
Abstract
Aβ oligomers cause a collection of molecular events associated with memory loss in Alzheimer’s disease, centering on disrupting the maintenance of synapse structure and function. In this brief review of the synaptotoxic effects of Aβ oligomers, we focus on the neuronal properties governing oligomer targeting and toxicity—especially with respect to binding sites and mechanisms of binding. We also discuss ways in which mechanistic insights from other diseases offer clues in the pursuit of the molecular basis of Alzheimer’s disease.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Marshall Nirenberg’s career was emblematic of his drive to uncover the fundamental nature of information storage in biology. After his seminal studies on the genetic code, he focused on the storage of information in the brain—a process then and now theorized to result from the formation of stable synaptic connections. Thus, he sought to learn how and why synapses form as a result of our interaction with the world around us. At the opposite extreme, Alzheimer’s disease (AD) is outwardly manifest as a debilitating inability to form new memories—and deficits in synapse formation and maintenance are now commonly thought to be its root cause.
The Initial Implication of Oligomeric Aβ in AD
The identification of the amyloid β peptide as the major component of senile plaques in AD brains (Masters and Beyreuther 1987; Glenner and Murphy 1989) spurred efforts to identify the mechanisms by which aggregates of Aβ might be capable of interfering with brain function. The amyloid cascade hypothesis predicted that reducing the buildup of amyloid plaques should reduce the memory impairment observed in AD (Hardy and Higgins 1992). However, experiments in animal models failed to attribute memory impairment to amyloid pathology—finding instead that plaque load does not dictate the severity of AD (Haass and Selkoe 2007). Soluble oligomeric forms of Aβ were first described in the early 1990s, but little thought was given to their being the primary toxic species in AD (Roher et al. 1991; Frackowiak et al. 1994; Kuo et al. 1996; Podlisny et al. 1995; Roher et al. 1993; Vigo-Pelfrey et al. 1993). Arising from the hypothesis that reducing amyloid fibrils would reduce Aβ toxicity, Oda et al. incubated Aβ peptides with clusterin, which prevented fibril formation but increased oxidative stress in PC12 cells treated with these preparations (Oda et al. 1994). In the wake of this finding, Lambert et al. (1998) showed that fibril-free soluble Aβ oligomers are neurotoxic and lead to rapid inhibition of long-term potentiation (LTP; an electrophysiological correlate of learning and memory) and eventually cell death. The name amyloid β-derived diffusible ligand (ADDL), coined in that work, was designed to emphasize the ability of these oligomers to act as specific toxins and was meant to differentiate those Aβ oligomers with dementing activity. That is, it was recognized that not every oligomer should be considered toxic. To this day, an exact assembly state and conformation of the toxin is still intensely sought after—if a single state exists—and the transient nature and heterogeneity of Aβ oligomers impedes efforts to define the structural basis of their toxic activity.
Soluble Aβ oligomers have been described that adopt a variety of aggregation states as small as dimers and as large as protofibrillar structures comprising dozens Aβ monomers (for reviews, see Glabe 2008; Rahimi et al. 2008). While toxic agency has been ascribed to oligomer preparations enriched in high and low molecular weight oligomers, suggesting the possibility of dementing activity across a range of oligomer species, dodecameric and dimeric Aβ species have been a focal points for studies of Aβ oligomer toxicity. Oligomers of Aβ containing roughly 12 monomers were identified in human brain extracts and found to bind cultured neurons (Gong et al. 2003) in a manner similar to synthetic ADDLs, in which high molecular weight species preferentially target neurons (Lacor et al. 2004). The implication of these brain-derived dodecamers as pathophysiologically relevant Aβ oligomers is supported by the isolation of a similar 56 kDa species from APP-overexpressing AD transgenic mice that was capable of disrupting memory upon injection into young wild type rats (Lesne et al. 2006). Dodecamers have also been described by additional groups using synthetic Aβ oligomers (Barghorn et al. 2005; Bernstein et al. 2009). At the lower end of the oligomer spectrum, dimeric Aβ was recently purified from AD brains and also found to inhibit LTP in hippocampal slices (Shankar et al. 2008) and induce Tau hyperphosphorylation and other degenerative effects in cultured neurons (Jin et al. 2011). This finding follows the extensive use of cell culture-derived oligomers by Selkoe and others that are reported to be primarily dimers and trimers of Aβ (Selkoe 2008; Walsh et al. 2002, 2000).
Synaptic Effects of Aβ Oligomers
Aβ oligomers exert their toxicity through binding at synapses. As might be expected for a molecule that disrupts LTP, synthetic oligomers as well as AD brain- or CSF-derived soluble oligomers consistently show abundant binding to the dendritic arbors of select hippocampal neurons in culture (Gong et al. 2003; Lacor et al. 2004) and this dendritic binding is evident in the form of punctate labeling that co-localizes with PSD-95 as visualized by high-resolution confocal fluorescence microscopy (Lacor et al. 2004). Oligomers also co-localize with PSD-95 in brain sections of AD transgenic mice (Koffie et al. 2009). Oligomer binding sites are enriched with postsynaptic proteins including calcium/calmodulin-dependent kinase II, Arc, spinophilin, drebrin and n-methyl-d-aspartate receptor (NMDAR), while the presynaptic marker, synaptophysin, is opposite these puncta (Lacor et al. 2004; Lacor et al. 2007). Excitatory synapses appear to be preferentially targeted, as oligomers co-localize with PSD-95 and Homer1b/c, scaffolding proteins that interact with glutamate receptors and Shank family proteins at excitatory synapses (Tu et al. 1999), while no colocalization is evident in the case of gephyrin, a scaffolding protein at inhibitory synapses (Renner et al. 2010). Immunoisolation of ADDL-binding complexes in synaptosomes results in the identification of several prominent glutamate receptors (NR1, NR2, GluR1, and mGluR5), EphB2, and neuroligin, but neither nicotinic acetylcholine receptors nor glycine receptors were present—further demonstrating excitatory synapse targeting (Lacor et al. 2007; Renner et al. 2010).
The effect of the synaptic association of Aβ oligomers is synapse loss, which is accompanied by a collection of effects on synapse size, shape, and composition (reviewed in Lacor 2007). Exposure to oligomers leads to a reduction in the stubby and mushroom spines found in healthy neurons and the transient formation of aberrant spines with slender filopodial or large, branched morphologies reminiscent of structures formed in mental retardation (Lacor et al. 2004; Lacor et al. 2007; Shrestha et al. 2006). In the early timescale prior to synapse loss, oligomers induce changes in the makeup of synaptic membranes, evident as a loss of surface glutamate receptors—both NMDA (Lacor et al. 2007; Snyder et al. 2005) and α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate (AMPA; Hsieh et al. 2006) subtypes—as well as EphB2 (Lacor et al. 2007; Cisse et al. 2011) and insulin receptors (De Felice et al. 2009; Zhao et al. 2009). The trafficking-mediated surface loss of synaptic receptors prior to structural changes and spine loss is consistent with the postulated loss of synaptic plasticity without cell death in early-stage AD (Klein et al. 2007; Lambert et al. 1998).
