
Abstract
The extent to which the diversity of forest-inhabiting bryophytes is related to forest management practices has been the subject of a recent study. However, the question of how these practices affect the spatial distribution pattern of bryophyte diversity throughout managed European forests remains unanswered. We surveyed bryophyte diversity in six large forest complexes in the Czech Republic. Our sites encompass a range of managed forest stands (including various temporal phases such as clearings and young forests) across a gradient of environmental conditions. In general, the bryophyte species richness in managed forests was less than in unmanaged forests. Only unmanaged forest stands that provided suitable substrates such as large dead woody debris and large diameter beeches (Fagus sylvatica) were able to support rare bryophyte species. Mature managed forests with simplified tree structure, dominated by either deciduous or coniferous species shared similar bryophyte species richness (α-diversity) and pool (γ-diversity). It appears that forest management, which leads to the loss of old-growth forest attributes, may result in low-diversity bryophyte communities regardless of tree species composition. Nevertheless, bryophyte species turnover (β-diversity) was spatially uniform in mature managed forests dominated by conifers, but quite variable in those dominated by deciduous tree species. Managed forests with diverse tree structure and composition approximated the species pool size of unmanaged forests, in spite of their small contribution to the total area of managed forests. Hence, forest management as commonly applied in the managed forests of the Czech Republic does not appear to be conducive to the persistence of diverse bryophyte assemblages. We suggest that some essential changes in forest management practices are necessary to maintain the conditions favorable to the preservation of bryophyte diversity in central European forests.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A sufficient area of old unmanaged forest with a humid microclimate and a sufficient quantity and variety of microhabitats on the surfaces of old trees and/or dead woody debris are considered major determinants of bryophyte diversity in European forests (Heilmann-Clausen et al. 2005; Fritz et al. 2008; Brunet et al. 2010). As forest management leads to a lack of old-growth forest attributes, bryophyte assemblages appear to be impoverished in most managed forests (Söderström 1988; Ódor and Standovár 2001; Friedel et al. 2006). However, we have a limited understanding as to how (and how extensively) the old-growth forest attributes should be preserved in managed forests to enhance their biodiversity (Bauhus et al. 2009). Many foresters have already recognized that some management practices may result in a decline in the diversity of bryophytes (as well as that of other forest-inhabiting taxa). However, they argue that these effects are spatially and temporarily restricted, as are the management practices themselves (Lindenmayer and Laurance 2012). According to this point of view, bryophytes have a large enough area of temporarily unmanaged forest, which sometimes includes the old trees and dead woody debris necessary for the maintenance of overall bryophyte diversity. This presumption may not stand up to scrutiny, because of the dispersal constraints of many sensitive and highly substrate-specialized bryophyte species (Snäll et al. 2005; Löbel et al. 2006; Löbel and Rydin 2010). We cannot, however, deny that relatively species-rich bryophyte communities have been able—at least temporarily—to persist in some types of managed forest (Fenton and Frego 2005; Lõhmus et al. 2007; Perhans et al. 2009).
Forest structure and tree species composition have been influenced by forest management for several centuries across central Europe (Grove 2002). Therefore, knowledge of which features of currently managed forests are natural and which artificial is less consistent than would be expected (Bengtsson et al. 2000). Because natural (unmanaged) forests occupy small areas and are usually fragmented into very small remnants (<50 ha), the idea of managing their development by simply allowing the occurrence of purely natural processes is illusory (Pickett and Thompson 1978). Moreover, most of these currently unmanaged forests have been managed in recent decades or centuries (Gillson and Willis 2004). These factors make the evaluation of the effects of forest management activities on bryophyte diversity even more difficult. Despite this, significant progress has been made over the last few years (Brunet et al. 2010; Ódor et al. 2013), but we still have little understanding of the potential of central European managed forests to provide valuable habitat for bryophytes. Previous studies that compared bryophyte communities in differently managed forests, or managed and unmanaged forests, were restricted to predefined type(s) of managed forest (e.g., Ódor and Standovár 2001). Therefore, these prior studies document bryophyte diversity patterns in particular segments of forest stands, not in managed forests as a whole.
