
Abstract
Elemental ratios of calcium (Ca), strontium (Sr) and barium (Ba) in plant tissue have been used to help identify sources of Ca in trees. However, recent work has clearly shown that ratios of Ca:Sr or Ca:Ba in plant tissues differ from soil values and discrimination factors (alteration of elemental ratios of Ca, Sr and Ba) have been established to account for differences between soil and foliar values. In this study, concentrations of Ca, Sr and Ba (and molar ratios) in bulk deposition, throughfall, soil water and stream water were measured along with tree chemistry and biomass (foliage, branches, bark, wood, roots and litter) at a forested catchment in central Ontario, Canada. Concentrations of Ca, Sr and Ba vary greatly among the eight dominant tree species and plant tissue and molar ratios of Ca:Sr or Ca:Ba vary more than tenfold. In general Ca:Sr ratios were highest in foliage (up to 3,175) and lowest in wood (low as 91) and the mass-weighted Ca:Sr ratio of the total tree biomass was approximately 340. The molar ratio of Ca:Sr in bulk precipitation was ~820, increased to over 1,000 in throughfall and then decreased steadily through the various soil horizons, falling to 320 in the B horizon. Conservative mass balances for the forest floor for Ca, Sr and Ba suggest that more than half the Ca, Sr and Ba annual uptake by trees is obtained from mineral soil, which is consistent with the observation that the weighted-average ratios of Ca:Sr, Ca:Ba or Ba:Sr in the forest biomass fall between values in water draining the forest floor and water draining the rooting zone (B horizon). This work shows that discrimination between Ca, Sr and Ba is primarily internal to trees and discrimination factors between soil and foliage should be interpreted with caution.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Catchment mass balance estimates (Likens et al. 1996; Thimonier et al. 2000; Watmough et al. 2005a) and soil resurveys (Tamm and Hallbacken 1986; Lawrence et al. 1997; Lapenis et al. 2004) in eastern North America and parts of Europe indicate that there have been large losses of exchangeable soil calcium (Ca) in recent decades. Losses of Ca have been attributed primarily to a combination of increased Ca leaching brought about by elevated acid deposition (Likens et al. 1998) and uptake and removal of Ca in biomass during harvesting (Federer et al. 1989; Johnson et al. 1992). The loss of Ca from soil can have numerous adverse impacts on forest ecosystems including reduced growth due to Ca limitation or aluminum (Al) or manganese (Mn) toxicity (Horsley et al. 2000; Schaberg et al. 2001), decreased tree resistance to cold or insects (Hawley et al. 2006) and broader ecological impacts such as reduced mollusc populations or decreased avian success (Hamburg et al. 2003; Skeldon et al. 2007).
Given the importance of Ca to forest ecosystems there is a need to better understand the biogeochemical cycling of Ca in forest ecosystems. To this end, Ca:Sr and to a lesser extent Ca:Ba ratios and Sr stable isotope ratios have been used to distinguish the various sources of Ca using mixing models (Dasch et al. 2006; Drouet and Herbauts 2008). These calculations assume that Ca, Sr and/or Ba have similar geochemical behaviour. In some ecosystem processes, such as ion exchange on clays, Ca and Sr behave almost identically (Appelo and Postma 1993), whereas Ca may be retained preferentially over Sr on soil organic matter (Baes and Bloom 1988). Until recently, most studies have assumed that the Ca:Sr ratio did not change appreciably upon plant uptake (Runia 1987; Blum et al. 2002), but recent studies have challenged this notion by showing that Ca:Sr isotopes may vary substantially across various tree tissues (Poszwa et al. 2000; Watmough and Dillon 2003; Bullen and Bailey 2005; Drouet and Herbauts 2008). If Ca:Sr discrimination occurs in plants, then mixing models that determine end member contributions from Ca:Sr ratios and 87Sr:86Sr ratios are inaccurate because the assumption of Ca and Sr mass balance does not hold.
In an attempt to circumvent the impact of uptake on Ca:Sr or Ca:Ba ratios in tree tissues, discrimination factors (alteration of elemental ratios of Ca, Sr and Ba between tree tissues of the same species or between soil and tree tissues of various species) have been estimated for a number of tree species (Dasch et al. 2006; Blum et al. 2008, 2012). Drouet and Herbauts (2008) evaluated the discrimination of Ca compared to Sr in beech (Fagus sylvatica L.) and oak (Quercus robur L.) stands in Europe and found that Ca:Sr discrimination between exchange sites on soil organic matter and bole wood was relatively small and that most discrimination occurred between the bole and foliage. Dasch et al. (2006) reported that various plant species accumulate Ca and Sr in foliage at a constant proportion relative to their nutrient source and established discrimination factors for Ca:Sr (foliage relative to source) of 1.14, 1.16 and 1.9 for sugar maple (Acer saccharum Marshall), yellow birch (Betula alleghaniensis Britt.) and American beech (Fagus grandifolia Ehrh.), respectively. Blum et al. (2012) confirmed these discrimination factors and calculated discrimination factors for additional tree species including white birch (Betula papyrifera Marshall), pin cherry (Prunus pensylvanica L.) and red maple (Acer rubrum L.). Accounting for the reported discrimination factor in sugar maple, Blum et al. (2008) found that across six sites with large difference in foliar Ca:Sr ratios in sugar maple, foliar Ca:Sr ratios (and 87Sr:86Sr) were very similar to the organic (Oie) horizon reportedly confirming that internal recycling of Ca between litter and soil organic horizons and vegetation dominate annual uptake in northern hardwood ecosystems.