Findings from post-mortem analyses of AD tissues have also shown an oligomer association with AD. A loss of specific synaptic proteins has been observed that is correlated with AD severity and regionally specific neurodegeneration (Proctor et al. 2010) and post-mortem staining of early stage AD brains using oligomer-selective antibodies (Lacor et al. 2004) show diffuse immunostaining reminiscent of the synaptic-type deposits observed in prion-associated diseases (Kovacs et al. 2002). Other experiments using oligomer-specific antibodies have identified a 70-fold increase in the concentration of Aβ oligomers in AD brains (Gong et al. 2003), an increase that is also detectable in the CSF of individuals with AD, although to a lesser degree (Georganopoulou et al. 2005). Oligomer staining using conformationally specific antibodies was also shown to be spatially distinct from fibrillar deposits in human brain slices (Kayed et al. 2003), implying that these species can form independently of amyloid fibrils.
Effect of Aβ Oligomers on Memory
The early discovery of LTP impairment by synthetic soluble Aβ oligomers (Lambert et al. 1998) has been supported by experiments using oligomers released by hAPP-transfected cells (Walsh et al. 2002) and with soluble extracts from the brains of AD subjects and transgenic mice (Lesne et al. 2006; Shankar et al. 2008). Oligomers also impede the reversal of long-term depression (LTD) (Hsieh et al. 2006; Wang et al. 2002), indicating a net shift of synaptic activity in favor of inhibition suggesting instability in synaptic composition and morphology (Klein et al. 2001; Lacor et al. 2004). LTP impairment in AD transgenic mice correlates with accumulation of soluble oligomers (Chang et al. 2003) but precedes Aβ deposition into plaques (Larson et al. 1999; Oddo et al. 2006). The temporal correlation of LTP with oligomer accumulation is borne out by behavioral testing for memory in animal models (Cleary et al. 2005; Lesne et al. 2006).
Various animal models have proven useful in defining the biological presence and dementing activity of Aβ oligomers. Early work by Mucke demonstrated that Aβ overexpression in transgenic mice causes AD-like synapse loss without the production of amyloid plaques (Mucke et al. 2000). Oligomeric Aβ was positively identified in transgenic animals in 2003 (Chang et al. 2003), and oligomers corresponding roughly to 12-mers were isolated from AD transgenic mice and found to impair memory upon injection into wild type rats (Lesne et al. 2006). The behavioral influence of oligomeric Aβ rather than plaques is further supported by experiments using transgenic mice in which soluble oligomers accumulate without plaque formation at any age tested, up to 24 months (Tomiyama et al. 2010). Similar mice lacking fibrils while at the same time producing oligomeric Aβ species have also been described (Gandy et al. 2010). These different lines of transgenic animals were made by altering the same amino acid residue in the amyloid precursor protein. In the former study, glutamic acid 693—corresponding to position 22 in the Aβ peptide—was deleted based on a similar deletion found in a Japanese family that develop a form of AD with decreased levels of Aβ, and studies of this mutated peptide reveal a propensity to form oligomers but not fibrils (Tomiyama et al. 2008). In the latter mice, the same residue was mutated to a glutamine (E693Q). It is interesting to note that despite the apparent lack of amyloid plaques in E693Q mice, the mutation in humans is responsible for a form of hereditary cerebral hemorrhage with amyloidosis (Levy et al. 1990), which is accompanied by Aβ deposition.
How Do Aβ Oligomers Target Neurons?
When considering the basis of oligomer attachment to neurons, three competing hypotheses dominate the landscape. One hypothesis is that oligomeric Aβ interacts directly with membranes—perhaps to form toxic pores. Findings that Aβ peptides and oligomers can insert into model membranes of varying compositions support this hypothesis (Lashuel and Lansbury 2006), which has been proposed to constitute a general gain-of-function underlying the toxicity of multiple proteins involved in neurodegenerative disease (Kayed et al. 2003). Studies using both model membranes and intact cells have identified an effect of negatively charged phospholipids in mediating the interaction of Aβ peptides with neuronal membranes (Alarcon et al. 2006; Hertel et al. 1997; McLaurin and Chakrabartty 1997; Wong et al. 2009), perhaps even catalyzing oligomer formation (Wong et al. 2009).
An alternative hypothesis is that toxic Aβ oligomers act within the cell (Takahashi et al. 2004; Walsh et al. 2000) to exert a toxic effect on synapses. Intracellular Aβ, though not necessarily oligomers, is observed in AD brains (Gouras et al. 2000) and there is evidence of intracellular Aβ generation (reviewed in LaFerla et al. 2007). Recent AD transgenic rat models feature intracellular oligomers as detected by an oligomer-specific monoclonal antibody, suggesting that Aβ oligomers are present inside neurons (Leon et al. 2010; Tomiyama et al. 2010). Whether oligomers form extracellularly, intracellularly, or both is not agreed upon in the literature. Oligomeric Aβ may form within neurons prior to export (Walsh et al. 2000), but the demonstration that Aβ monomer concentrations in the interstitial fluid of the brain undergo a circadian cycle in living mice highlights the likelihood that oligomer formation is also occurring extracellularly (Kang et al. 2009). The toxicity of intracellular Aβ may also depend on the aggregation state, as an inverse correlation was reported between intracellular Aβ monomers and nucleic acid oxidation—hypothesized to be a protective mechanism against oxidative stress (Nunomura et al. 2010).
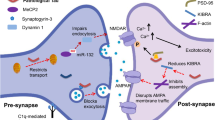
Finally, the loss of synaptic activity associated with Aβ oligomers may be triggered by oligomer binding to specific sites on the neuronal surface—an attractive model because it immediately offers a basis for therapeutic intervention against toxic oligomers. All regions of the AD brain are not equally affected (Braak and Braak 1991), and this holds true at the cellular level, where the cultured hippocampal neurons are highly targeted by Aβ oligomers, while cerebellar neurons are not affected (Gong et al. 2003; Klein et al. 2001). Further evidence of specific susceptibility of different neuronal populations is the triggering of mitochondrial dysfunction in cortical, but not cerebellar preparations by Aβ oligomers (Eckert et al. 2008). Dramatic cell-to-cell differences in oligomer binding exist in hippocampal cultures, illustrating that even in a targeted region there is some determining factor that enables oligomers to bind a given cell (Lacor et al. 2007). Similar to these cell-to-cell differences, Aβ oligomers seem to be specific for a subset of synapses within a single cell. While 90% of oligomers co-localize with synaptic markers, only half of the excitatory synapses bind oligomers (Lacor et al. 2004). This fractional synaptic targeting increases upon neuronal activation (Deshpande et al. 2009), which may reflect changes in synaptic receptor regulation. The trypsin sensitivity of oligomer binding and observations that these molecules preferentially affect mature neurons—developing the ability to bind oligomers around 14 days in vitro—suggest the presence of a developmentally regulated proteinaceous receptor (Lacor et al. 2007; Lambert et al. 1998; Shughrue et al. 2010).