In an attempt to expand our present knowledge, we designed a study to investigate differences in species richness (α-diversity), species turnover (β-diversity), and species pool (γ-diversity) of bryophyte communities in relation to environmental conditions and intensity of forest management in the most widespread forest types of the Czech Republic. We covered all the most widespread forest types, to truly reflect the intensity and temporal aspects of forest management currently found in central Europe. In particular, in our study, we focused on the substrate preferences of species that were specific to unmanaged or a particular managed forest type.
Materials and methods
Study areas
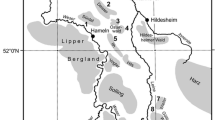
This study was carried out in six study areas (SA1–SA6) distributed across the Czech Republic to cover the widest possible ranges of the geography, elevation, and other relevant environmental gradients (e.g., geology) (Fig. 1; Table 1). Each study area was part of a large continuous forest patch with an area of 1.4–10.0 km2. The size and boundaries of each study area were designed to include a sufficient area of each of a predefined set of forest stand types (Fig. 2). The stands with spatially and vertically uniform (but not necessarily homogeneous) forest structure, and a size greater than 2 ha, were classified as core forest stands (grain-sized structure). However, in forest stands younger than 70 years old, the size of the cores started at 0.5 ha, because of the limited total area of some of these stands. Consequently, we associated each core stand with one of five types of forest stands, classified according to the predominant management practices, age structure, and tree species composition of forests within the Czech Republic, and in central Europe in general. Firstly, we distinguished three categories of mature forest (>70 years old): (a) nature reserves without regular forest management (hereafter referred to as unmanaged forests), (b) managed stands of deciduous or mixed tree species (mature deciduous forests), and (c) managed stands of coniferous species (mature coniferous forests). Secondly, we divided regularly managed stands younger than 70 years into two habitat types: (d) regularly managed stands of deciduous, mixed, and coniferous species 11–69 years old (young forests), and (e) clear-cut stands 2–10 years old (clearings). Finally, we established a category (f) termed as forest mosaics which combined all remaining forest stands, deemed “unclassifiable” as one of the previous habitat types because of insufficient area or habitat heterogeneity. The largest part of each study area was occupied by mature coniferous stands, followed by mature deciduous stands. The size of the nature reserves ranged from 10 to 100 ha, respectively. There was one nature reserve inside each study area, with the exception of SA4 and SA5, in which there were two and none, respectively. Therefore, the aim of having at least one example of all six forest stand types inside each study area was met in four study areas (SA1–SA4), while the two remaining study areas at the lowest elevations (SA5, SA6) met this aim only if they were coupled (Table 1).

Location of the six study areas

Division of the forests within the study areas into six categories, according to management type
Unmanaged forests are predominantly composed of beech (Fagus sylvatica) with an admixture of spruce (Picea abies), and fir (Abies alba), which both increase in prevalence with elevation. Ash (Fraxinus excelsior), maples (Acer pseudoplatanus, A. platanoides), and lime (Tilia platyphyllos) are other common native tree species. The only exception was unmanaged forests at the lowest elevations, which were dominated by oak (Quercus petraea) instead of beech. Mature deciduous forests were generally composed of beech and oak at higher and lower elevation, respectively, whereas monocultures of spruce and pine (Pinus sylvestris) generally made up mature coniferous forests. Mature managed forest stands were around 100 years old and rarely reaching 180 years. Small remnants of old managed forest (mostly classified as forest mosaics) have survived for more than 200 years. The age of currently unmanaged forests ranges between 150 and 400 years, according to forest management plans (provided by Ministry of Environment). These forests have mostly been protected since the first half of twentieth century, the earliest and latest having been protected since 1838 and 1964, respectively.
Forest continuity in the study areas was established according to the maps created during the first and second military mapping of the Austrian Empire during 1764–1768, and 1836–1852, respectively (archived in Czech Office for Surveying, Mapping and Cadastre, Prague).