Foliage, however, typically represents less than 5 % of the above ground forest biomass. If elemental ratios in foliage differ markedly from other tree tissues this may alter the interpretation of Ca:Sr discrimination between soil and plants that is based only on foliar or leaf litter tissue. The objective of this study was to calculate fluxes of Ca, Sr and Ba in water as it passes through a forest ecosystem along with molar ratios (Ca:Sr, Ca:Ba and Sr:Ba) in water and tree tissues to test the hypotheses that foliar and soil chemistry alone cannot be used to identify the source of Ca, Sr and Ba in trees and that trees are accessing Ca, Sr and Ba from both the upper organic soil horizons and mineral soil.
Materials and methods
Site description
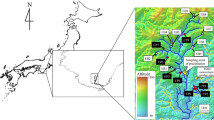
The Plastic Lake catchment is located in Haliburton County, Ontario (45°11′N, 78°50′W), about 20 km south of Dorset, Ontario (Fig. 1). The mean annual temperature for the Dorset area is 4.8 °C and the mean annual precipitation is 1,100 mm (Environment Canada 2002 30-year average 1971–2000). Approximately 80 % of the upland area in the Plastic 1 (PC1) sub-catchment drains through a Sphagnum-conifer swamp (Fig. 1) before discharging to the 32 ha headwater lake. The soils in the catchment are weakly developed podzols that occur on the well drained upland slopes, while histosols (organic soils) and gleysols occur in the wetlands, depressions and stream channels (Lozano 1987; Soil Classification Working Group 1998; WRB 2007). The average soil depth is only 0.5 m and bedrock outcrops consisting of granitized biotite, hornblende, and gneiss cover approximately 10 % of the area (Dillon and Lazerte 1992). The vegetation is a mixed forest comprised mainly of white pine (Pinus strobus L.), eastern hemlock (Tsuga canadensis L.) and red maple on the upland slopes. Other upland tree species include red oak (Quercus rubra L.), white birch, poplar (Populus tremuloides Michx.), American beech, balsam fir (Abies balsamea L. (Mill)). White cedar (Thuja occidentalis L.) and black spruce (Picea mariana Mill.) are present in the stream valleys and lower areas, and the dominant tree species in the wetland is white cedar. Mass balance studies at this site have indicated that large losses of Ca have occurred since 1980, but that current losses approximate inputs of Ca in deposition and mineral weathering, such that the catchment is approaching steady state (Watmough and Dillon 2004).

Location of the Plastic Lake catchment in central Ontario
Monitoring and sample collection
Bulk deposition was collected in a small clearing located adjacent to the PC1 sub-catchment using an open container fitted with a Teflon-lined funnel (diameter 25 cm) covered with 80 μm Nitex mesh to prevent contamination from debris between September 1, 2002 and August 30, 2003. Samples were collected for chemical analysis every 1–2 weeks. Stream discharge was measured continuously in the two streams (PC1-08 and PC1) using weirs and stilling wells with float devices connected to chart recorders. Discharge was calculated based on established stage-discharge relationships for each of the weirs (Scheider et al. 1983). Samples were collected from the stream outflow sampling locations in acid-washed Teflon bottles using clean techniques every 1–2 weeks, except when there was no flow.
Canopy throughfall was collected from spring through autumn every 2–4 weeks using 20 collectors situated in the upland forest of PC1-08. The collectors consisted of 8-cm diameter funnels with 80-μm mesh fitted to pails lined with plastic sample bags tested to ensure no metal contamination occurred. Winter throughfall samples were collected approximately monthly using open buckets. Litterfall was collected using 20 traps (0.5 m × 0.5 m) located adjacent to the throughfall collectors. The traps remained in place all year and the litter was collected 4 times (September, November, March, June) annually. Litter traps were well drained and there was no evidence of decomposition in the collected litter. Soil water was collected from five zero-tension lysimeter nests in the upland slopes of the PC1-08 subcatchment, which were installed in the mid 1980s. The lysimeters consisted of wooden frames draining through polyethylene tubing to plastic containers lined with plastic sample bags, which again were tested to ensure no metal contamination occurred. Soil water was collected from the organic (LFH), Ae and B horizons where those horizons existed. Soil water samples were collected every 2–4 weeks when sufficient soil water flow occurred.
