
Abstract
After abandonment, dry grassland (Festuco-Brometea) areas decline due to gradual overgrowing by woody species and the expansion of perennial tall grass species. Dry grassland vegetation was formed by extensive livestock grazing, thus grazing is considered one of the most natural methods for managing this type of vegetation. Six years after introducing low-intensity sheep and goat grazing in seven nature reserves in Prague (Czech Republic), the following impact of this management on dry grassland vegetation was observed: The cover of expansive woody species, particularly Ligustrum vulgare, and to a smaller extent Cornus sanguinea and Prunus spinosa declined. In addition, a significant, long-term declining trend of the expansive species Arrhenatherum elatius was also observed. Also the cover of Pimpinella saxifraga and Allium senescens declined significantly with regard to statistical evaluation. On the contrary, the cover of Achillea millefolium, Centaurea stoebe, Securigera varia, Elytrigia repens, Erysimum crepidifolium, Falcaria vulgaris, Fallopia convolvulus and Verbascum lychnitis increased. The cover of species characteristic of dry grasslands (Festuco-Brometea) increased significantly. No changes were observed in the number and cover of the Red List species. In addition, the presence of nitrophilous and ruderal species increased. Species diversity also significantly increased. From our findings we can conclude that managing dry grasslands with low-intensity grazing can help to keep dry grassland vegetation in good condition and conserve its plant diversity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the Czech Republic, dry grassland formed of Festuco-Brometea plant communities belongs to the strongly endangered types of vegetation with the largest presence of endangered species (see Moravec et al. 1995; Holub and Procházka 2000; Procházka 2001). In many European countries, this type of vegetation is receiving much attention in regard to conservation management (e.g. Zobel et al. 1996; Dzwonko and Loster 1998; Poschlod et al. 1998; Barbaro et al. 2001; Jäger and Mahn 2001; Münzbergová 2001; Willems 2001; Wallis De Vries et al. 2002; Bąba 2003, 2004; Richter et al. 2003; Baier and Tischew 2004; Bornkamm 2006).
In the Czech Republic, dry grassland was formed mainly as a secondary vegetation after clear-cutting of original thermophilous oak woods or oak-hornbeam woods and more rarely beech woods as well. A small area of dry grasslands, usually found on some rocks, rock edges or spots disturbed by erosion was formed as primary vegetation (Chytrý et al. 2001; see also Kolbek 1978).
Wild animal grazing that was later replaced by extensive grazing by goats, sheep, cattle and horses, was the main reason for the development of this vegetation and the establishment of its species richness (Kubíková 1999; Jäger and Mahn 2001; Dolek and Geyer 2002; Richter et al. 2003). Occasional or regular mowing of accessible spots or impact of fire caused positive influence on development of this vegetation type (Kubíková 1999; Kahmen et al. 2002). Existence of dry grassland thus depends, as in case of the other grassland plant communities, on human activities (Ložek 1971).
Apart from the above-mentioned disturbances, formation of plant communities is significantly influenced by stress caused by extreme characteristic habitat conditions, especially by low humidity, high temperature and low content of nutrients (Kubíková 1999).
Gradual changes in landscape farming throughout the 20th century, associated with intensification of farming, resulted in abandonment of relatively infertile habitats of dry grassland. This fact, together with increased atmospheric nutrient deposition (especially nitrogen—see e.g. Heil et al. 1988), result in reducing disturbances as well as stress factors. As a result, the number of tall, particularly perennial plants that competitively suppress plants of low growth, including terophytes and geophytes is increasing. In addition, this competitive effect is amplified by the accumulation of remaining old vegetation (Grime 1979) the presence of which mitigates extreme habitat conditions that determine the development and prosperity of conserved xerothermic vegetation. This process is regarded as one of the principal reasons for the reduction in species richness of dry calcareous grasslands (Willems 1983; Willems et al. 1993).
A key precondition of successful restoration or management of such vegetation is the necessity to renew disturbances using original or alternative farming methods (Zobel et al. 1996; Dzwonko and Loster 1998).
Grazing is considered one of the most natural methods of dry grassland management. Its impact results in removal of species of taller growth, with soft tissues (Hadar et al. 1999) that, in many cases, limit prosperity of conserved dry grassland communities. On the contrary, grazing supports the development of xerophilous species that contain strongly sclerotized tissues and species containing mechanical or chemical protective mechanisms (Ellenberg 1996). Trampling also contributes to the increase of heterogeneity of the environment, especially by creating gaps for the start of seedling growth (Bullock et al. 2001).
A number of habitats with dry grassland vegetation are being maintained in Prague. These currently have the status of nature reserves. Woody species, most frequently blackthorn (Prunus spinosa) gradually overtake these habitats and permanent expansive grass species, especially Arrhenatherum elatius become predominant.