Proposed Binding Sites for Aβ Oligomers
As yet, no single protein seems to recapitulate all of the necessary characteristics of a true oligomer receptor. Even non-protein binding sites have been proposed, including GM1 gangliosides and the lipid rafts they inhabit, which have been implicated in multiple aspects of AD pathophysiology including APP processing (Fonseca et al. 2010) and Aβ oligomer binding (Gong et al. 2003; Zampagni et al. 2010). In fact, GM1 gangliosides were reported to directly mediate the binding of toxic calcitonin oligomers (Malchiodi-Albedi et al. 2010). Ganglioside removal by neuraminidase treatment caused the complete elimination of calcium influx in the presence of calcitonin oligomers, but lipid raft disruption by ganglioside removal may also disorganize other receptors prominent in these membrane domains.
Many proteinaceous receptors for Aβ oligomers have been reported in the past several years. Among them are the P75 neurotrophin receptors, the antibody-based blockade of which prevents cell death after exposure to relatively high doses of Aβ oligomers (Knowles et al. 2009); the receptor for advanced glycation endproducts, or “RAGE” (Sturchler et al. 2008); the frizzled receptor (Magdesian et al. 2008), and nicotinic acetylcholine receptors (Magdesian et al. 2005), which were shown to bind monomeric Aβ peptides.
Perhaps the most notable and controversial recent example of a receptor for Aβ oligomers is the cellular prion protein, PrPc, identified in a gene expression screen in a non-neuronal cell line for proteins allowing ADDL binding, which the cells normally lack (Lauren et al. 2009). The prospect that PrPc is the binding site for Aβ oligomers was quickly and alternately refuted (Calella et al. 2010; Kessels et al. 2010; Balducci et al. 2010) and supported (Gimbel et al. 2010; Chen et al. 2010) by multiple follow-up studies. Inconsistent or system-specific findings of PrPc-independent oligomer toxicity and memory deficits may require a mechanism by which PrPc participates in the clustering of oligomers with other receptors, allowing a stronger response to low oligomer concentrations (Laurén et al. 2010).
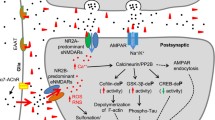
As mentioned above, loss of EphB2 receptors accompanies Aβ oligomer binding to neurons in culture (Lacor et al. 2007). Recent investigations of the mechanism of oligomer-mediated receptor internalization point to a direct interaction of dimers and trimers of Aβ with EphB2, leading to its internalization and proteasomal degradation and a surface reduction in NR1—a subunit of NMDA receptors (Cisse et al. 2011). Treatment of cultured neurons with antibodies against NR1 leads to only a 50% reduction in oligomer binding while eliminating oligomer-induced generation of reactive oxygen species, and oligomer-induced effects can also be reduced using the NMDAR antagonist, memantine (De Felice et al. 2007; Lacor et al. 2007). The AMPA receptor subunit GluR2 may also contribute to oligomer binding but, as with other receptor candidates, reduction in AMPA receptor surface expression incompletely reduces synaptic oligomer binding (Zhao et al. 2010).
Finally, experiments using a mouse knockout of mGluR5 implicate this receptor in oligomer-induced synaptic pathology (Renner et al. 2010). While the approaches used in this study did not permit a direct analysis of whether mGluR5 interacts with oligomeric Aβ, the work demonstrates an essential role of mGluR5 in oligomer-induced synaptic reorganization as described in the next section.
Receptor Clustering: A Mechanism for Oligomer Toxicity in AD
How can so many receptors play a role in oligomer binding and synaptotoxicity? One hypothesis is that because Aβ oligomers likely comprise a distribution of states, they bind to multiple receptors. Another is that a single oligomer can alternately bind to different, low-affinity receptors. If either of these models were true, then the sum of the contributions of each receptor should equal the observed oligomer binding. While it appears that when each of the proposed receptors is ablated through some means, only a fractional decrease in oligomer binding is typically observed (De Felice et al. 2009; Lauren et al. 2009; Renner et al. 2010; Zhao et al. 2010), a combination of PrPc, mGluR5, and NR1 antibodies applied simultaneously to cultured hippocampal neurons does not augment the fractional reduction in oligomer binding achieved using any of the individual receptor antibodies (Renner et al. 2010). This non-additivity suggests that a more complex mechanism is controlling the synaptic targeting of Aβ oligomers.
A new mechanism (Fig. 1) that may explain the implication of multiple receptors in oligomer toxicity is based on findings that oligomers undergo a progressive shift from an extrasynaptic, freely diffusive state toward the formation of static synaptic clusters. This clustering was monitored through the real-time single-particle tracking of quantum dot-labeled oligomers on the surface of live neurons (Renner et al. 2010). The reduced oligomer diffusion observed in these experiments mirrors the reduced diffusion accompanying transmembrane protein recruitment to specific sites (Douglass and Vale 2005; Geng et al. 2009), suggesting that oligomer clustering and immobilization may depend on specific receptors. Specifically, tracking mGluR5 diffusion following oligomer treatment reveals a diffusional restriction at synapses—reminiscent of oligomer tracking data (Renner et al. 2010). Even in the absence of Aβ oligomers, antibodies to an extracellular mGluR5 epitope artificially reduce mGluR5 diffusion and induce its clustering at synaptic sites, suggesting that oligomeric Aβ acts as an extracellular scaffold to bring together clusters of mGluR5—and likely other proteins—at the synapse. Interestingly, oligomer treatment has no effect on the surface diffusion of AMPA-type glutamate and GABAA receptors, suggesting a specificity of Aβ oligomers for mGluR5-associated binding sites (Renner et al. 2010). Though there is currently no evidence that the receptor responsible for oligomer-induced clustering is mGluR5, its signaling activity following artificial clustering could be responsible for NR1 surface withdrawal and the rise in intracellular calcium levels in response to Aβ oligomers.

Formation and synaptic targeting of Aβ oligomers. A neuron is represented with blue neurites and red dendritic spines (background image). Following APP cleavage at the neural membrane by β- and γ-secretase, the 42-residue Aβ peptide (red) can be deposited into senile plaques or can alternatively form a collection of oligomeric species (left inset). Some component of this collection of oligomers interacts with neurons to form synaptic clusters which are associated with a variety of synaptic pathologies including the internalization of receptors involved in synapse function and maintenance and the initiation of toxic signaling cascades (right inset)
The shift in mGluR5 toward synapses in the presence of oligomers is significant in that mGluR5 is involved in the mechanisms of synaptic plasticity underlying learning and memory (Simonyi et al. 2005) and contributes to oligomer-induced synaptotoxicity (Hsieh et al. 2006; Li et al. 2009; Wang et al. 2004). The involvement of mGluR5 also suggests a mechanism for how oligomers affect LTP/LTD and calcium homeostasis, especially given that mGluR5 antagonists prevent these cellular responses to oligomer treatment (Renner et al. 2010; Shankar et al. 2008; Townsend et al. 2007; Wang et al. 2004).