Field survey
To cover the internal heterogeneity of the different forest stands in the study areas, we delineated one to four square sampling plots (proportional to the size of the plots), each 2,500 m2, in each type of forest stand in all study areas. We established 20 and 13 sampling plots in SA1–SA4 and SA5–SA6, respectively, for a total of 106 sampling plots (Table 1; for details, see Appendix 1 in ESM). In summer 2009 and 2010, in all sampling plots, we recorded all bryophyte species found 0–2 m up from the forest floor. We surveyed all substrates on the forest floor (e.g., soil, stones, stumps, lying dead wood), as well as live and dead-standing trees. Those individuals not identified in the field were collected and identified in the laboratory using microscopy and/or the help of experts. A few samples were identifiable only to genus (indicated by “sp.”). In the final dataset, we included only those records identified to genus, which clearly represented other species than those already mentioned in the list. The presence of each species at a particular plot was appended by the specification of occupied substrate(s) according to the list of 32 potential substrates, and subsequently grouped into the nine categories reported here. Species nomenclature and conservation status was taken from the Check List and Red List of bryophytes of the Czech Republic (Kučera et al. 2012).
Data analysis
We compared differences in species richness (α-diversity), species turnover (β-diversity), and total species pools (γ-diversity) in the different types of forest stands. The Jaccard dissimilarity index was used for the evaluation of community species turnover within forest stand types. Analysis of variance of both species richness and species turnover among types of forest stands was assessed by a non-parametric Kruskal–Wallis test (KW) at significance level p < 0.05, followed by an adjustment for multiple comparisons, according to Hochberg (1988). Exact unconditioned rarefaction curves were built for the determination of total species pools. Correlation was assessed by Pearson correlation coefficients, at a significance level of p < 0.05. Computations were performed in R (R Development Core Team 2013), using the “agricolae” (de Mendiburu 2014) and “vegan” packages (Oksanen et al. 2012).
Additionally, we looked at substrate differences among the bryophyte species associated exclusively with one forest management type (see Appendix 2 in ESM). The importance of the substrates was expressed as the sum of their contribution to the occurrence of the exclusive species. Each species had an equal total increment (=1) regardless of differences in species frequency, which was either ascribed to one or proportionally divided among more substrates, according to the occupation frequency of respective species.
Results
Bryophyte communities
In total, we found 108 species in the 106 sampling plots, comprised 78 mosses and 30 liverworts (Appendix 1 and 2 in ESM). Hypnum cupressiforme, the most widespread species, occurred in 103 sampling plots, whereas 25 species were each recorded in only one sampling plot. Species richness per sampling plot ranged from 5 to 38 species. The highest species richness was found in unmanaged forest within forest reserves, and the lowest species richness was generally found in clearings (Kruskal–Wallis test; p < 0.001; Fig. 3). Similarly, the mean contribution of liverworts to bryophyte species richness (α-diversity) was highest and lowest in unmanaged forests (18 %) and clearings (7 %), respectively, and ranged between 13 and 15 % in all other forest management stands.

Bryophyte species richness in particular plots in a given type of forest stand: A unmanaged forests, B mature deciduous forests, C mature coniferous forests, D young forests, E clearings, F forest mosaics. A mean, bottom, and upper quartiles, and deciles and outliers of species richness are depicted. Different letters indicate differences in species richness (p < 0.05)
With regard to species turnover, the most similar bryophyte assemblages occupied mature coniferous stands, followed by young forests, whereas spatial species turnover peaked in clearings (Kruskal–Wallis test; p < 0.001; Fig. 4). We identified 33 species inherent to specific forest stand type within one or more of the study areas (Fig. 5; Appendix 1 in ESM). Forest reserves, followed by clearings, contained the greatest number of species that were restricted to a particular forest stand type (Fig. 5) if we neglect to account for the differences in the number of plots surveyed in each particular forest stand type.

Bryophyte species turnover between pairs of plots within particular types of forest stands: A unmanaged forests, B mature deciduous forests, C mature coniferous forests, D young forests, E clearings, F forest mosaics. A mean, bottom, and upper quartiles, and deciles and outliers of species richness are depicted. Different letters indicate a difference in species composition (p < 0.05)

Substrate affinities of species recorded only in particular types of forest stands: A unmanaged forests, B mature deciduous forests, C mature coniferous forests, D young forests, E clearings, F forest mosaics
Although species pools of bryophytes varied among study areas, rarefaction curves convincingly show the extent to which the γ-diversity of managed forests is compromised when compared to forest reserves (Fig. 6). Only the forest mosaics approximated the species pool found in nature reserves. We detected a depletion of species pools in both deciduous and coniferous mature forest stands, with almost no difference between the two types (Fig. 6). The species pool recorded in young forests matched that found in mature managed forests, but clearing species pool did not.