Samples of roots, bark, wood, stems and leaves were collected from the eight dominant tree species present in the Plastic Lake catchment. Biomass estimates were obtained from the destructive sampling of trees growing outside the catchment in 1983 (methods are fully described in Watmough and Dillon 2004).
Chemical analysis
All water samples (bulk precipitation, throughfall, soil water, stream water and lake water) were passed through a coarse filter (80 μm) to remove large particulate matter. Throughfall samples were bulked into 4 replicate samples per collection period. More than 80 % of samples were sub-sampled and passed through a 0.45 μm filter in order to determine the proportion of “dissolved” Ca, Sr and Ba. Concentrations of the fine 0.45 μm filtered samples were approximately equal to the coarse filtered samples, indicating metals were almost entirely in the “dissolved” form (the <0.45 μm size fraction includes both dissolved as well as colloidal forms). Samples were acidified to <pH 2 with ultra pure HNO3 (Optima, Fisher Scientific) prior to analysis. Vegetation (washed in deionized water), soil and sediment samples were oven dried (65 °C) for at least 72 h and ground using a Wiley Mill. Vegetation samples were cold digested with HNO3 overnight and digested under reflux at 100 °C for 8 h. Sediment samples were digested in 50 % HNO3 under reflux at 100 °C for 8 h. All digests were diluted and filtered through a Whatman 42 filter paper before analysis. Samples were analysed using an Element II Thermo-Finnigan high resolution single collector Inductively Coupled Plasma Mass Spectrometer (ICP-MS). An internal standard (5 ppb Rh) was used to correct for instrument drift. Recoveries were determined using the standard reference material Trace Metals in Natural Water (NIST-1640), and each of the reported certified metals typically had a recovery within ±15 %.
Calcium, Sr and Ba forest pool size and mass balance calculations
Pools of Ca, Sr and Ba in the forest and elemental ratios in various tissues were calculated using chemical concentrations in the respective vegetation tissue of each species multiplied by biomass estimates for that tissue and values were then summed for each species (Watmough and Dillon 2004). Calcium, Sr and Ba fluxes in the streams were calculated using the continuous hydrology record and the periodic concentration measurements, whereby the measured concentrations are substituted into the period between sampling events (Dillon et al. 1982). Bulk deposition and throughfall fluxes were estimated by summing the input of Ca, Sr and Ba for each collection period between September 1, 2002 and August 30, 2003. Since the sample volumes obtained from the lysimeters are not quantitative, Ca, Sr and Ba fluxes in the soil water were calculated using discharge from the upland PC1-08 stream. This will underestimate fluxes in the upper soil horizons as water is lost through evapotranspiration from soil below the organic and A horizons and hence mass balances for the forest floor will be conservative. Differences in Ca, Sr and Ba concentrations and Ca:Sr, Ca:Ba and Ba:Sr ratios among species for each tree tissue (foliage, branch, wood, bark, root) and among various litterfall components (foliage, branch, seeds) were assessed using a non-parametric one way analysis of variance test, Kruskal–Wallis. The post hoc test, Dunn’s Method, was used for multiple comparisons between species for each tree tissue.
Results
Tree chemistry
Concentrations of Ca, Sr and Ba varied tremendously both among and within tree species (Table 1). Calcium concentrations tended to be highest in the bark and lowest in wood (Table 1). For example, the highest Ca concentration was measured in the bark of red oak (>22,000 mg kg−1) and the lowest concentration was measured in the wood of white pine (250 mg kg−1) (Table 1). Differences among tree tissues in Sr and Ba were less pronounced than for Ca, but within tissues differences among tree species in Sr and Ba were much greater than for Ca (Table 1). For example, Ca concentrations in foliage were between 3,500 and 8,500 mg kg−1, whereas Sr concentrations were between 3 and 58 mg kg−1, and Ba concentrations were between 4 and 147 mg kg−1 (Table 1). Consequently molar ratios of Ca:Sr, Ca:Ba and Sr:Ba also varied greatly and variation among species tended to be greater than variation within tree species (Table 1). Molar ratios of Ca:Sr for example, were generally highest in foliage and lowest in wood and were as high as 3,175 in white pine foliage and as low as 91 in the wood of poplar (Table 1). A similar amount of variability was observed in the Ca:Ba ratio, which was as high as 4,560 in the foliage of white pine and as low as 33 in the wood of white birch (Table 1). Molar ratios of Sr:Ba were much less variable within and among tree species; having a maximum value of 1.7 in white pine branches and a minimum value of 0.2 in the bark of black spruce and white birch (Table 1).