Mechanistic Clues from Other Diseases
As details of the molecular basis of AD emerge, it is becoming clear that AD shares aspects of other brain diseases. We wish to highlight two examples of diseases that appear to share some of the phenomenological and molecular details of AD.
mGluR5 as a connection to Fragile X syndrome
While AD-related deficits in learning and memory have a strong connection to aging, a possible parallel can be found in Fragile X syndrome—an autism spectrum disorder and the leading hereditary cause of mental retardation in children. As with AD, Fragile X appears to be the result of synaptic deficiencies leading to decreased mental capabilities. Many of the synaptic effects mirror those discussed above in AD, including aberrant spine morphology (Irwin et al. 2001), AMPA and NMDA receptor dysfunction (Yun and Trommer 2010; Zalfa et al. 2007), Arc overexpression and elevated LTD (Park et al. 2008), and the activation of the protein phosphatase PP2A and subsequent inactivation of MAPK1/2 (Kim et al. 2008)—also impaired in oligomer-exposed neurons (Wang et al. 2004).
Mechanistically, the most relevant commonality between Fragile X and oligomer synaptotoxicity is the involvement of mGluR5 receptors. In normal neurons, mGluR5 mobilization of calcium leads to dephosphorylation of fragile X mental retardation protein (FMRP), a protein that regulates the trafficking of specific mRNA transcripts to synaptic spines and their subsequent translation. FMRP phosphorylation downstream of mGluR5 relieves the translational repression of FMRP on its mRNA cargo (Bear et al. 2004). Excessive mGluR5 signaling activity is prevented by FMRP itself, through a negative feedback loop (Bear et al. 2004). A reduction in FMRP—whatever the cause—is thus expected to have a 2-fold effect on neurons: a loss of transcriptional control over synaptic mRNA pools and an inability to suppress hyperactive mGluR5 signaling. In Fragile X syndrome, FMRP activity is reduced at the level of translation (Jin and Warren 2000). In AD, on the other hand, it is possible that oligomer-induced clusters of mGluR5 stimulate a feedback-insensitive loss of FMRP activity. Functionally, this potential impact of oligomers would be equivalent to eliminating FMRP protein, as in Fragile X. Two expected AD-like consequences of the hypothesized mGluR5 cluster-mediated repression of FMRP activity are (i) the upregulation of Arc, which is under direct FMRP repression (Iacoangeli et al. 2008; Zalfa et al. 2003) and (ii) the alteration of synapse morphology through destabilization of PSD-95 mRNA, which is stabilized through a direct interaction with FMRP (Zalfa et al. 2007). Finally, because mGluR5 stimulates FMRP to permit translation of APP (Westmark and Malter 2007) an oligomer-induced loss in FMRP activity would also be expected to reinforce the effects of mGluR5 clustering by increasing APP translation and subsequent Aβ production.
Though the symptoms of AD and Fragile X are by no means identical, the loss of FMRP activity through either transcriptional or functional mechanisms could account for the similarities between these diseases with respect to synapses. To advance this hypothesis a step further, the significant similarities between Fragile X syndrome and the oligomer-induced synaptic deficits in AD may warrant the consideration of AD as an adult-onset oligomer-induced manifestation of an autism spectrum disorder.
Connection between AD and diabetes
Insulin signaling (i.e., the series of intracellular signaling cascades initiated by insulin receptor activation) in the aging brain is another area that may provide a mechanistic link between a healthy brain and one that develops AD. Given the positive effect of insulin signaling on synaptic function (Chiu et al. 2008; van der Heide et al. 2006) and memory (Marks et al. 2009), there is a burgeoning interest in a role for insulin in dementia. Epidemiological studies reveal an increased prevalence of dementia in individuals with diabetes, including an approximately 2-fold increase in the risk for developing AD (Leibson et al. 1997; Ott et al. 1999). Aberrant insulin signaling in the absence of diabetes also increases the risk of dementia (Peila et al. 2004). Mouse models of diabetes exhibit AD-like pathology, including elevated levels of Aβ and tau phosphorylation, which can be reduced by insulin treatment (Jolivalt et al. 2008), while inducing Type 1 or 2 diabetes in AD transgenic mice exacerbates AD and diabetic phenotypes (Jolivalt et al. 2010; Ke et al. 2009; Plaschke et al. 2010; Takeda et al. 2010), indicating that common underlying mechanisms such as insulin dysfunction link diabetes and AD.
Oligomeric Aβ antagonizes insulin signaling, as oligomer-bound neurons exhibit reduced insulin receptor activity and a reduction in synaptic insulin receptors (De Felice et al. 2009; Zhao et al. 2008). On the other hand, cognitive impairment by Aβ oligomers in a mouse model can be protected against by inhibiting the PI3K-Akt-mTOR pathway (Caccamo et al. 2010), while insulin activation of this signaling pathway mediates several processes integral to synapse function, including receptor trafficking (Huang et al. 2004), synaptic plasticity (van der Heide et al. 2006), and protein synthesis (Lee et al. 2005). Consistent with a co-antagonistic relationship between oligomer toxicity and protective insulin signaling, pretreatment of neurons with insulin protects against oligomer binding and toxicity (De Felice et al. 2009; Zhao et al. 2009).
The age-dependent decline in insulin signaling (Fernandes et al. 2001) could therefore increase the susceptibility of neurons to oligomer toxicity, providing a possible mechanism for the development of sporadic AD. In this mechanism, oligomeric Aβ could further hinder the already-waning neuroprotective effect of insulin signaling, resulting in synapse loss and neuronal death. This hypothesis for the induction of sporadic AD predicts that methods of strengthening insulin signaling could potentially stave off AD pathogenesis if administered during the appropriate phase of disease development.
Conclusion
At a fundamental level, developing an understanding of how specific molecules and events create brain states in which memory storage cannot occur will allow us to gain great insight into the mechanisms of normal information storage in the human brain. To this end, the evolution of the amyloid cascade hypothesis to focus on the toxic action of oligomeric Aβ has highlighted the importance of determining the molecular mechanisms by which these species contribute to AD. As details emerge regarding the toxic pathways actuated by Aβ oligomers, shared mechanisms between AD and other diseases affecting the organization of the brain can be recognized. We expect these forthcoming connections to promote both theoretical and experimental inroads into the oligomeric basis of AD and potential disease-modifying therapies.