Rarefaction curves for the comparison of bryophyte species pools in forest stand types. Intervals of standard deviations are indicated by dotted lines
With respect to the national red list of bryophytes, we found one endangered species (Anacomptodon splachnoides), one vulnerable species (Syzygiella autumnalis), two near-threatened species (Cephalozia catenulata, C. leucantha), and six species included in the affiliated attention list. All these species occurred predominantly in nature reserves, only rarely in managed forests, and never in clearings.
Substrate preference in unmanaged versus managed forests
Species restricted to a particular type of forest stand showed an affinity for certain substrates. The most remarkable was the difference between nature reserves and clearings (Fig. 5). However, different substrates were generally occupied either by species exclusive to nature reserves, or species found only in all other managed forests (Fig. 5). While most samples of species exclusive to nature reserves were found on thick-trunked deciduous trees and downed woody debris, species exclusive to managed forests occurred primarily on soil and stones. Accordingly, the ecological requirements of species restricted to particular forest stand types corresponded to the predicted environmental conditions in these forest stand types. For example, species associated with the bare soil surfaces of open stands (e.g., Bryum pallens, Campylopus flexuosus, Pohlia prolingera) occurred only in clearings, whereas species such as Alleniella complanata and Riccardia latifrons, which are associated with stands of little human disturbance, were repeatedly recorded in forest reserves only. In general, downed logs, and less often stumps and thick beech stems, were substrates preferentially inhabited by red-listed species, regardless of forest stand type.
Discussion
In the predominant stand types of managed forests in the Czech Republic, neither species richness nor the species pool of bryophyte communities aligned with those in unmanaged forest reserves (Figs. 3, 6). These results correspond to those of previous studies, which were mostly limited to select types of managed and unmanaged forests in other European countries: Denmark (Aude and Poulsen 2000), Estonia (Vellak and Paal 1999), Germany (Friedel et al. 2006), Hungary (Ódor and Standovár 2001), and Sweden (Söderström 1988). Apart from this general agreement, we found considerable differences in bryophyte diversity among particular types of managed forest stands. While the species richness and species pool observed in forest mosaics were not very different from those of forest reserves, a lack of bryophyte diversity was apparent in all other types of managed forest stands (Figs. 3, 6). This exceptional positioning of forest mosaics relative to managed forest stands could be explained by an inherent attribute supporting bryophyte diversity: higher environmental heterogeneity arising from rich forest structure and/or tree species composition, with the occasional occurrence of large old trees (Heilmann-Clausen et al. 2005; Brunet et al. 2010; Ódor et al. 2013). In addition, the spatial heterogeneity of environmental conditions in forest mosaics, and in other managed stand types as well, inadvertently benefited because of the large area covered by managed forests, which far exceed the area of unmanaged forests (10–100 ha). However, the occurrence of large dead wood components (logs) in forest mosaics was as infrequent as in other managed forests, which featured a similar lack of epixylic bryophyte species as was observed in other managed forests (Fig. 5; Appendix 1 in ESM).
The paucity of bryophyte assemblages was most remarkable in forest stands in the initial phases of the clear-cutting management cycle (clearings and young forests). The disappearance of both suitable substrates and stable humid conditions due to clear-cutting did not allow for the persistence of forest-dwelling bryophytes, especially liverworts, in these stands (Nelson and Halpern 2004; Fenton and Frego 2005). The retention of some living and dead trees after cutting to serve as structural legacies for diversity “lifeboating,” has been considered effective in some areas (Fenton and Frego 2005; Lindenmayer et al. 2012; Rudolphi et al. 2013), but less so in others (Nelson and Halpern 2004; Rosenvald and Lõhmus 2008). Nevertheless, even green-tree retention cutting is not a common method in managed forests in the Czech Republic.