Throughfall, soil water and stream water chemistry
Annual average measured concentrations of Ca, Sr and Ba were lowest in bulk precipitation and increased by approximately 2.5 (Sr and Ba) to 4 times (Ca) in throughfall (Table 2). As a result Ca:Sr and Ca:Ba ratios increased by approximately 50 % in throughfall relative to bulk deposition, whereas there was little change in the Sr:Ba ratio (Table 2). The concentration of Ca in water draining the organic horizon (LFH) was about 4 times higher than throughfall, whereas concentrations of Sr and Ba increased about ten and sixfold, respectively leading to a large decrease in the Ca:Sr and Ca:Ba ratio and a smaller decrease in the Sr:Ba ratio. Concentrations of Ca in mineral soil water (A and B horizons) were approximately half the concentration draining the forest floor, whereas there was little change in Sr and Ba concentrations such that Ca:Sr and Ca:Ba ratios in mineral soil water decreased by about 50 % compared with water draining the forest floor. Measured Ca concentrations in the stream draining the upland catchment (PC1-08) were slightly lower than the B horizon, while Sr and Ba concentrations were comparable, resulting in a further decrease in the Ca:Sr and Ca:Ba ratios, but no change in the Sr:Ba ratio (Table 2). Concentrations of Ca, Sr and Ba in water draining the wetland (PC1-03 and PC1) were the most variable, but the mean values were higher than PC1-08 (Table 2).
Litterfall
Litterfall represents the largest annual flux of Ca, Sr and Ba to the forest floor (Fig. 2). Calcium concentrations were lower in tree reproductive parts (seeds, flowers) compared with foliage and branches, although the bulk of the litterfall is comprised of foliar tissue (Table 3). Differences in Sr and Ba concentration among litterfall tissues was minimal and ratios of Ca:Sr and Ca:Ba and were lower in reproductive parts compared with foliage or branches, but there was no difference in Sr:Ba ratios (Table 3).

Estimated Ca, Ba and Sr annual fluxes (mg m−2) and pools along with Ca:Sr, Ca:Ba and Sr:Ba ratios in vegetation and water at the Plastic Lake catchment
Calcium, Sr and Ba pools, fluxes and budgets
The annual Ca input in throughfall was approximately 3 times higher than bulk deposition and represented about one quarter of the total Ca input to the forest floor (Fig. 2). Calcium inputs in litterfall were estimated to be 2,172 mg m−2 year−1 (Fig. 2). A conservative estimate for the Ca flux from the forest floor was 1,427 mg m−2 year−1. The Ca flux decreased down the soil profile and was estimated to be 566 mg m−2 year−1 in water draining the rooting zone (B horizon). The Ca flux in water draining the upland catchment (PC1-08) was similar to the B horizon, but approximately 2 times higher in PC1-03 and PC-1, due to a large increase in Ca concentration that occurred following summer drought (Fig. 3). Assuming no net forest growth, no Ca accumulation in the forest floor and the enrichment in throughfall Ca is due solely to leaching and not dry deposition, it is estimated that of the trees annual Ca demand (excluding fine root turnover: litterfall + foliar leaching = 2,594 mg m−2 year−1), 1,398 mg m−2 year−1 (or 54 %) could be supplied from the forest floor, with the remainder being supplied from the mineral soil. Calcium weathering rates at PC1 have been estimated to be around 200–270 mg m−2 year−1 (Watmough and Dillon 2003), which when combined with bulk deposition (231 mg m−2 year−1) approximate losses from the rooting zone or the upland-draining stream.

Monthly volume weighted concentrations of Ca, Sr, Ba and SO4 at two sample points a PC1-03 and b PC1 draining the wetland at the Plastic Lake catchment
Similar patterns to Ca are evident in the Sr and Ba flux and budget estimates (Fig. 2). It is estimated that of the forests annual uptake (litterfall + leaching = 12.3 mg m−2 year−1 (Ba) and 7.2 mg m−2 year−1 (Sr)), only 3.1 mg m−2 year−1 (25 %) and 3.4 mg m−2 year−1 (47 %), respectively could be supplied by the forest floor. Assuming no net loss of Sr and Ba from the upland catchment, Sr and Ba weathering rates would have to be approximately 2.7 mg m−2 year−1 and 10.9 mg m−2 year−1, respectively.
The mass weighted Ca:Sr ratio in litterfall was approximately 830, which is 1.5 times higher than the Ca:Sr ratio in water draining the forest floor (Fig. 2). The mass-weighted Ca:Sr ratio for the forest was 341, which is much lower than litterfall due to lower Ca:Sr ratios in branches, bark, roots and particularly stemwood compared with litter and falls between the Ca:Sr ratios in organic (LFH) (569) and B horizon (324) water (Fig. 2). A very similar pattern was observed for Ca:Ba ratios; the Ca:Ba ratio of litterfall was approximately 2 times higher than water draining the LFH horizon and the mass weighted ratio for the forest trees (195) was between that of the LFH (392) and B horizon (152). Molar ratios of Sr:Ba were much less variable, although Sr:Ba ratios in litterfall were 1.4 times higher than the LFH, and the mass weighted Sr:Ba ratio (0.6) similarly fell between the Sr:Ba ratio in the LFH layer (0.7) and the B horizon (0.5) (Fig. 2).