References
Alarcon JM, Brito JA, Hermosilla T, Atwater I, Mears D, Rojas E (2006) Ion channel formation by Alzheimer’s disease amyloid β-peptide (Aβ40) in unilamellar liposomes is determined by anionic phospholipids. Peptides 27:95–104
Balducci C, Beeg M, Stravalaci M, Bastone A, Sclip A, Biasini E, Tapella L, Colombo L, Manzoni C, Borsello T, Chiesa R, Gobbi M, Salmona M, Forloni G (2010) Synthetic amyloid-β oligomers impair long-term memory independently of cellular prion protein. Proc Natl Acad Sci USA 107:2295–2300
Barghorn S, Nimmrich V, Striebinger A, Krantz C, Keller P, Janson B, Bahr M, Schmidt M, Bitner RS, Harlan J, Barlow E, Ebert U, Hillen H (2005) Globular amyloid beta-peptide oligomer—a homogenous and stable neuropathological protein in Alzheimer’s disease. J Neurochem 95:834–847
Bear MF, Huber KM, Warren ST (2004) The mGluR theory of fragile X mental retardation. Trends Neurosci 27:370–377
Bernstein SL, Dupuis NF, Lazo ND, Wyttenbach T, Condron MM, Bitan G, Teplow DB, Shea JE, Ruotolo BT, Robinson CV, Bowers MT (2009) Amyloid-beta protein oligomerization and the importance of tetramers and dodecamers in the aetiology of Alzheimer’s disease. Nat Chem 1:326–331
Braak H, Braak E (1991) Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol 82:239–259
Caccamo A, Majumder S, Richardson A, Strong R, Oddo S (2010) Molecular interplay between mammalian target of rapamycin (mTOR), amyloid-β, and Tau: effects on cognitive impairments. J Biol Chem 285:13107–13120
Calella AM, Farinelli M, Nuvolone M, Mirante O, Moos R, Falsig J, Mansuy IM, Aguzzi A (2010) Prion protein and Aβ-related synaptic toxicity impairment. EMBO Mol Med 2:306–314
Chang L, Bakhos L, Wang Z, Venton DL, Klein WL (2003) Femtomole immunodetection of synthetic and endogenous amyloid-β oligomers and its application to Alzheimer’s disease drug candidate screening. J Mol Neurosci 20:305–313
Chen S, Yadav SP, Surewicz WK (2010) Interaction between human prion protein and amyloid-beta (Abeta) oligomers: role of N-terminal residues. J Biol Chem 285:26377–26383
Chiu SL, Chen CM, Cline HT (2008) Insulin receptor signaling regulates synapse number, dendritic plasticity, and circuit function in vivo. Neuron 58:708–719
Cisse M, Halabisky B, Harris J, Devidze N, Dubal DB, Sun B, Orr A, Lotz G, Kim DH, Hamto P, Ho K, Yu GQ, Mucke L (2011) Reversing EphB2 depletion rescues cognitive functions in Alzheimer model. Nature 469:47–52
Cleary JP, Walsh DM, Hofmeister JJ, Shankar GM, Kuskowski MA, Selkoe DJ, Ashe KH (2005) Natural oligomers of the amyloid-β protein specifically disrupt cognitive function. Nat Neurosci 8:79–84
De Felice FG, Velasco PT, Lambert MP, Viola K, Fernandez SJ, Ferreira ST, Klein WL (2007) Aβ oligomers induce neuronal oxidative stress through an N-methyl-d-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J Biol Chem 282:11590–11601
De Felice FG, Vieira MN, Bomfim TR, Decker H, Velasco PT, Lambert MP, Viola KL, Zhao WQ, Ferreira ST, Klein WL (2009) Protection of synapses against Alzheimer’s-linked toxins: insulin signaling prevents the pathogenic binding of Aβ oligomers. Proc Natl Acad Sci USA 106:1971–1976
Deshpande A, Kawai H, Metherate R, Glabe CG, Busciglio J (2009) A role for synaptic zinc in activity-dependent Aβ oligomer formation and accumulation at excitatory synapses. J Neurosci 29:4004–4015
Douglass AD, Vale RD (2005) Single-molecule microscopy reveals plasma membrane microdomains created by protein–protein networks that exclude or trap signaling molecules in T cells. Cell 121:937–950
Eckert A, Hauptmann S, Scherping I, Meinhardt J, Rhein V, Drose S, Brandt U, Fandrich M, Muller WE, Gotz J (2008) Oligomeric and fibrillar species of β-amyloid (Aβ42) both impair mitochondrial function in P301L tau transgenic mice. J Mol Med 86:1255–1267
Fernandes ML, Saad MJ, Velloso LA (2001) Effects of age on elements of insulin-signaling pathway in central nervous system of rats. Endocrine 16:227–234
Fonseca AC, Resende R, Oliveira CR, Pereira CM (2010) Cholesterol and statins in Alzheimer’s disease: current controversies. Exp Neurol 223:282–293
Frackowiak J, Zoltowska A, Wisniewski HM (1994) Non-fibrillar β-amyloid protein is associated with smooth muscle cells of vessel walls in Alzheimer disease. J Neuropathol Exp Neurol 53:637–645
Gandy S, Simon AJ, Steele JW, Lublin AL, Lah JJ, Walker LC, Levey AI, Krafft GA, Levy E, Checler F, Glabe C, Bilker WB, Abel T, Schmeidler J, Ehrlich ME (2010) Days to criterion as an indicator of toxicity associated with human Alzheimer amyloid-β oligomers. Ann Neurol 68:220–230
Geng L, Zhang HL, Peng HB (2009) The formation of acetylcholine receptor clusters visualized with quantum dots. BMC Neurosci 10:80
Georganopoulou DG, Chang L, Nam JM, Thaxton CS, Mufson EJ, Klein WL, Mirkin CA (2005) Nanoparticle-based detection in cerebral spinal fluid of a soluble pathogenic biomarker for Alzheimer’s disease. Proc Natl Acad Sci USA 102:2273–2276
Gimbel DA, Nygaard HB, Coffey EE, Gunther EC, Lauren J, Gimbel ZA, Strittmatter SM (2010) Memory impairment in transgenic Alzheimer mice requires cellular prion protein. J Neurosci 30:6367–6374
Glabe CG (2008) Structural classification of toxic amyloid oligomers. J Biol Chem 283:29639–29643
Glenner GG, Murphy MA (1989) Amyloidosis of the nervous system. J Neurol Sci 94:1–28
Gong Y, Chang L, Viola KL, Lacor PN, Lambert MP, Finch CE, Krafft GA, Klein WL (2003) Alzheimer’s disease-affected brain: presence of oligomeric Aβ ligands (ADDLs) suggests a molecular basis for reversible memory loss. Proc Natl Acad Sci USA 100:10417–10422
Gouras GK, Tsai J, Naslund J, Vincent B, Edgar M, Checler F, Greenfield JP, Haroutunian V, Buxbaum JD, Xu H, Greengard P, Relkin NR (2000) Intraneuronal Aβ42 accumulation in human brain. Am J Pathol 156:15–20
Haass C, Selkoe DJ (2007) Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer’s amyloid beta-peptide. Nat Rev Mol Cell Biol 8:101–112
Hardy JA, Higgins GA (1992) Alzheimer’s disease: the amyloid cascade hypothesis. Science 256:184–185