The species turnover of bryophyte communities in mature coniferous forests was low, even though there was relatively high species richness per plot, and a broad gradient in environmental conditions resulting from the large total area of these forests (Figs. 3, 4). The rate of bryophyte community turnover was the only apparent difference in the diversity of coniferous and deciduous mature managed forests, which otherwise had similar mean species richness and total species pools (Figs. 3, 6). Pure coniferous forest represents a forest type that does not naturally occur anywhere along the elevation gradient encompassed by our study (Chytrý et al. 2012). Beech is, in contrast, considered an important or even dominant tree species of natural forest vegetation (Chytrý et al. 2012). The similarity of the mean bryophyte species richness that we found in deciduous and coniferous mature managed forests seems contrary to the expectation that native tree species forests would be more species rich than artificial forests (Hansen et al. 1991; Felton et al. 2010). However, the observed species richness may not indicate a contradiction to this expectation at all, but instead may demonstrate that the seemingly “close to natural” conditions of deciduous managed forests and forest reserves exempt from recent timber harvesting are not as natural as previously thought (Lõhmus et al. 2007).
In central Europe, deciduous forests have a predisposition for maintaining the naturalness of their forest environment more thoroughly than artificial coniferous forests (Bengtsson et al. 2000). Apart from differences in tree species composition, the rotation period in deciduous managed forests usually extends over several decades, relative to conifer forests, in which harvesting takes place earlier. In our study, the age of coniferous and deciduous mature managed forests averaged 94 and 124 years, respectively, while the oldest stands reached 124 and 182 years. The age of some deciduous managed forests overlapped the age of unmanaged forests, which ranged between 144 and 315 years and thus achieved the critical age threshold for diversity of some forest-dwelling taxa in beech-dominated forests, which is estimated to be between 100 and 220 years (Moning and Müller 2009; see also Fritz et al. 2008).
This result implies that forest age is not the only condition required for the (re-)establishment of bryophyte communities, unless it is accompanied by the recovery of the structural attributes which constitute suitable microhabitats for bryophytes (Nordén and Appelquist 2001; Fritz et al. 2008). A serious failure of bryophyte recovery due to ongoing forest management practices aiming for a simplification of forest structure and systematic removal of dead and windfall trees may result in unexpectedly low species richness even in old deciduous managed forests.
The purportedly low level of diversity in saproxylic beetles in European beech forests has recently been questioned (Müller et al. 2012). The authors demonstrated that beech forests should not be considered as species poor as is sometimes indicated, owing to the low number of specialist associated with beech (Müller et al. 2013). These forests are indeed able to host a considerable quantity of saproxylic diversity, providing that the forest contains more than 30–50 m3 of dead wood per hectare. This condition is, however, rarely present in most managed forests in Europe (Friedman and Walheim 2000; Müller and Büttler 2010).
The amount of time needed to recreating forest naturalness in previously managed forests may be surprisingly long or short, depending upon forest structure and composition at the beginning of the process, the frequency and extent of natural disturbances occurring during the process, and the aspects of the naturalness of interest (Vandekerkhove et al. 2005, 2009). Forest reserves included in our study have been protected from periods of less than half a century, to more than a century and a half (the forest reserve in study area 2 is the oldest in Europe, having been protected since 1838). Human intervention usually ceases several decades before the beginning of protection. On the other hand, not every intervention has ended immediately upon the implementation of protection, and some even continue today (e.g., occasional harvesting of snags and logs). The forest reserves situated at low elevations, and therefore near traditionally human-inhabited areas, were probably more heavily affected by forest management in the past than forest reserves in mountainous areas (Pokorný 2005; Kaplan et al. 2009).
Conclusions
According to expectations, bryophyte communities inhabiting large areas of managed forest were generally more species poor than those in unmanaged forests. Epixylic and other rare bryophyte species found suitable substrates mainly in unmanaged forests. However, with the exception of these bryophyte species, managed forests may host approximately the same level of bryophyte diversity as unmanaged forests, but only those managed forests with diverse tree structure and composition. Deciduous and coniferous mature managed forests have similar species richness and species pool of bryophytes. The higher potential for deciduous forests to host a rich bryophyte diversity may have been suppressed by forest management to nearly the same level as that of artificial pure coniferous forests. Hence, forest management in the form in which it is presently carried out in virtually all managed forest areas in the Czech Republic appears to be incompatible with the presence of rich bryophyte assemblages. Additionally, the repercussions of past forest management appears to still limit bryophyte diversity, even in many unmanaged forests. We thus can conclude that some essential changes in forest management practices are necessary, if forests are to provide appropriate conditions for the maintenance of bryophyte diversity in central European forests.