Discussion
Several previous studies have shown that elemental ratios of Ca, Sr and Ba vary within tree tissues of individual species and among species (Poszwa et al. 2000; Watmough and Dillon 2003; Bullen and Bailey 2005; Drouet and Herbauts 2008). The reasons for this discrimination are largely unknown. Drouet and Herbauts (2008) and Gierth et al. (1998) argue that discrimination occurs in the passage of Ca and Sr through the endodermis that separates conducting vessels from the leaf mesophyll. It has also been suggested that discrimination between Ca and Sr may occur by ion exchange processes during ascent in the xylem (Clarkson 1984), although Momoshima and Bondietti (1990) reported that the Ca:Sr ratio was constant at different trunk heights of Picea rubens (Sarg.). Gulpin et al. (1995) suggested that discrimination may occur due to preferential binding in Ca-oxalate precipitated in bark or other tissues.
Despite obvious differences in chemistry within and among tree species, previous studies have shown that discrimination among Ca, Sr and Ba in tree tissues appears to be constant within a species (Dasch et al. 2006; Drouet and Herbauts 2008; Blum et al. 2008, 2012). For example, Drouet and Herbauts (2008) reported that discrimination factors from bole wood to leaves for Sr:Ca, Ba:Ca and Sr:Ba in European beech in Belgian forests were 2.99 ± 0.6, 4.64 ± 1.76 and 0.70 0.22, respectively. Similar results were reported between other tissues in beech as well as for Quercus robur (Drouet and Herbauts 2008). Drouet and Herbauts (2008) further argued that discrimination factors from wood to leaves were relatively constant over a large range in soil Ca levels and that elemental ratios in foliage may be corrected with a constant factor (Sr:Ca: 2.99 for beech and 6.54 for oak) to account for differences in ratios between foliage and soil water. Discrimination factors for Ca:Sr between foliage and the forest floor have been established for a number of eastern North American tree species and range between 1.16 (sugar maple) and 1.90 (red maple) (Dasch et al. 2006; Blum et al. 2008, 2012) and have been used to suggest that mineral soil is not a major source of Ca to foliage because Ca:Sr ratios decrease with soil depth and are even more distinct from foliar values (Blum et al. 2008).
In the present study, the Ca:Sr ratio in water draining the forest floor is 569 ± 165 compared with 838 ± 520 in litterfall, which can be used to approximate foliar ratios as Ca:Sr ratios differ little between foliage and litter (Fig. 2; Blum et al. 2008). The Ca:Sr ratios in mineral soil (A and B horizons) water are even lower than the forest floor and simply interpreting Ca:Sr ratios in soil water and foliage would suggest that the forest floor is the major source of Ca to the canopy with an overall discrimination factor of around 1.5, which is within the range of six eastern North American species reported by Blum et al. (2012). Similarly, if we re-evaluate data for 11 sugar maple stands in central Ontario (Watmough et al. 2005b) that vary widely in pH (4.2–7.7) and Ca:Sr ratios in A horizon soil (organic (LFH) horizon not always present), there is a linear relationship between foliar Ca:Sr ratios and water-extractable Ca:Sr ratios in A horizon soil, with a discrimination factor of 1.51 (Fig. 4). This discrimination factor for sugar maple is higher than the value (1.16) reported by Blum et al. (2008), which is likely because soil water in the A horizon rather than the organic horizon was used and mineral soil horizons have lower Ca:Sr ratios than organic horizons (e.g. this study; Blum et al. 2008). Similar results were observed for Ca:Ba ratio with an apparent discrimination factor of 1.56, between sugar maple foliage and A horizon soil (Fig. 4) compared with approximately 1.9 between foliage and the organic horizon at Plastic Lake. Drouet and Herbauts (2008) reported that discrimination factors between wood and foliage for oak and beech were around 3.0–3.7, but also suggested that Ba and Ca had contrasting behaviour (unlike Sr and Ca), which does not appear to be the case at Plastic Lake.