Hertel C, Terzi E, Hauser N, Jakob-Rotne R, Seelig J, Kemp JA (1997) Inhibition of the electrostatic interaction between β-amyloid peptide and membranes prevents β-amyloid-induced toxicity. Proc Natl Acad Sci USA 94:9412–9416
Hsieh H, Boehm J, Sato C, Iwatsubo T, Tomita T, Sisodia S, Malinow R (2006) AMPAR removal underlies Aβ-induced synaptic depression and dendritic spine loss. Neuron 52:831–843
Huang CC, Lee CC, Hsu KS (2004) An investigation into signal transduction mechanisms involved in insulin-induced long-term depression in the CA1 region of the hippocampus. J Neurochem 89:217–231
Iacoangeli A, Rozhdestvensky TS, Dolzhanskaya N, Tournier B, Schutt J, Brosius J, Denman RB, Khandjian EW, Kindler S, Tiedge H (2008) On BC1 RNA and the fragile X mental retardation protein. Proc Natl Acad Sci USA 105:734–739
Irwin SA, Patel B, Idupulapati M, Harris JB, Crisostomo RA, Larsen BP, Kooy F, Willems PJ, Cras P, Kozlowski PB, Swain RA, Weiler IJ, Greenough WT (2001) Abnormal dendritic spine characteristics in the temporal and visual cortices of patients with fragile-X syndrome: a quantitative examination. Am J Med Genet 98:161–167
Jin P, Warren ST (2000) Understanding the molecular basis of fragile X syndrome. Hum Mol Genet 9:901–908
Jin M, Shepardson N, Yang T, Chen G, Walsh D, Selkoe DJ (2011) Soluble amyloid β-protein dimers isolated from Alzheimer cortex directly induce Tau hyperphosphorylation and neuritic degeneration. Proc Natl Acad Sci USA 108(14):5819–5824
Jolivalt CG, Lee CA, Beiswenger KK, Smith JL, Orlov M, Torrance MA, Masliah E (2008) Defective insulin signaling pathway and increased glycogen synthase kinase-3 activity in the brain of diabetic mice: parallels with Alzheimer’s disease and correction by insulin. J Neurosci Res 86:3265–3274
Jolivalt CG, Hurford R, Lee CA, Dumaop W, Rockenstein E, Masliah E (2010) Type 1 diabetes exaggerates features of Alzheimer’s disease in APP transgenic mice. Exp Neurol 223:422–431
Kang JE, Lim MM, Bateman RJ, Lee JJ, Smyth LP, Cirrito JR, Fujiki N, Nishino S, Holtzman DM (2009) Amyloid-β dynamics are regulated by orexin and the sleep-wake cycle. Science 326:1005–1007
Kayed R, Head E, Thompson JL, Mcintire TM, Milton SC, Cotman CW, Glabe CG (2003) Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science 300:486–489
Ke YD, Delerue F, Gladbach A, Gotz J, Ittner LM (2009) Experimental diabetes mellitus exacerbates tau pathology in a transgenic mouse model of Alzheimer’s disease. PLoS One 4:e7917
Kessels HW, Nguyen LN, Nabavi S, Malinow R (2010) The prion protein as a receptor for amyloid-β. Nature 466:E3–E4 discussion E4-5
Kim SH, Markham JA, Weiler IJ, Greenough WT (2008) Aberrant early-phase ERK inactivation impedes neuronal function in fragile X syndrome. Proc Natl Acad Sci USA 105:4429–4434
Klein WL, Krafft GA, Finch CE (2001) Targeting small Aβ oligomers: the solution to an Alzheimer’s disease conundrum? Trends Neurosci 24:219–224
Klein WL, Lacor PN, De Felice FG, Ferreira ST (2007) Molecules that disrupt memory circuits in Alzheimer’s disease: the attack on synapses by Aβ oligomers (ADDLs). In: Bontempi B, Silva AJ, Christen Y (eds) Memories: molecules and circuits. Springer, Berlin
Knowles JK, Rajadas J, Nguyen TV, Yang T, Lemieux MC, Vander Griend L, Ishikawa C, Massa SM, Wyss-Coray T, Longo FM (2009) The p75 neurotrophin receptor promotes amyloid-beta(1-42)-induced neuritic dystrophy in vitro and in vivo. J Neurosci 29:10627–10637
Koffie RM, Meyer-Luehmann M, Hashimoto T, Adams KW, Mielke ML, Garcia-Alloza M, Micheva KD, Smith SJ, Kim ML, Lee VM, Hyman BT, Spires-Jones TL (2009) Oligomeric amyloid β associates with postsynaptic densities and correlates with excitatory synapse loss near senile plaques. Proc Natl Acad Sci USA 106:4012–4017
Kovacs GG, Head MW, Hegyi I, Bunn TJ, Flicker H, Hainfellner JA, Mccardle L, Laszlo L, Jarius C, Ironside JW, Budka H (2002) Immunohistochemistry for the prion protein: comparison of different monoclonal antibodies in human prion disease subtypes. Brain Pathol 12:1–11
Kuo YM, Emmerling MR, Vigo-Pelfrey C, Kasunic TC, Kirkpatrick JB, Murdoch GH, Ball MJ, Roher AE (1996) Water-soluble Aβ (N-40, N-42) oligomers in normal and Alzheimer disease brains. J Biol Chem 271:4077–4081
Lacor PN (2007) Advances on the understanding of the origins of synaptic pathology in AD. Curr Genomics 8:486–508
Lacor PN, Buniel MC, Chang L, Fernandez SJ, Gong Y, Viola KL, Lambert MP, Velasco PT, Bigio EH, Finch CE, Krafft GA, Klein WL (2004) Synaptic targeting by Alzheimer’s-related amyloid β oligomers. J Neurosci 24:10191–10200
Lacor PN, Buniel MC, Furlow PW, Clemente AS, Velasco PT, Wood M, Viola KL, Klein WL (2007) Aβ oligomer-induced aberrations in synapse composition, shape, and density provide a molecular basis for loss of connectivity in Alzheimer’s disease. J Neurosci 27:796–807
Laferla FM, Green KN, Oddo S (2007) Intracellular amyloid-β in Alzheimer’s disease. Nat Rev Neurosci 8:499–509
Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, Liosatos M, Morgan TE, Rozovsky I, Trommer B, Viola KL, Wals P, Zhang C, Finch CE, Krafft GA, Klein WL (1998) Diffusible, nonfibrillar ligands derived from Aβ1-42 are potent central nervous system neurotoxins. Proc Natl Acad Sci USA 95:6448–6453
Larson J, Lynch G, Games D, Seubert P (1999) Alterations in synaptic transmission and long-term potentiation in hippocampal slices from young and aged PDAPP mice. Brain Res 840:23–35
Lashuel HA, Lansbury PT Jr (2006) Are amyloid diseases caused by protein aggregates that mimic bacterial pore-forming toxins? Q Rev Biophys 39:167–201
Lauren J, Gimbel DA, Nygaard HB, Gilbert JW, Strittmatter SM (2009) Cellular prion protein mediates impairment of synaptic plasticity by amyloid-β oligomers. Nature 457:1128–1132
Laurén J, Gimbel DA, Nygaard HB, Gilbert JW, Strittmatter SM (2010) Laurén et al. reply. Nature 466:E4–E5
Lee CC, Huang CC, Wu MY, Hsu KS (2005) Insulin stimulates postsynaptic density-95 protein translation via the phosphoinositide 3-kinase-Akt-mammalian target of rapamycin signaling pathway. J Biol Chem 280:18543–18550
