
Abstract
Biogeographical analyses are applied to skipper (Hesperioidea) presence/absence data from the Western Mediterranean mainland and the three largest islands (Sardinia, Corsica and Sicily) in order to identify potential conservation issues. The analyses performed on species, both collectively and individually, indicate that regional species richness and occurrence in the Mediterranean zone are largely predicted by latitude and area but that islands have impoverished faunas. Several species, predicted to be present on these islands from logistic regression of their continental distributions, are actually absent. The number of species predicted to be present from logistic regression analyses for each island, closely matched the number of species predicted to occur in regional-focused multiple regression analysis. This suggests that missing species have been identified. When compared with species that occur in Sicily and Corsica, the missing species are shown to differ for ecological traits, mainly those linked to altitudinal tolerance. No ecological distinctions were disclosed for Sardinian skippers suggesting a mainly stochastic colonisation. These results, and those from an analogous study carried out on Papilionoidea, point to Hesperioidea having (i) overall more impoverished faunas on islands and (ii) being subject to stochastic or historical colonisation events more than Papilionoidea. Species not predicted to occur on islands based on their mainland distributions and ecological traits, are foci for conservation attention. However, as many species becoming extinct on the islands may be irreplaceable, all species, in particular the Sardinian ones, deserve to be conserved.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Island faunas and floras are distinct from those on mainlands in several respects. Among these, species’ impoverishment is the most apparent feature of island communities with islands usually having fewer species compared to mainland regions that share similar characteristics (i.e., biotopes, environmental conditions and area). Impoverishment of islands is caused by two main factors: (i) lower colonisation rates of suitable patches owing to increased isolation by sea barriers, and (ii) higher extinction rates owing to smaller populations from a limited resource base and reduced rescue effect and gene flow (Williamson 1981; Whittaker 1998; Dapporto and Dennis 2009). For these reasons, island communities are often disharmonic with respect to those on nearby mainlands and contain assemblages distinguished by species with greater dispersal capacity and ecological tolerance (Whittaker 1998; Dapporto and Dennis 2009 for butterflies).
Island faunas are the result of both deterministic and stochastic events. Occasional island colonisation by a species is a stochastic event, but it is more likely in species showing high dispersal capability and environmental tolerance (Williamson 1981; Whittaker 1998). Clearly, when island isolation is small compared to the dispersal ability of a particular taxon, environmental factors are predicted to prevail in determining its occurrence; but, when isolation is much larger than dispersal capability, stochastic and historical elements underlying occasional or ancient dispersal are predicted to prevail (Whittaker 1998; Fattorini 2009; Dapporto and Dennis 2009). In the first case, island populations can be considered as part of metapopulations dominated by a mainland habitat patch, in which continuous gene flow probably occurs (see Dennis and Shreeve 1996, 1997; Dennis 2000; Dapporto and Dennis 2008a for island butterflies). As with patches in mainland metapopulations, extinction on islands is predicted but the extinction of any population is readily replaced by new propagules from the mainland (i.e., rescue effect) (Hanski 1999). Conversely, when island populations are established after exceptional colonisation events, they are unlikely to be re-established after extinction. Moreover, when gene flow is reduced, the probability of an island population diverging to produce an endemic taxon is increased (Williamson 1981; Whittaker 1998; Dapporto and Dennis 2009). These observations suggest that there are two main reasons to protect species that have colonized islands after exceptional events or persisted there as relict populations: (i) they are unlikely to re-colonize the island following extinction and (ii) they may represent endemic (thus unique and irreplaceable) populations.
In a recent paper such questions have been tested in west Mediterranean butterflies belonging to the superfamily Papilionoidea (Dapporto and Dennis 2009). The islands of Sardinia and Corsica had substantially impoverished butterfly faunas. Predicted absence or occurrence of each species on islands were achieved by modelling incidences from mainland distributions (Dennis 2000; Dapporto 2009; Dapporto and Dennis 2008a, 2009; Dapporto et al. 2009). The missing species were shown to have distinct ecological traits compared to species found on the islands (i.e., lower occurrence at mainland, lower altitudinal tolerance, shorter flight period) (Dapporto 2009; Dapporto and Dennis 2008a, 2009). The importance of these characteristics in explaining butterfly distributions, is in pointing to deterministic factors underlying impoverishment. Yet, some island species had the same ecological traits as missing species, suggesting ancient or stochastic colonization. Intriguingly, most of these species represent island endemics (Dapporto and Dennis 2009). On the other hand, butterflies on Sicily did not have the same impoverishment pattern and this would suggest that the narrow Messina Channel (3 km wide) fails to generate impoverishment (Dapporto and Dennis 2009).