To maintain the existing areas of xerothermic vegetation, workers of the Prague City Council who are in charge of the nature reserve maintenance, made the decision to remove undesirable woody species from the most valuable parts of the vegetation and introduce extensive grazing by goats and sheep in those areas to limit undesirable growth of woody species and expansive grass species and thus to maintain the remaining areas of still preserved or only slightly degraded xerothermic grassland.
The goal of this contribution is to evaluate to what extent extensive grazing can (1) prevent undesirable growth of woody species, (2) help maintain suitable species composition of xerothermic grassland and the occurrence and prosperity of endangered species and (3) influence the degree of occurrence of nitrophilous ruderal species.
Materials and methods
Study area
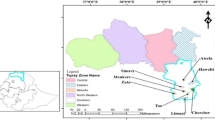
The study was carried out in seven nature reserves in Prague (Czech Republic; latitude 50° 05′ N, longitude 14° 25′ E). Figure 1 shows the more detailed location of the individual nature reserves. The individual reserves were established during 1968–1982. The studied nature reserves with xerothermic vegetation are located mainly on rocky hills along the Vltava River with an elevation of approx. 200–300 m a. s. l. The geological substratum in the Salabka, Podhoří, Podbabské skály, Baba and Jenerálka Nature Reserves (see Fig. 1) is mainly greywacke and shale of the Upper Proterozoicum on which a shallow layer of soil of protoranker type developed. In the Prokopské údolí and Homolka Nature Reserves, the geological substratum is mainly Silurian and Devonian limestone. The climate is of the mild humid zone type, with a pronounced but not too cold winter, but features of a transition to a mild arid zone type with hot summers and cold winters are also present (Walter and Lieth 1962). The mean annual temperature on the slope with dry grasslands is 9.0°C. The mean temperature for the period of April to September ranges from 15.0 to 15.7°C in the Prague Basin and from 14.2 to 14.6°C on the surroundings plateau. The mean annual total precipitation is 476–487 mm in the Prague Basin and 521–575 mm on the Prague Plateau. Climatic data represent the fifty years average and were processed using information from the Czech Hydrometeorologic Institute climatologic database. Earlier, grassland covering a considerable part of the hills was extensively grazed by domestic animals. When at the turn of the 1940s and 1950s extensive farming of these areas stopped, shrubs and trees began to invade and overgrow the grassland. At present, rich shrub vegetation is found in the nature reserves in which Prunus spinosa especially prevails. More advanced stages of succession, heading towards woodland communities dominated mainly by Quercus petraea, a potential natural vegetation on rendzinas and rankers, are visible on some spots. In Prague, these are especially xerothermic oak forests with Cynanchum vincetoxicum (Cynancho-Quercetum) and xerothermic downy oak forests (Lathyro vesicoloris-Quercetum pubescentis) (for details see Moravec et al. 1991). As a result of the above-mentioned advanced stages of succession, the overall area of dry grassland has become considerably smaller. In spite of this, the preserved dry grasslands that remain in these areas are relatively large. These dry grasslands are represented by the communities of alliances Festucion valesiacae and Alysso-Festucion pallentis. These species-rich communities are formed by character species such as Festuca valesiaca, F. rupicola, F. pallens, Koeleria macrantha, Carex humilis, Potentilla arenaria, Eryngium campestre, Thymus pannonicus, Centhaurea stoebe, Artemisia campestris, Seseli osseum, Sedum acre, and spring ephemerals such as Arenaria serpyllifolia, Erophilla verna or Cerastium umbellatum (for more details on this type of vegetation, see Kubíková 1976; Chytrý et al. 2001; Chytrý and Tichý 2003). Many of the stands that are degrading, especially as a result of the spreading of the expansive species Arrhenatherum elatius, however, can be classified only at the level of the class Festuco-Brometea.

Map showing position of studied nature reserves on the Prague territory (1—Podhoří, 2—Podbabské skály, 3—Salabka, 4—Baba, 5—Jenerálka, 6—Prokopské údolí, 7—Homolka)
Data collection
Data were collected in a similar way as used by Krahulec et al. (2001). To collect data, the system of permanent plots in the form of 1-m2squares was used. During the 2000–2006, we examined 70 plots in seven nature reserves (Salabka—6 plots, Podhoří—3 plots, Baba—7 plots, Podbabské skály—10 plots, Jenerálka—9 plots, Homolka—20 plots, Prokopské údolí—17 plots; see also Fig. 1). Plots were arranged in such a way to cover total vegetation heterogeneity (a mosaic of patches with cut shrubs, dominated by the expansive species Arrhenatherum elatius and in the good condition Festucion valesiacae communities). A 3 × 3 grid was used to divide each plot into 9 smaller subplots. In each subplot, the percentage of vegetation cover of all species and of each individual species was estimated in the middle of June, when the vegetation is found in its optimum stage of development. In this way, an estimate of cover for each species was obtained for the whole plot including mean value and standard deviation. In 2000, the state of vegetation cover was described prior to the start of extensive grazing. Three flocks of 25–35 sheep and 2–5 goats overgrazed the localities. The pasture season ranged from 1 to 4 weeks depending on the grazed area. Grazing intensity shown in Animal Days (AD) per unit area = stocking rate × days/area [ha] (see Hodgson 1979) was approximately 500 for sheep and 60 for goats. All seven nature reserves were treated the same way during the six-year study.