Relationship between foliar Ca:Sr ratios in sugar maple and Ca:Sr ratios in water extracts from A horizons at 11 stands in central Ontario (see Watmough et al. 2005b). Dashed line shows the 1:1 ratio
Mass balance estimates for Plastic Lake however, indicate that the mineral soil is an important source of Ca in litter (and trees). Mass balances were constructed by assuming annual inputs to the forest floor were litterfall (2,172 mg m−2 year−1) and throughfall (653 mg m−2 year−1) and leaching losses (1,427 mg m−2 year−1) were calculated based on concentrations of Ca in LFH leachate multiplied by runoff recorded at PC1-08. This approach assumes that all evapotranspiration occurs from the forest floor, which is not the case and hence this mass balance is conservative as leaching losses from the forest floor will be higher than assumed in this study. The results of this mass balance indicate that annually, the forest floor gains at most 1,398 mg m−2 year−1 of Ca. The annual Ca demand of the forest (fine root turnover was excluded) was estimated as litterfall + (throughfall − bulk deposition) and was 2,594 mg m−2 year−1. This approach will slightly overestimate the annual forest demand as it is assumed that all the Ca in throughfall is from foliar leaching and additional dry deposition is excluded. Previous work at Plastic Lake however has indicated that based on Na-enrichment in throughfall, dry deposition would increase bulk deposition by just 13 % (Neary and Gizyn 1994). Results of this mass balance, which assumes no accumulation in the forest floor or biomass suggest that at least 1,196 mg m−2 year−1 (or 46 %) of the foliar Ca must be obtained from mineral soil. This estimate is similar to the estimate reported by Miller et al. (1993) for a spruce-fir forest in the north eastern United States based on Sr-isotope data. Calculations for Sr and Ba, indicate that at best, the forest floor can supply 36 % and 16 %, respectively of the forests annual uptake.
In order to explain the apparent discrepancy between mass budgets and elemental ratios in soil and plant tissue, it is necessary to take into account other tree tissues. For example, Ca:Sr ratios are much higher in litter (foliage) compared with branches, bark, roots and wood. Taking into account the biomass of the various tree tissues, the forest at PC1 has a mass weighted Ca:Sr ratio of 341, which falls between the values observed in the forest floor (569 + 165) and B horizon (324 + 157). In this study it is assumed that ratios in soil water reflect the source of elements to trees as no exchangeable concentrations were available. While there may be slight differences in selectivity between the soil exchange complex and soil water for Ba relative to Ca and Sr (Drouet and Herbauts 2008), other studies have shown that elemental ratios of Ca, Sr and Ba in soil water and the soil exchange complex are generally comparable (Blum et al. 2002; Drouet and Herbauts 2008). The variability in Ca:Sr ratios, particularly in soil water precludes a reliable estimate of the relative contribution from various soil horizons, but results are consistent with mineral soil providing a substantial amount of the forests’ annual Ca, Sr and Ba uptake. While there is currently no net increase in forest biomass (Watmough and Dillon 2004), individual trees at Plastic Lake are growing (increase in biomass is offset by tree death) and Ca, Sr and Ba are being deposited in all plant tissues and hence foliar (or litter) Ca:Sr ratios will not reflect the relative annual uptake of these elements. Similar results are obtained for both Ca:Ba and Sr:Ba ratios as mass-weighted values calculated for the forest biomass (195 (Ca:Ba) and 0.6 (Sr:Ba)) fall between values observed in water draining the forest floor (392 ± 176 (Ca:Ba) and 0.7 ± 0.2 (Sr:Ba)) and water in the B horizon (152 ± 93 (Ca:Ba) and 0.5 ± 0.1 (Sr:Ba)).
Rather than interpreting similarities in Ca:Sr (and Ba) ratios in foliar (or litterfall) and upper organic horizons to indicate that upper organic horizons are the major sources of Ca to trees (Blum et al. 2008), the similarity in ratios may occur because the Ca:Sr ratio in the upper organic horizons is determined primarily by Ca:Sr ratio in litterfall and throughfall. Moreover, both Poszwa et al. (2000) and Drouet and Herbauts (2008) reported that molar ratios of Ca:Sr and Ca:Ba in roots were similar to soil horizons in which they are located. In the present study, roots were sampled from the upper mineral soil (A horizon) and elemental ratios of Ca:Sr, Ca:Ba and Sr:Ba are close to values observed in this horizon. Roots were thoroughly cleaned so soil contamination is unlikely, but cannot be totally discounted.
If the mass balance is extended to the mineral soil and it is assumed that inputs to mineral soil are solely from leachate from the organic horizon (1,427 mg m−2 year−1) and Ca weathering, which is estimated between 200 and 270 mg m−2 year−1 (Watmough and Dillon 2004) and losses represent the flux in the B horizon (566 mg m−2 year−1), the results indicate that the gain in soil Ca is between 1,061 and 1,131 mg m−2 year−1. As soils at Plastic Lake have lost Ca over the past few decades (Watmough and Dillon 2004), rather than Ca accumulating in soil, it is more likely that these values approximate the amount of Ca supplied by mineral soil to trees supporting the contention that mineral soils are an important source of Ca to trees at Plastic Lake. Finally, assuming that the Plastic Lake catchment is approaching steady state (Watmough and Dillon 2004), this approach suggests that weathering estimates for Sr and Ba are 2.7 and 10.9 mg m−2 year−1, respectively. Blum et al. (2002) reported that the Ca:Sr ratios in digests that dissolve primarily Ca silicate minerals (e.g. hornblende, plagioclase) are between 170 and 279. The mass balance constructed here suggests a weathering Ca:Sr ratio at Plastic Lake between 162 and 219, consistent with the observation that plagioclase and hornblende are the primary weathering sources of Ca (Watmough and Dillon 2004; Watmough and Aherne 2008).