In this paper we have applied a similar analysis to the Hesperioidea (skippers). Hesperioidea differ from Papilionoidea in many respects; mainly in their small size, high wing loading (the ratio of wing area to body mass), thus the particular kind of flight, and their narrower biotope requirements (meadow association). These differences are expected to confer a lower colonisation capability to skippers compared to butterflies. Moreover, owing to their inconspicuousness, these Lepidoptera are often neglected in conservation projects. Thus, identifying the factors that determine the occurrence and impoverishment of skippers on islands, and comparing the results with those obtained on Papilionoidea, may usefully identify the deterministic, historical and stochastic processes underlying island impoverishment as well as potentially endangered species that deserve to be conserved.
Materials and methods
Study area and geological history
Sardinia (Sa), Sicily (Si) and Corsica (Co) are the largest islands in the West Mediterranean Area (WMA). Together, Sardinia, and Corsica have a very different geological history and geographical position than Sicily. The former islands were separated from the West European continent (Spain and southern France) in the Miocene and have remained isolated from the mainland during the last 3–5 MY (Meulenkamp and Sissingh 2003). Sardinia and Corsica, separated by 11 km of sea, but 80 km from the mainland, are closely neighbouring but isolated landmasses. In contrast, Sicily is part of the African plate but is separated from the Italian Peninsula by only a 3 km strait.
Data sources
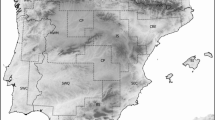
Presence and absence data on butterflies (Superfamily Hesperioidea) were compiled for the WMA and the three islands (Corsica, Sardinia and Sicily) (Appendix 1, Fig. 1) from a number of recent sources (see below). As the records are the product of long term intensive survey and refer to broad areas, they can be considered an accurate data source for the study (i.e. the absence of a species from a region is unlikely to be due to sampling inaccuracies). The data are compiled for (i) Italian regions including the Italian Peninsula and the south-eastern part of Alps representing the eastern WMA (Val D’Aosta, Va; Lombardia, Lo; Trentino Alto Adige, Ta; Veneto, Ve; Friuli Venezia Giulia, Fv; Piemonte, Pi; Liguria, Li, Emilia Romagna, Er; Toscana, To; Marche, Ma, Umbria, Um; Lazio, La, Abruzzo, Ab; Molise, Mo; Campania, Ca; Puglia, Pu; Basilicata, Ba; Calabria, Cl); (ii) Central-Mediterranean France representing the northern part of WMA (Auvergne, Au; Rhone Alpes, Ra; Provence-Alpes-Cote d’Azur, Pa; Midi Pyrenees, Mp; Languedoc-Roussillon, Lr); (iii) Spain representing the western WMA (Catalunya, Ct; Aragon, Ar; Comunidad Valenciana, Cv; Murcia, Mu, Castilla la Mancha, Cm; Extremadura, Ex; Andalusia, An); and (iv) Mediterranean Africa (from the southern slopes of Atlas until Mediterranean coast of Morocco, Algeria and Tunisia) representing the southern WMA. For Northern Africa detailed locational data are unavailable. Therefore, Mediterranean Morocco has been divided into two regions, the first comprising the regions of the Mediterranean coast (Mn) and the second the regions of High and Middle Atlas (Ms). The Ms area is particularly important in regression analyses as it represents a region south of Sicily with comparable high altitudes. We divided Mediterranean Algeria into two parts, West Algeria (Aw) and East Algeria (Ae). For continental Italy, data were obtained from Balletto et al. (2005) and Parenzan and Porcelli (2006), for continental France from Lafranchis (2000) and for Spain from García-Barros et al. (2004). For North Africa, data were extracted from distribution maps in Higgins and Riley (1983) and Tolman and Lewington (1997). For the large islands there is discordance for some old records. For this reason, data for Corsica (Co), Sardinia (Sa) and Sicily (Si) were compiled with regard to maximum concordance for the different sources (Lafranchis (2000), Kudrna (2002), Brusseaux and Nel (2004), Balletto et al. (2005), Parenzan and Porcelli (2006)) and confirmed by unpublished records collected by the first author (Appendix 1).