Leibson CL, Rocca WA, Hanson VA, Cha R, Kokmen E, O’Brien PC, Palumbo PJ (1997) The risk of dementia among persons with diabetes mellitus: a population-based cohort study. Ann NY Acad Sci 826:422–427
Leon WC, Canneva F, Partridge V, Allard S, Ferretti MT, Dewilde A, Vercauteren F, Atifeh R, Ducatenzeiler A, Klein W, Szyf M, Alhonen L, Cuello AC (2010) A novel transgenic rat model with a full Alzheimer’s-like amyloid pathology displays pre-plaque intracellular amyloid-β-associated cognitive impairment. J Alzheimers Dis 20:113–126
Lesne S, Koh MT, Kotilinek L, Kayed R, Glabe CG, Yang A, Gallagher M, Ashe KH (2006) A specific amyloid-β protein assembly in the brain impairs memory. Nature 440:352–357
Levy E, Carman MD, Fernandez-Madrid IJ, Power MD, Lieberburg I, Van Duinen SG, Bots GT, Luyendijk W, Frangione B (1990) Mutation of the Alzheimer’s disease amyloid gene in hereditary cerebral hemorrhage, Dutch type. Science 248:1124–1126
Li S, Hong S, Shepardson NE, Walsh DM, Shankar GM, Selkoe D (2009) Soluble oligomers of amyloid β protein facilitate hippocampal long-term depression by disrupting neuronal glutamate uptake. Neuron 62:788–801
Magdesian MH, Nery AA, Martins AH, Juliano MA, Juliano L, Ulrich H, Ferreira ST (2005) Peptide blockers of the inhibition of neuronal nicotinic acetylcholine receptors by amyloid beta. J Biol Chem 280:31085–31090
Magdesian MH, Carvalho MM, Mendes FA, Saraiva LM, Juliano MA, Juliano L, Garcia-Abreu J, Ferreira ST (2008) Amyloid-beta binds to the extracellular cysteine-rich domain of Frizzled and inhibits Wnt/beta-catenin signaling. J Biol Chem 283:9359–9368
Malchiodi-Albedi F, Contrusciere V, Raggi C, Fecchi K, Rainaldi G, Paradisi S, Matteucci A, Santini MT, Sargiacomo M, Frank C, Gaudiano MC, Diociaiuti M (2010) Lipid raft disruption protects mature neurons against amyloid oligomer toxicity. Biochim Biophys Acta 1802:406–415
Marks DR, Tucker K, Cavallin MA, Mast TG, Fadool DA (2009) Awake intranasal insulin delivery modifies protein complexes and alters memory, anxiety, and olfactory behaviors. J Neurosci 29:6734–6751
Masters CL, Beyreuther K (1987) Neuronal origin of cerebral amyloidogenic proteins: their role in Alzheimer’s disease and unconventional virus diseases of the nervous system. Ciba Found Symp 126:49–64
Mclaurin J, Chakrabartty A (1997) Characterization of the interactions of Alzheimer β-amyloid peptides with phospholipid membranes. Eur J Biochem 245:355–363
Mucke L, Masliah E, Yu GQ, Mallory M, Rockenstein EM, Tatsuno G, Hu K, Kholodenko D, Johnson-Wood K, Mcconlogue L (2000) High-level neuronal expression of abeta 1-42 in wild-type human amyloid protein precursor transgenic mice: synaptotoxicity without plaque formation. J Neurosci 20:4050–4058
Nunomura A, Tamaoki T, Tanaka K, Motohashi N, Nakamura M, Hayashi T, Yamaguchi H, Shimohama S, Lee HG, Zhu X, Smith MA, Perry G (2010) Intraneuronal amyloid β accumulation and oxidative damage to nucleic acids in Alzheimer disease. Neurobiol Dis 37:731–737
Oda T, Pasinetti GM, Osterburg HH, Anderson C, Johnson SA, Finch CE (1994) Purification and characterization of brain clusterin. Biochem Biophys Res Commun 204:1131–1136
Oddo S, Caccamo A, Tran L, Lambert MP, Glabe CG, Klein WL, Laferla FM (2006) Temporal profile of amyloid-β (Aβ) oligomerization in an in vivo model of Alzheimer disease. A link between Aβ and tau pathology. J Biol Chem 281:1599–1604
Ott A, Stolk RP, Van Harskamp F, Pols HA, Hofman A, Breteler MM (1999) Diabetes mellitus and the risk of dementia: the Rotterdam Study. Neurology 53:1937–1942
Park S, Park JM, Kim S, Kim JA, Shepherd JD, Smith-Hicks CL, Chowdhury S, Kaufmann W, Kuhl D, Ryazanov AG, Huganir RL, Linden DJ, Worley PF (2008) Elongation factor 2 and fragile X mental retardation protein control the dynamic translation of Arc/Arg3.1 essential for mGluR-LTD. Neuron 59:70–83
Peila R, Rodriguez BL, White LR, Launer LJ (2004) Fasting insulin and incident dementia in an elderly population of Japanese-American men. Neurology 63:228–233
Plaschke K, Kopitz J, Siegelin M, Schliebs R, Salkovic-Petrisic M, Riederer P, Hoyer S (2010) Insulin-resistant brain state after intracerebroventricular streptozotocin injection exacerbates Alzheimer-like changes in Tg2576 AβPP-overexpressing mice. J Alzheimers Dis 19:691–704
Podlisny MB, Ostaszewski BL, Squazzo SL, Koo EH, Rydell RE, Teplow DB, Selkoe DJ (1995) Aggregation of secreted amyloid β-protein into sodium dodecyl sulfate-stable oligomers in cell culture. J Biol Chem 270:9564–9570
Proctor DT, Coulson EJ, Dodd PR (2010) Reduction in post-synaptic scaffolding PSD-95 and SAP-102 protein levels in the Alzheimer inferior temporal cortex is correlated with disease pathology. J Alzheimers Dis 21:795–811
Rahimi F, Shanmugam A, Bitan G (2008) Structure-function relationships of pre-fibrillar protein assemblies in Alzheimer’s disease and related disorders. Curr Alzheimer Res 5:319–341
Renner M, Lacor PN, Velasco PT, Xu J, Contractor A, Klein WL, Triller A (2010) Deleterious effects of amyloid β oligomers acting as an extracellular scaffold for mGluR5. Neuron 66:739–754
Roher AE, Ball MJ, Bhave SV, Wakade AR (1991) β-amyloid from Alzheimer disease brains inhibits sprouting and survival of sympathetic neurons. Biochem Biophys Res Commun 174:572–579
Roher AE, Palmer KC, Yurewicz EC, Ball MJ, Greenberg BD (1993) Morphological and biochemical analyses of amyloid plaque core proteins purified from Alzheimer disease brain tissue. J Neurochem 61:1916–1926
Selkoe DJ (2008) Soluble oligomers of the amyloid beta-protein impair synaptic plasticity and behavior. Behav Brain Res 192:106–113
Shankar GM, Li S, Mehta TH, Garcia-Munoz A, Shepardson NE, Smith I, Brett FM, Farrell MA, Rowan MJ, Lemere CA, Regan CM, Walsh DM, Sabatini BL, Selkoe DJ (2008) Amyloid-β protein dimers isolated directly from Alzheimer’s brains impair synaptic plasticity and memory. Nat Med 14:837–842