Conclusions
Similar to previous studies, elemental ratios of Ca, Sr and Ba vary tremendously, both within and among tree species and ratios in litterfall are much closer to values observed in upper organic horizons as opposed to mineral layers. However, mass balance estimates at Plastic Lake show that the mineral soil must be a major source of Ca, Sr and Ba to trees and this may only be reconciled by considering mass-weighted ratios in the entire forest, which fall between values observed in the upper organic horizon and the B horizon. Given the large variability in elemental ratios in soil water and vegetation some slight discrimination between soil and trees cannot be discounted, but considering only ratios in litter (or foliage) and soil water would provide an erroneous conclusion that mineral soils at Plastic Lake provide little to no Ca in trees and discrimination factors used in other regions should be interpreted with caution.
References
Appelo CAJ, Postma D (1993) Geochemistry, groundwater and pollution. A.A. Balkema, Rotterdam, p 536
Baes AU, Bloom PR (1988) Exchange of alkaline earth cations in soil organic matter. Soil Sci 146:6–14
Blum JD, Klaue A, Nezat CA, Driscoll CT, Johnson CE, Siccama TG, Eagar C, Fahey TJ, Likens GE (2002) Mycorrhizal weathering of apatite as an important calcium source in base poor forest ecosystems. Nature 417:729–731
Blum JD, Dasch AA, Hamburg SP, Yanai RD, Arthur MA (2008) Use of foliar Ca/Sr discrimination and Sr-87/Sr-86 ratios to determine soil Ca sources to sugar maple in a northern hardwood forest. Biogeochem 87:287–296
Blum JD, Hamburg SP, Yanai RD, Arthur MA (2012) Determination of foliar Ca/Sr discrimination factors for six tree species and implications for Ca sources in northern hardwood forests. Plant Soil 356:303–314
Bullen TD, Bailey SW (2005) Identifying calcium sources at an acid deposition-impacted spruce forest: a strontium isotope, alkaline-earth element multi-tracer approach. Biogeochem 74:63–99
Clarkson DT (1984) Calcium transport between tissues and its distribution in the plant. Plant Cell Environ 7:449–456
Dasch A, Blum JD, Eagar C, Fahey TJ, Driscoll CT, Siccama TG (2006) The relative uptake Ca and Sr into tree foliage using a whole-watershed calcium addition. Biogeochem 80:21–41
Dillon PJ, Lazerte BD (1992) Response of the plastic lake catchment, Ontario, to reduced sulfur deposition. Environ Pollut 77(2–3):211–217
Dillon PJ, Jeffries DS, Scheider WA (1982) The use of calibrated lakes and watersheds for estimating atmospheric deposition near a large point source. Water Air Soil Pollut 18:241–258
Drouet T, Herbauts J (2008) Evaluation of the mobility and discrimination of Ca, Sr and Ba in forest ecosystems: consequence of the use of alkaline-earth element ratios as tracers of Ca. Plant Soil 302:105–124
Environment Canada (2013) Canada climate archive. http://climate.weather.gc.ca
Federer CA, Hornbeck JW, Tritton LM, Martin CW, Pierce RS, Smith CT (1989) Long-term depletion of calcium and other nutrients in eastern US forests. Environ Manage 13:593–601
Gierth M, Stelzer R, Lehmann H (1998) Endodermal Ca and Sr partitioning in needles of the European larch (Larix decidua (L.) Mill.). J Plant Physiol 152(1):25–30
Gulpin M, Turk S, Fink S (1995) Ca nutrition in conifers. Z Pflanzenernahr Bodenk 158:519–527
Hamburg SP, Yanai RD, Arthur MA, Blum JD, Siccama TG (2003) Biotic control of calcium cycling in northern hardwood forests: acid rain and aging forests. Ecosystems 6:399–406
Hawley GJ, Schaberg PG, Eager C, Borer CH (2006) Calcium addition at the Hubbard Brook experimental forest reduced winter injury to red spruce in a high-injury year. Can J For Res 36(10):2544–2549
Horsley SB, Long RP, Bailey SW, Hallett RA, Hall TJ (2000) Factors associated with the decline disease of sugar maple on the Allegheny plateau. Can J For Res 30:1365–1378
IUSS Working Group WRB (2007) World Reference Base for Soil Resources 2006, first update 2007. World Soil Resources Reports No. 103. FAO, Rome
Johnson CE, Johnson AH, Siccama TG (1992) Whole-tree clear–cutting effects on exchangeable cations and soil acidity. Soil Sci Soc Amer J 55:502–508
Lapenis AG, Lawrence GB, Andreev AA, Bobrov AA, Torn MS, Harden JW (2004) Acidification of forest soil in Russia: from 1893 to present. Glob Biogeochem Cycles. doi:10.1029/2003GB002107
Lawrence GB, David MB, Bailey SW, Shortle WC (1997) Assessment of soil calcium status in red spruce forests in the north-eastern United States. Biogeochem 38:19–39
Likens GE, Driscoll CT, Buso DC (1996) Long-term effects of acid rain: response and recovery of a forest ecosystem. Science 272:244–246
Likens GE, Driscoll CT, Buso DC, Siccama TG, Johnson CE, Lovett GM, Fahey TJ, Reiners WA, Ryan DF, Martin CW, Bailey SW (1998) The biogeochemistry of calcium at Hubbard Brook. Biogeochem 41:89–173
Lozano F (1987) Physical and chemical properties of the soils at the southern biogeochemistry study site Report BGC-018. Dorset Research Centre, Ontario Ministry of the Environment, Dorset (Ont.)