West Mediterranean study regions and islands. Grey scale illustrates butterfly species richness. See Methods for regional abbreviations
Environmental data and species’ designation
The following geographical characteristics were measured for each region and island: area, maximum altitude, minimum distance from sea, median latitude (between maximum and minimum latitude) (Lt) and longitude (Lg), latitudinal and longitudinal range (calculated as the maximum minus the minimum value, LtE, LgE, respectively). As in Dapporto and Dennis (2009) the main focus of this paper is impoverishment; the absence of a species on islands is not considered a “true absence” when it occurs in the guise of a sister species. Therefore, we have amalgamated data for well recognised endemics with their sister species and for several recently separated taxa in Europe following recent reviews (Tolman and Lewington 1997; Dennis 1997; Kudrna 2002). In particular the Pyrgus malvae complex includes P. malvae (Linnaeus, 1758) and P. malvoides (Elwes & Edwards, 1897); Pyrgus alveus complex includes P. alveus (Hübner, 1803) and P. centralitaliae (Verity, 1920); Pyrgus foulquieri complex includes P. foulquieri (Oberthür, 1910) and P. picenus (Verity, 1920); Spialia sertorius complex includes S. sertorius (Hoffmannsegg, 1804), S. therapne (Rambur, 1832), S. orbifer (Hübner, 1823) and S. ali (Oberthür, 1881); Carcharodus alceae complex includes C. alceae (Esper, 1780) and C. tripolinus (Verity, 1925); Carcharodus baeticus complex includes C. baeticus (Rambur, 1840) and C. stauderi Reverdin, 1913.
The range size of any species’ occurrence at continental sources is taken to be the number of 30 × 30 km2 squares occupied in Europe and the distribution in coastal areas is the number of coastline 30 × 30 km2 squares occupied by each species. These data are abstracted for Europe from Kudrna (2002). Data are missing only for North Africa. The length of the flight period, voltinism (number of generations), maximum altitude, and minimum altitude at which each species are recorded in Europe have been taken from Higgins and Riley (1983) and Tolman and Lewington (1997). The number of known host-plants has been abstracted from 20 different sources and is applied as a measure of larval resource generalism and colonisation capacity (Dennis, unpublished data). As suggested by McPherson and Jetz (2007), we refer to these characteristics as “ecological traits” even if some are intrinsic to the species while others derive from interactions with the environment.
Statistical analyses
Impoverishment was first assessed for species richness. As species richness usually depends on area, we first verified that the areas for the three islands do not differ from those of the 35 potential source regions (Mann–Whitney U = 42.0, P = 0.620). Subsequently, stepwise multiple regression analysis was applied using log-transformed regional species’ richness (referred to as region-focused analysis) as the dependent variable and with log-transformed geographical characteristics as predictors: area, maximum altitude, minimum distance from sea, latitude, longitude, latitude and longitude ranges. Following the construction of the regression model, the three islands (Sicily, Sardinia, and Corsica) were included and their theoretical number of species predicted. Species’ richness residuals for Sicily, Sardinia, and Corsica determine the extent to which their faunas are impoverished compared to three virtual continental areas having the same geographical characteristics of the three islands.
Thereafter, a three step analysis was carried out to investigate the nature of insular impoverishment.
Step 1: Predicting species’ incidence for islands
The probability of individual species occurring on islands was determined using logistic regression analyses (species-focused analysis). Species’ presence or absence for each region was entered as the dependent variable and log transformed geographical regional characteristics (maximum altitude, minimum distance from sea, latitude and longitude) as predictors. Species absent in less than five regions (for which logistic regression was unreliable) were classified as ubiquitous and regarded as being present in all three islands; similarly, species present in less than five regions were classified as rare species and considered to be absent from the islands (Dapporto and Dennis 2009). Once the regression models were established, the three islands (Sicily, Sardinia, and Corsica) were included and the theoretical occurrence of each species predicted. This procedure ensured a classification of species into those correctly and incorrectly predicted to be present or absent on islands. The sum of those predicted to be present from the species-focussed analysis (SPP) could then be compared with the number of species predicted to be present from the region-focussed analysis for species richness.
Step 2: Determining which species are missing
A list of missing species describing impoverishment (i.e. SPPA) on islands is obtained by verifying which species are actually present (SPPP) and which are absent (SPPA) among those predicted to be present (SPP).