References
Aude E, Poulsen RS (2000) Influence of management on the species composition of epiphytic cryptogams in Danish Fagus forests. Appl Veg Sci 3:81–88
Bauhus J, Puettmann K, Messier C (2009) Silviculture for old-growth attributes. For Ecol Manag 258:525–537
Bengtsson J, Nilsson SG, Franc A, Menozzi P (2000) Biodiversity, disturbances, ecosystem function and management of European forests. For Ecol Manag 132:39–50
Brunet J, Fritz Ö, Richnau G (2010) Biodiversity in European beech forests—a review with recommendations for sustainable forest management. Ecol Bull 53:77–94
Chytrý M (2012) Vegetation of the Czech Republic: diversity, ecology, history and dynamics. Preslia 84:427–504
de Mendiburu F (2014) Statistical procedures for agricultural research. R—package, version 1:1–7
Felton A, Lindbladh M, Brunet J, Fritz Ö (2010) Replacing coniferous monocultures with mixed-species production stands: an assessment of the potential benefits for forest biodiversity in northern Europe. For Ecol Manag 260:933–947
Fenton NJ, Frego KA (2005) Bryophyte (moss and liverwort) conservation under remnant canopy in managed forests. Biol Conserv 122:417–430
Friedel A, von Oheimb G, Dengler J, Härdtle W (2006) Species diversity and species composition of epiphytic bryophytes and lichens—a comparison of managed and unmanaged forests in NE Germany. Feddes Repert 117:172–185
Friedman J, Walheim M (2000) Amount, structure, and dynamics of dead wood on managed forestland in Sweden. For Ecol Manag 131:23–36
Fritz Ö, Gustafsson L, Larsson K (2008) Does forest continuity matter in conservation?—a study of epiphytic lichens and bryophytes in beech forests of southern Sweden. Biol Conserv 141:655–668
Gillson L, Willis KJ (2004) ′As earth′s testimonies tell′: wilderness conservation in a changing world. Ecol Lett 7:990–998
Grove SJ (2002) Saproxylic insect ecology and the sustainable management of forest. Annu Rev Ecol Evol Syst 33:1–33
Hansen AJ, Spies TA, Swanson FJ, Ohmann JL (1991) Conserving biodiversity in managed forests. Bioscience 41:382–392
Heilmann-Clausen J, Aude E, Christensen M (2005) Cryptogam communities on decaying deciduous wood—does tree species diversity matter? Biodivers Conserv 14:2061–2078
Hochberg Y (1988) A sharper Bonferroni procedure for multiple tests of significance. Biometrika 75:800–803
Kaplan JO, Krumhardt KM, Zimmermann N (2009) The prehistoric and preindustrial deforestation of Europe. Quat Sci Rev 28:3016–3034
Kučera J, Váňa J, Hradílek Z (2012) Bryophyte flora of the Czech Republic: updated checklist and Red List and a brief analysis. Preslia 84:813–850
Lindenmayer DB, Laurance WF (2012) A history of hubris—cautionary lessons in ecologically sustainable forest management. Biol Conserv 151:11–16
Lindenmayer DB, Franklin JF, Lõhmus A, Baker SC, Bauhus J, Beese W, Brodie A, Kiehl B, Kouki J, Martínez Pastur G, Messier C, Neyland M, Palik B, Sverdrup-Tygeson A, Volney J, Wayne A, Gustafsson L (2012) A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv Lett 5:421–431
Löbel S, Rydin H (2010) Trade-offs and habitat constraints in the establishment of epiphytic bryophytes. Funct Ecol 24:887–897
Löbel S, Snäll T, Rydin H (2006) Metapopulation processes in epiphytes inferred from patterns of regional distribution and local abundance in fragmented forest landscapes. J Ecol 94:856–868
Lõhmus A, Lõhmus P, Vellak K (2007) Substratum diversity explains landscape-scale co-variation in the species-richness of bryophytes and lichens. Biol Conserv 135:405–414
Moning C, Müller J (2009) Critical forest age thresholds for the diversity of lichens, molluscs and birds in beech (Fagus sylvatica L.) dominated forests. Ecol Indic 9:922–932
Müller J, Bütler R (2010) A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. Eur J For Res 129:981–992
Müller J, Brunet J, Brin A, Bouget C, Brustel H, Bussler H, Förster B, Isacsson G, Köhler F, Lachat T, Gossner MM (2012) Implications from large-scale spatial diversity patterns of saproxylic beetles for the conservation of European Beech forests. Insect Conserv Diver 6:162–169