Shrestha BR, Vitolo OV, Joshi P, Lordkipanidze T, Shelanski M, Dunaevsky A (2006) Amyloid β peptide adversely affects spine number and motility in hippocampal neurons. Mol Cell Neurosci 33:274–282
Shughrue PJ, Acton PJ, Breese RS, Zhao WQ, Chen-Dodson E, Hepler RW, Wolfe AL, Matthews M, Heidecker GJ, Joyce JG, Villarreal SA, Kinney GG (2010) Anti-ADDL antibodies differentially block oligomer binding to hippocampal neurons. Neurobiol Aging 31:189–202
Simonyi A, Schachtman TR, Christoffersen GR (2005) The role of metabotropic glutamate receptor 5 in learning and memory processes. Drug News Perspect 18:353–361
Snyder EM, Nong Y, Almeida CG, Paul S, Moran T, Choi EY, Nairn AC, Salter MW, Lombroso PJ, Gouras GK, Greengard P (2005) Regulation of NMDA receptor trafficking by amyloid-β. Nat Neurosci 8:1051–1058
Sturchler E, Galichet A, Weibel M, Leclerc E, Heizmann CW (2008) Site-specific blockade of RAGE-Vd prevents amyloid-beta oligomer neurotoxicity. J Neurosci 28:5149–5158
Takahashi RH, Almeida CG, Kearney PF, Yu F, Lin MT, Milner TA, Gouras GK (2004) Oligomerization of Alzheimer’s β-amyloid within processes and synapses of cultured neurons and brain. J Neurosci 24:3592–3599
Takeda S, Sato N, Uchio-Yamada K, Sawada K, Kunieda T, Takeuchi D, Kurinami H, Shinohara M, Rakugi H, Morishita R (2010) Diabetes-accelerated memory dysfunction via cerebrovascular inflammation and Aβ deposition in an Alzheimer mouse model with diabetes. Proc Natl Acad Sci USA 107:7036–7041
Tomiyama T, Nagata T, Shimada H, Teraoka R, Fukushima A, Kanemitsu H, Takuma H, Kuwano R, Imagawa M, Ataka S, Wada Y, Yoshioka E, Nishizaki T, Watanabe Y, Mori H (2008) A new amyloid β variant favoring oligomerization in Alzheimer’s-type dementia. Ann Neurol 63:377–387
Tomiyama T, Matsuyama S, Iso H, Umeda T, Takuma H, Ohnishi K, Ishibashi K, Teraoka R, Sakama N, Yamashita T, Nishitsuji K, Ito K, Shimada H, Lambert MP, Klein WL, Mori H (2010) A mouse model of amyloid β oligomers: their contribution to synaptic alteration, abnormal tau phosphorylation, glial activation, and neuronal loss in vivo. J Neurosci 30:4845–4856
Townsend M, Mehta T, Selkoe DJ (2007) Soluble Aβ inhibits specific signal transduction cascades common to the insulin receptor pathway. J Biol Chem 282:33305–33312
Tu JC, Xiao B, Naisbitt S, Yuan JP, Petralia RS, Brakeman P, Doan A, Aakalu VK, Lanahan AA, Sheng M, Worley PF (1999) Coupling of mGluR/Homer and PSD-95 complexes by the Shank family of postsynaptic density proteins. Neuron 23:583–592
Van Der Heide LP, Ramakers GM, Smidt MP (2006) Insulin signaling in the central nervous system: learning to survive. Prog Neurobiol 79:205–221
Vigo-Pelfrey C, Lee D, Keim P, Lieberburg I, Schenk DB (1993) Characterization of β-amyloid peptide from human cerebrospinal fluid. J Neurochem 61:1965–1968
Walsh DM, Tseng BP, Rydel RE, Podlisny MB, Selkoe DJ (2000) The oligomerization of amyloid β-protein begins intracellularly in cells derived from human brain. Biochemistry 39:10831–10839
Walsh DM, Klyubin I, Fadeeva JV, Cullen WK, Anwyl R, Wolfe MS, Rowan MJ, Selkoe DJ (2002) Naturally secreted oligomers of amyloid β protein potently inhibit hippocampal long-term potentiation in vivo. Nature 416:535–539
Wang HW, Pasternak JF, Kuo H, Ristic H, Lambert MP, Chromy B, Viola KL, Klein WL, Stine WB, Krafft GA, Trommer BL (2002) Soluble oligomers of β amyloid (1-42) inhibit long-term potentiation but not long-term depression in rat dentate gyrus. Brain Res 924:133–140
Wang Q, Walsh DM, Rowan MJ, Selkoe DJ, Anwyl R (2004) Block of long-term potentiation by naturally secreted and synthetic amyloid β-peptide in hippocampal slices is mediated via activation of the kinases c-Jun N-terminal kinase, cyclin-dependent kinase 5, and p38 mitogen-activated protein kinase as well as metabotropic glutamate receptor type 5. J Neurosci 24:3370–3378
Westmark CJ, Malter JS (2007) FMRP mediates mGluR5-dependent translation of amyloid precursor protein. PLoS Biol 5:e52
Wong PT, Schauerte JA, Wisser KC, Ding H, Lee EL, Steel DG, Gafni A (2009) Amyloid-β membrane binding and permeabilization are distinct processes influenced separately by membrane charge and fluidity. J Mol Biol 386:81–96
Yun SH, Trommer BL (2010) Fragile X mice: reduced long-term potentiation and N-methyl-d-aspartate receptor-mediated neurotransmission in dentate gyrus. J Neurosci Res 89:176–182
Zalfa F, Giorgi M, Primerano B, Moro A, Di Penta A, Reis S, Oostra B, Bagni C (2003) The fragile X syndrome protein FMRP associates with BC1 RNA and regulates the translation of specific mRNAs at synapses. Cell 112:317–327
Zalfa F, Eleuteri B, Dickson KS, Mercaldo V, De Rubeis S, Di Penta A, Tabolacci E, Chiurazzi P, Neri G, Grant SG, Bagni C (2007) A new function for the fragile X mental retardation protein in regulation of PSD-95 mRNA stability. Nat Neurosci 10:578–587
Zampagni M, Evangelisti E, Cascella R, Liguri G, Becatti M, Pensalfini A, Uberti D, Cenini G, Memo M, Bagnoli S, Nacmias B, Sorbi S, Cecchi C (2010) Lipid rafts are primary mediators of amyloid oxidative attack on plasma membrane. J Mol Med 88:597–608
Zhao WQ, De Felice FG, Fernandez S, Chen H, Lambert MP, Quon MJ, Krafft GA, Klein WL (2008) Amyloid β oligomers induce impairment of neuronal insulin receptors. FASEB J 22:246–260
Zhao WQ, Lacor PN, Chen H, Lambert MP, Quon MJ, Krafft GA, Klein WL (2009) Insulin receptor dysfunction impairs cellular clearance of neurotoxic oligomeric Aβ. J Biol Chem 284:18742–18753
Zhao WQ, Santini F, Breese R, Ross D, Zhang XD, Stone DJ, Ferrer M, Townsend M, Wolfe AL, Seager MA, Kinney GG, Shughrue PJ, Ray WJ (2010) Inhibition of calcineurin-mediated endocytosis and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors prevents amyloid β oligomer-induced synaptic disruption. J Biol Chem 285:7619–7632
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Wilcox, K.C., Lacor, P.N., Pitt, J. et al. Aβ Oligomer-Induced Synapse Degeneration in Alzheimer’s Disease. Cell Mol Neurobiol 31, 939–948 (2011). https://doi.org/10.1007/s10571-011-9691-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10571-011-9691-4