Step 3: Determining the factors underlying species’ occurrence on islands
Unexpected absences of species at a regional scale (SPPA) are explained by species’ ecological traits underlying migration and colonisation capacity (Dennis et al. 2000). Previously, logistic regression analysis on Papilionoidea was used to compare SPPP and SPPA against ecological trait variables (Dapporto and Dennis 2009). This approach is not possible here owing to the low number of SPPP for Sardinia and Corsica. For this reason we compared ecological traits (wingspan, length of flying period, voltinism, the maximum altitude, minimum altitude, number of known host-plants, number of occupied coastal dots and range size) for SPPP and SPPA using the exact P values from a Mann–Whitney non-parametric test. Species present, and predicted to be present (SPPP), but forming an outgroup and falling in the opposite quartile for ecological traits along with SPPA are considered as deserving to be conserved because they are predicted not to have the ecological traits allowing them to occur on the particular island (Dapporto and Dennis 2009).
Results
Assessment of overall impoverishment
Regional species’ richness in the West Mediterranean is largely accounted for by increases in latitude (stepwise multiple regression model: F = 26.86, R 2 = 0.627, P < 0.001; latitude: Beta = 0.572, R 2 = 0.481, P < 0.001) and area (Beta = 0.401, R 2 = 0.146, P = 0.001). A plot of observed versus predicted numbers of species confirms high predictability for regional species’ richness in the continental West Mediterranean with the exception of Tunisia which has fewer species than expected. When islands are included in the model, large negative residuals for Sardinia (observed S = 5, predicted S = 16.8 ± 1.5, residual = −13.8) and Corsica (observed S = 6, predicted S = 20.1 ± 1.3, residual = −14.1 species) emerge (Fig. 2). On the other hand Sicily is less impoverished but has fewer species than estimated from the lower confidence limit of the predicted value (observed S = 14, predicted S = 18.0 ± 2.5, residual = −4.0) (Figs. 1, 2).

Observed versus predicted values of species’ richness (logs) based on multiple regression analysis. See Methods for regional abbreviations
Step 1: Predicting species’ occurrences on islands
Among the 35 Hesperioidea species examined here, 4 have limited incidences and occur within one to four regions; they are predicted to be absent from the three islands. On the other hand, 9 are present in virtually all regions and are thus predicted to be present on all three islands (Appendix S1). For the remaining species, logistic regression analyses disclosed the significant influence of at least one geographical predictor. Overall, occurrence data are explained by latitude (20 species), longitude (7 species), maximum altitude (2 species) and minimum distance to the sea (1 species) (Appendix S1). The primacy of latitude conforms with the results of the stepwise multiple regression analysis on species’ richness. Altogether, the individual species’ logistic regression analyses predicted the occurrence of 18 species in Sicily, 20 in Sardinia, and 22 in Corsica. These numbers closely match predicted richness independently obtained by multiple regression.
Step 2: Determining which species are missing
Results of the logistic regressions indicate that all species observed on the three islands are actually predicted to occur with one exception, Pyrgus armoricanus (Oberthür, 1910), which is predicted to be absent in Sicily when in fact present. This result together with the high concordance of Spp numbers with predicted island richness strongly points to SppA species accounting for impoverishment. A detailed list is provided in supplemental material (Appendix S1).
Step 3: Determining the factors of species’ occurrence on islands
Mann–Whitney comparisons of ecological traits and distributional data for species predicted to be present and actually present (SPPP) versus species predicted to be present but absent (SPPA) generate different results for the three islands. For Sicily SPPA species have a high minimal altitude than SPPP (Fig. 3c, Table 1); all SPPP are known to live at the sea level except for Carcharodus baeticus and C. flocciferus (Zeller, 1847), which are identified as outgroup cases. For Corsica, significant differences occur for wingspan and altitudinal range (Table 1, Fig. 3a, b). SPPP species are smaller than SPPA but no outgroups exist, nor do SPPP species have larger wingspans than the median for SPPA. SPPP have a higher altitudinal range than SPPA though P. armoricanus, a SPPP species, has a range lower than the median SPPA value. For Sardinia no significant effects emerged (Table 1).