Müller J, Jarzabek-Müller A, Bussler H, Gossner MM (2013) Hollow beech trees as keystone structures for saproxylic beetles by analyses of functional and phylogenetic diversity. Anim Conserv 17:154–162
Nelson CR, Halpern CB (2004) Short-term effects of timber harvest and forest edges on ground-layer mosses and liverworts. Can J Bot 83:610–620
Nordén B, Appelquist T (2001) Conceptual problems of ecological continuity and its bioindicators. Biodivers Conserv 10:779–791
Ódor P, Standovár T (2001) Richness of bryophyte vegetation in near-natural and managed beech stands: the effects of management-induced differences in dead-wood. Ecol Bull 49:219–229
Ódor P, Király I, Tinya F, Bortignon F, Nascimbene J (2013) Patterns and drivers of species composition of epiphytic bryophytes and lichens in managed temperate forests. For Ecol Manag 306:256–265
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2012) Vegan: community ecology package. R—package, version 2.0–5
Perhans K, Appelgren L, Jonsson F, Nordin U, Söderström B, Gustafsson L (2009) Retention patches as potential refugia for bryophytes and lichens in managed forest landscapes. Biol Conserv 142:1125–1133
Pickett STA, Thompson JN (1978) Patch dynamics and the design of nature reserves. Biol Conserv 13:27–36
Pokorný P (2005) Role of man in the development of Holocene vegetation in Central Bohemia. Preslia 77:113–128
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, http://www.r-project.org/. Accessed Feb 2013
Rosenvald R, Lõhmus A (2008) For chat, when, and where is green-tree retention better than clear-cutting? a review of the biodiversity apsects. For Ecol Manag 255:1–15
Rudolphi J, Jönsson MT, Gustafsson L (2013) Biological legacies buffer local species extinction after logging. J Appl Ecol 51:53–62
Snäll T, Ehrlén J, Rydin H (2005) Colonization-extinction dynamics of an epiphyte metapopulation in a dynamic landscape. Ecology 86:106–115
Söderström L (1988) The occurrence of epixylic bryophyte and lichen species in an old natural and a managed forest stand in northeastern Sweden. Biol Conserv 45:169–178
Tolasz R (ed) (2007) Climate atlas of Czechia. Czech Hydrometerorological Institute, Prague
Vandekerkhove K, De Keersmaeker L, Baeté H, Walleyn R (2005) Spontaneous re-establishment of natural structure and related biodiversity in a previously managed beech forest in Belgium after 20 years of non intervention. For Snow Landsc Res 79:145–156
Vandekerkhove K, De Keersmaeker L, Menke N, Meyer P, Verschelde P (2009) When nature takes over from man: dead wood accumulation in previously managed oak and beech woodlands in North-western and Central Europe. For Ecol Manag 258:425–435
Vellak K, Paal J (1999) Diversity of bryophyte vegetation in some forest types in Estonia: a comparison of old unmanaged and managed forests. Biodivers Conserv 8:1595–1620
Acknowledgments
We thank K. Baltaziuk, L. Čížek, V. Iarema, A. Petrbok, and A. Tenčík for field assistance. GIS analyses were provided by T. Svoboda and P. Kukla, and some problematic records helped to determine J. Kučera. We thank J. Titus and Editage for revising the English. The Editor and anonymous referee kindly provided valuable comments and suggestions on the manuscript. We are grateful to the respective forest administrations for cooperation in our research. This study was supported by Grant SP/2d1/146/08 from the Ministry of the Environment of the Czech Republic.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T.G. Allan Green.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hofmeister, J., Hošek, J., Holá, E. et al. Decline in bryophyte diversity in predominant types of central European managed forests. Biodivers Conserv 24, 1391–1402 (2015). https://doi.org/10.1007/s10531-015-0863-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-015-0863-2