Miller EK, Blum JD, Friedland AJ (1993) Determination of soil exchangeable-cation loss and weathering rates using Sr isotopes. Nature 362:438–441
Momoshima N, Bondietti EA (1990) Cation binding in wood: applications to understanding historical changes in divalent cation availability to red spruce. Can J For Res 20:1840–1849
Neary AJ, Gizyn WI (1994) Throughfall and stemflow chemistry under deciduous and coniferous forest canopies in South-Central Ontario. Can J For Res 24:1089–1100
Poszwa A, Dambrine E, Pollier B, Atteia O (2000) A comparison between Ca and Sr cycling in forest ecosystems. Plant Soil 225:299–310
Runia LT (1987) Strontium and calcium distribution in plants: effects on palaeodietary studies. J Archaeol Sci 14(6):599–608
Schaberg PG, DeHayes DH, Hawley GJ (2001) Anthropogenic calcium depletion: a unique threat to forest ecosystem health? Ecosys Health 7:214–228
Scheider WA, Cox CM, Scott LD (1983) Hydrological data for lakes and watersheds in the Muskoka-Haliburton study area (1976–1980). Ontario Ministry of Environment Data Report 83/6
Skeldon M, Vadeboncoeur MA, Hamburg SP, Blum JD (2007) Terrestrial gastropod distribution in the northern hardwood forest: the role of calcium and forest composition. Can J Zool 85:994–1007
Soil Classification Working Group (1998) The Canadian System of Soil Classification Publ. 1646 (Revised):187
Tamm CO, Hallbacken L (1986) Changes in soil pH over a 50-year period under different forest canopies, in SW Sweden. Water Air Soil Pollut 31:337–341
Thimonier A, Dupouey JL, Tacon FL (2000) Recent losses of base cations from soils of Fagus sylvatica L. stands in northeastern France. Ambio 29:314–321
Watmough SA, Aherne J (2008) Estimating calcium weathering rates and future lake calcium concentrations in the Muskoka-Haliburton region of Ontario. Can J Fish Aquat Sci 65:821–833
Watmough SA, Dillon PJ (2003) Mycorrhizal weathering in base-poor forests. Nature 423:823–824
Watmough SA, Dillon PJ (2004) Major element fluxes from a coniferous catchment in central Ontario, 1983–1999. Biogeochem 67:369–398
Watmough SA, Aherne J, Alewell C, Arp P, Bailey S, Clair T, Dillon PJ, Duchesne L, Eimers MC, Fernandez I, Foster N, Larseen T, Miller E, Mitchell MJ, Page S (2005a) Sulphate, nitrogen and base cation budgets at 21 forested catchments in Canada, the United States and Europe. Environ Monitor Assess 109:1–36
Watmough SA, Dillon PJ, Epova EN (2005b) Metal partitioning and uptake in central Ontario forests. Environ Pollut 134:493–502
Acknowledgments
This work was funded through by the Canadian Water Network and the Natural Sciences and Engineering Council of Canada (NSERC). The author gratefully acknowledges the help from staff at the Ontario Ministry of Environment Dorset Environmental Centre and various Trent students who helped with data collection and analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: E. Veldkamp.
Rights and permissions
About this article
Cite this article
Watmough, S.A. Calcium, strontium and barium biogeochemistry in a forested catchment and insight into elemental discrimination. Biogeochemistry 118, 357–369 (2014). https://doi.org/10.1007/s10533-013-9938-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-013-9938-x