Boxplot representations of median, interquartile and extreme values between SPPA and SPPP of wingspan and altitudinal range in Corsican species A, B and minimal altitude in Sicilian species C. Grey squares represent individual species, black squares represent outgroup species and SPPP species showing values comprised in the unpredicted 50% of SPPA
Discussion
With the exception of Tunisia, skipper richness is suitably modelled for West Mediterranean mainland regions on the basis of two independent variables (latitude and region area). Comparatively, Tunisia has a depauperate fauna. This may arise from the lack of high mountain chains where most species have refuges, as in Morocco and Algeria, and/or from possible sampling deficiencies. If the latter, several additional species may be added to the Tunisia fauna in future. When the three largest islands of West Mediterranean are included in the model, their expected richness is based on the premise that conditions match those of mainland regions. Observed species’ richness, however, is below the 95% confidence intervals in each case. As such, Sicily, Sardinia and Corsica have impoverished skipper faunas. This result is confirmed by an independent finding; for each island, species-focussed logistic regression analyses identified a matching number of predicted species determined by region-focused multiple regression analysis. The concordance in these results facilitates the identification of species underlying island impoverishment. This finding is reinforced by the fact that all species occurring on the three islands are actually predicted to occur on them with the sole exception of Pyrgus armoricanus in Sicily. Having established which species fall into SPPA and SPPP, it has been possible to identify ecological traits correlated with their occurrence probabilities. Differences occur between SPPA and SPPP for Sicily and Corsica, but for Sardinia none emerged. Below, the biogeographic and conservation implications are discussed.
Biogeography of West Mediterranean skippers
Mediterranean skippers differ substantially from butterflies (superfamily Papilionoidea) mainly in being smaller, with higher wing loading, and in performing short, darting, rapid flights (Higgins and Riley 1983). All Hesperioidea are associated with meadows where exists both flowers for adults and their herbaceous larval host plants (Higgins and Riley 1983). These characteristics may substantially influence their dispersal and colonisation capabilities. Several skipper species typically inhabit mountain areas, most of these confined to the Alps and Pyrenees. The majority of the remainder are tolerant of high altitude and can be found between sea level and 2000–2500 m a.s.l. (Appendix S1). Thus, as with the Papilionoidea, the Alps and Pyrenees form the richest regions for skippers (compare Fig. 1 in Dapporto and Dennis 2009). However, most of the mountain species are restricted to the marginal Mediterranean latitudes and are thus not predicted to occur on the three islands. Consequently, their absence has not contributed to island impoverishment. Among the remaining, widespread species there are some taxa more tolerant of low altitudes and others that have wider altitudinal ranges. Differences in overall altitudinal tolerance distinguish SPPP and SPPA species in both Sicily and Corsica. Mediterranean islands are usually warmer and drier than mainland regions located at the same latitudes and mountain areas are less prominent. Mountains on islands offer environmental diversity and refuge from dry summers, the reason why large numbers of species and endemics are concentrated on them, but, presenting an ecological island within a physical island, stochastic processes, including extinction, are predicted to occur frequently. An ability to persist at lower altitudes (as occurs on Sicily) or to exploit biotopes over a wide range of altitudes (as revealed on Corsica) may well increase survival and island incidence.
One counterintuitive finding is that SPPP species on Corsica are smaller than SPPA. Generally, it is expected that larger species disperse better, and colonize islands more frequently, than small species. Smaller butterflies tend to be associated with a stress-tolerant strategy and have lower dispersal capability (Dennis et al. 2004). However, dispersal by passive transport has been reported many times for butterflies and is more likely in small species (see Dantart et al. 2009 and literature therein). It may well be the case that Hesperioidea engage in less active long-dispersal than Papilionoidea and that their dispersal relates more to size and “occasional” wind driven events. It has been also hypothesised in the past that certain insect faunas of Tyrrhenian islands have been generated by propagules blown by prevailing North-West winds (Baroni Urbani 1974; Fattorini 2009). On the other hand, investigations of adult movement in Hesperia comma (Linnaeus, 1758) carried out on 998 individuals in England found that approximately 90% of individuals moved less than 100 m, and 67% moved less than 50 m among patches of suitable habitat (only one individual moved 1070 m) (Hill et al. 1996). A similar analysis for Erynnis tages (Linnaeus, 1758) disclosed that about 75% of recaptured specimens were found in patches where they had been marked and that only some 25% of movements were further than 100 m (Gutierrez et al. 1999).
In the more isolated Sardinia, where mountains rise less than 1850 m, no distinctions in ecological traits occur between SPPP and SPPA. This suggests either that some influential variables have been missed or that historical and/or stochastic effects determine Hesperioidea fauna on this island as much as on Sicily and Corsica. Such types of determinants in Hesperioidea occurrence are also suggested by the absence of any influence of range size and mainland coastal distribution. Instead, as for Papilionoidea (Dapporto 2009; Dapporto and Dennis 2008a, 2009) mainland source abundance is the main determinant for island species’ richness and occurrence, implicating ongoing dispersal as a key variable in determining island faunas at specific level.
