights and permissions
About this article
Cite this article
Wooldridge, S.A., Done, T.J., Thomas, C.R. et al. Safeguarding coastal coral communities on the central Great Barrier Reef (Australia) against climate change: realizable local and global actions. Climatic Change 112, 945–961 (2012). https://doi.org/10.1007/s10584-011-0229-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-011-0229-z