A comparison of the butterfly faunas of the three islands provides a further indication of the lower dispersal capability of Hesperioidea compared to Papilionoidea. For Papilionoidea, Sicily is not significantly impoverished; the number of species present but expected to be absent is higher than the number of species that are absent but expected to occur. This suggests that many species from the Italian mainland have been are able to cross the Messina Channel and colonize suitable areas of Sicily. On the other hand, among Hesperioidea on Sicily, only one species P. armoricanus is present that was predicted to be absent, whereas five species predicted to be present are actually missing. This has facilitated the comparison between SPPP and SPPA that was not possible to carry out for Papilionoidea (Dapporto and Dennis 2009).
A final indication of a lower colonisation capability among skippers compared to Papilionoidea is evident in the ratios between SPPP and SPP. For Papilionoidea, on Sicily, Sardinia and Corsica, the ratios between species predicted to occur and species actually present are 1.06, 0.46 and 0.46, respectively (Dapporto and Dennis 2009). For skippers, these values are lower: 0.42, 0.20 and 0.23, respectively. A similar result was obtained by Dennis and Shreeve (1996 p. 60) by regressing island occurrence of butterfly on British offshore islands against their ecological traits. All the eight Hesperioidea showed incidence on islands lower than predicted, thus suggesting a lower colonisation ability compared to remnant butterflies. Similar reasoning can be applied to Zygaenidae moths. These Lepidoptera are characterized by very slow flight and consequently have very low dispersal capacity. About 50 species (more than Hesperioidea) occur on West Mediterranean mainland but only one endemic species (Zygaena corsica Boisduval, 1828) occurs in Corsica and two endemic species (Zygaena corsica and the North African Zygaena orana Duponchel, 1835) occur in Sardinia (Karsholt and Razowski 1996).
Overall, it would seem that in colonising islands, Hesperioidea depend more on historical or stochastic driven events than Papilionoidea. This may be owing to lower dispersal capability or to one or both of the following potential factors.
-
(i)
Skipper faunas may be less accurately recorded than Papilionoidea. Skippers are small and inconspicuous and may be missed during recording (Dennis et al. 2006). In fact, the record for Thymelicus lineola (Ochsenheimer 1808) on Sardinia was not accepted in recent revisions (Balletto et al., 2005 and Parenzan and Porcelli 2006). However, during 2009, the first author found dozens of individuals in Sardinia in the neighbourhood of Gennargentu Mountain. Similarly, there are single and old records for Pyrgus alveus (Hubner, 1803), P. carthami (Hübner, 1813), Carcharodus lavatherae (Esper, 1783) and Erynnis tages for Sicily and for Carcharodus flocciferus, C. baeticus and Thymelicus acteon (Rottemburg, 1775) from Sardinia which, lacking confirmation, induce recent assessments (Tolman and Lewington 1997; Kudrna 2002; Balletto et al. 2005), and this study, to exclude them from the islands’ fauna. Future studies may confirm their occurrence as for T. lineola in Sardinia, thus changing the frequencies for faunal impoverishment.
-
(ii)
The skippers considered here number 35 species against 284 Papilionoidea species examined by Dapporto and Dennis (2009). This discrepancy may create some type II errors (i.e. undetected existing phenomena) owing to low sample size, mostly in the SPPA vs SPPP Mann–Whitney U tests.
Conservation issues
Butterfly species occurring on Mediterranean islands should be always regarded as potential conservation targets (Dapporto and Dennis 2008b). In the West Mediterranean islands most Satyrinae have unexpectedly distinctive phenotypes implicating historical events (Dapporto 2010). Usually, migrant species, and those that can easily re-colonize an island from the mainland and/or other islands, are not considered to be a conservation priority (Dapporto and Dennis 2008b, 2009). On the contrary, species with populations that are unlikely to be re-established following extinction should be regarded as deserving conservation attention (Dapporto and Dennis 2009).
For this reason Pyrgus armoricanus, which unexpectedly occurs on Sicily, deserves to be conserved. The same may be said for Carcharodus flocciferus and C. baeticus from Sicily and P. armoricanus from Corsica. On these islands, they have low altitude ranges and are intolerant of sea level altitudes as is the case for most SPPA species. As such these species are atypical on the islands and probably vulnerable to extinction. When the 30 × 30 km mapped sampling points for these species are counted for Corsica and Sicily (in Kudrna 2002) it emerges that P. armoricanus has been recorded for just one on Corsica between 1980 and 2002 (cf., an average of 1.7 for the six SPPP species); in Sicily the mean number of occupied sampling points is 6 while P. armoricanus, C. baeticus and C. flocciferus were recorded for three, two and six, respectively. This suggests that these species are probably less common than the others or represent relict populations linked to particular biotopes with limiting resources. For Sardinia stochastic and/or historical immigration and establishment events seem to be the most likely explanation for species occurrence and, on the basis of our analyses, no species can be identified as deserving special treatment. Even so, if species colonized islands in the past when environmental and geographical settings were different from the contemporary ones or by means of stochastic events, re-colonization following possible extinction may be considered an unlikely event. On this basis, we suggest that all the Sardinian species, without any distinction, deserve to be conserved. Owing to the sparse distribution of Hesperioidea on all islands this reasoning can also be extended to Sicilian and Corsican species. Reference to Spialia species supports this view. In the species-focussed analyses the widespread, mainland Spialia sertorius was amalgamated with the Sardinian and Corsica endemic S. therapne and with the exceptional (probably relictual) eastern Mediterranean S. orbifer. The occurrence of pre-glacial and post-glacial relict populations seems to be the rule on Mediterranean islands (Habel et al. 2009, 2010; Dapporto 2010; Dapporto et al. 2009). Clearly if native island Spialia populations are lost, they cannot be replaced by propagules with the same genotypes from the neighbouring mainland.
In conclusion, Hesperioidea impoverishment on Sicily, Sardinia and Corsica discloses the action of deterministic, historical and stochastic factors. Overall impoverishment and missing species is easily evaluated by comparison with species distributions on the nearby mainland. A comparison with a previous study on the other butterfly superfamily (Papilionoidea) indicates that Hesperioidea island faunas are more impoverished; this suggests that they have a lower dispersal capability and are reliant to a greater degree on historical or exceptional (stochastic) dispersal events for island colonization. In reviewing conservation priorities, our main conclusion is that the extinction of Hesperioidea species on islands should be considered as irreparable events, and that these butterflies, often neglected owing to their inconspicuousness, are worthy of conservation programmes. These results, together with their implications, have been obtained by minimizing subjective evaluations and by strictly adhering to biogeographic analyses, as advocated by conservation biogeography (Whittaker et al. 2005).
References
Balletto E, Monelli S, Cassulo L (2005) Checklist e distribuzione della fauna italiana. 10.000 specie terrestri e delle acque interne: Insecta Lepidoptera Papilionoidea (Rhopalocera). Mem Museo civico Storia Nat Verona 16:259–263+CD Rom
Baroni Urbani C (1974) Studi sulla mirmecofauna d’Italia. Le Isole Pontine. Fragmenta Entomol 9:225–252
Brusseaux G, Nel J (2004) Révision de la liste-inventaire de Charles E.E. Rungs (1988) des Lépidoptères de Corse. Revue de l’Association Rousillonnaise d’Entomologie 13:1–145
Dantart J, Stefanescu C, Avila A et al (2009) Long-distance wind-borne dispersal of the moth Cornifrons ulceratalis (Lepidoptera: Crambidae: Evergestinae) into the northern Mediterranean. Eur J Entomol 106:225–229
Dapporto L (2009) Core and satellite butterfly species on Elba island (Tuscan Archipelago, Italy). A study on persistence based on 120 years of collection data. J Insect Conserv 13:421–428
Dapporto L (2010) Satyrinae butterflies from Sardinia and Corsica show a kaleidoscopic intraspecific biogeography (Lepidoptera, Nymphlidae). Biol J Linn Soc 100:195–212
Dapporto L, Dennis RLH (2008a) Species’ richness, rarity and endemicity of Italian offshore islands: complementary signals from island-focused and species-focused analyses. J Biogeogr 35:664–674
Dapporto L, Dennis RLH (2008b) Island size is not the only consideration. Ranking priorities for the conservation of butterflies on Italian offshore islands. J Insect Cons 12:237–249
Dapporto L, Dennis RLH (2009) Conservation biogeography of large Mediterranean islands. Butterfly impoverishment, conservation priorities and inferences for an ecological “island paradigm”. Ecography 32:169–179
Dapporto L, Bruschini C, Baracchi D, Cini A, Gayubo SF, Gonzàlez JA, Dennis RLH (2009) Phylogeography and counter-intuitive inferences in island biogeography: evidence from morphometric markers in the mobile butterfly Maniola jurtina (Linnaeus) (Lepidoptera, Nymphalidae). Biol J Linn Soc 98:677–692
Dennis RLH (1997) An inflated conservation load for European butterflies: increases in rarity and endemism accompany increases in species richness. J Insect Cons 1:43–63
Dennis RLH (2000) The comparative influence of source population size and migration capacity on the persistence of butterfly species on a small offshore island. Ent Gaz 51:39–52
Dennis RLH, Shreeve TG (1996) Butterfies on British and Irish Offshore Islands. Gem Publishing Company, Wallingford
Dennis RLH, Shreeve TG (1997) Diversity of butterfly species on British islands: ecological influences underlying the roles of area, isolation and faunal source. Biol J Linn Soc 60:257–275
Dennis RLH, Shreeve TG, Olivier A (2000) Contemporary geography dominates butterfly diversity gradients within the Aegean archipelago (Lepidoptera: Papilionoidea, Hesperioidea). J Biogeog 27:1365–1384
Dennis RLH, Hodgson JG, Grenyer R et al (2004) Host plants and butterfly biology. Do host plant strategies drive butterfly status? Ecol Entomol 29:12–26
Dennis RLH, Shreeve TG, Isaac NJB et al (2006) The effects of visual apparency on bias in butterfly recording and monitoring. Biol Conserv 128:486–492
Fattorini S (2009) Both Recent and Pleistocene geography determine animal distributional patterns in the Tuscan Archipelago. J Zool 277:291–301
García-Barros E, Munguira ML, Martín Cano L et al (2004) Atlas de las mariposas diurnas de la Península Ibérica e islas Baleares (Lepidoptera: Papilionoidea and Hesperioidea). Monografias S.E.A. 11:1–228
Gutierrez D, Thomas CD, Leon-Cortes JL (1999) Dispersal, distribution, patch network and metapopulation dynamics of the dingy skipper butterfly (Erynnis tages). Oecologia 121:506–517
Habel JC, Dieker P, Schmitt T (2009) Biogeographical connections between the Maghreb and the Mediterranean peninsulas of southern Europe. Biol J Linn Soc 98:693–703
Habel JC, Rödder D, Scalercio S, Meyer M, Schmitt T (2010) Strong genetic cohesiveness between Italy and North Africa in four butterfly species. Biol J Linn Soc 99:818–830
Hanski I (1999) Metapopulation ecology. Oxford University Press, Oxford
Higgins LG, Riley ND (1983) A field guide to the butterflies of Britain and Europe. Collins, London
Hill JK, Thomas CD, Lewis OT (1996) Effects of habitat patch size and isolation on dispersal by Hesperia comma butterflies: implications for metapopulation structure. J Anim Ecol 65:725–735
Karsholt O, Razowski J (1996) The Lepidoptera of Europe. A Distributional Checklist, Apollo Books, Stenstrup
Kudrna O (2002) The distribution atlas of European butterflies. Oedippus 20:1–343
Lafranchis T (2000) Les papillons de jour de France. Belgique et Luxembourg et leurs chenilles, Collection Parthénope, Mèze
McPherson JM, Jetz W (2007) Effects of species’ ecology on the accuracy of distribution models. Ecography 30:135–151
Meulenkamp JE, Sissingh W (2003) Tertiary palaeogeography and tectonostratigraphic evolution of the Northern and Southern Peri-Tethys platforms and the intermediate domains of the African-Eurasian convergent plate boundary zone. Palaeogeogr Palaeocl 196:209–228
Parenzan P, Porcelli F (2006) I Macrolepidotteri italiani. Fauna Lepidopterorum Italiae (Macrolepidoptera). Phytophaga 15:1–393
Tolman T, Lewington R (1997) Butterflies of Britain and Europe. Harper Collins, London
Whittaker RJ (1998) Island biogeography: ecology, evolution, and conservation. Oxford University Press, Oxford
Whittaker RJ, Araujo MB, Jepson P et al (2005) Conservation biogeography: assessment and prospect. Divers Distrib 11:3–23
Williamson MH (1981) Island populations. Oxford University Press, Oxford
Acknowledgements
Our grateful thanks to two anonymous referees for their useful suggestions.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Dapporto, L., Dennis, R.L.H. Skipper impoverishment on large West Mediterranean islands (Lepidoptera Hesperioidea): deterministic, historical and stochastic factors. Biodivers Conserv 19, 2637–2649 (2010). https://doi.org/10.1007/s10531-010-9863-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-010-9863-4