Four species groups that provide general information on the overall state of vegetation with regard to nature conservation and successfulness of grazing management were distinguished: woody species, nitrophilous and ruderal species, Red List species and Festuco-Brometea diagnostic species. Red List species are listed according to Procházka (2001). Diagnostic species of Festuco-Brometea are listed according to Chytrý and Tichý (2003). The publication of Chytrý and Tichý (2003) also includes Arrhenatherum elatius among the character species of the class Festuco-Brometea. In the communities of xerothermic steppes, however, this species behaves expansively and has a negative impact on the other species that are the subject of nature conservation (eg. Hroudová and Prach 1994; Baier and Tischew 2004). Controlling Arrhenatherum elatius was also one of the reasons for introducing grazing management. As a result, this species was not included in the group of character species of dry grassland vegetation.
Data analysis
Cover values of individual species within each of the subplots were averaged, so mean cover of every species per year and plot was obtained. The Shannon-Wiener index of species diversity (Pielou 1966) was calculated for every plot and year. The number and total cover of woody species, nitrophilous and ruderal species, Red List species and diagnostic species of the class Festuco-Brometea were calculated for each plot and year.
PD (percentage dissimilarity) was chosen as a coefficient of between-year dissimilarity. The dissimilarity values of PD conform well to the mental scaling of dissimilarities originating from ecologists’ field observations (Gauch 1982). For every plot percentage dissimilarity between years j and k was quantified as:
The summations are over all species i = 1−n and A ij and A ik are the abundances (cover) of species i in year j and k.
Percentage trend PT was calculated as a measure of the ratio of trend and oscillations in the cover changes in the period of m years of every plot as
The data of species cover, diversity, PD and PT were analyzed using SOLO (BMDP) statistical software. ANOVA for normal distributed data and Kruskall–Wallis test for other data were used to test the differences between the years and plots. Duncan’s multiple range test and Connover test were applied to compare the individual means.
Cannonical Correspondence Analysis (CCA; program CANOCO; ter Braak and Šmilauer 1998) with TIME as an environmental variable and plots as covariables was applied to examine the dependence of the cover of species or their groups on time. Significance of the effect of the environment variable was evaluated using the Monte Carlo test.
Results
Rate of changes in communities
Species turnover and rate of change in communities were characterized by mean percentage dissimilarities and trends in the species cover in the plots with reference to the first year of observation and to the preceding year (Table 1).
Percentage dissimilarity with reference to the first year continually increased till 2004. No significant increase was observed in the following years. Changes in communities occurred during the first four years after the start of grazing and then the communities were more stable.
Percentage dissimilarity with reference to the preceding year was surprisingly highest not after the beginning of grazing, but in the years 2003–2004 and 2004–2005. The rate was probably effected by a significant drought in 2003 that could have caused more significant changes in vegetation in the following two years.
The trend values show if in the course of 2000–2006 these changes in vegetation had, a certain direction and to what extent. The mean values of trend continually decreased.
Among the individual nature reserves, however, there are certain differences in the dynamics of changes of the composition of plant communities that are reflected by different values of percentage dissimilarity and trend (Table 2). Vegetation dissimilarity between the starting year (i.e., 2000) and ending year (i.e., 2006) of monitoring was highest in the Podbabské skály Nature Reserve, and the lowest in the Salabka, Podhoří, Jenerálka and Homolka Nature Reserves. The highest trend values were found in the Prokopské údolí Nature Reserve, and the lowest in the Baba, Podbabské skály, Podhoří, Jenerálka and Homolka Nature Reserves.
Species diversity
Table 3 shows that diversity values mostly increased throughout the monitoring period. A significant increase in the diversity value was observed in the first year after the start of grazing. In 2000–2003, values of species diversity were evenly and slightly increasing. In the next period of monitoring (2004–2006), however, stagnation in the growth of species diversity was observed.
Despite that total values of species diversity show steady growth in the period of monitoring, certain differences are visible between individual nature reserves (see the Table 4). The number of species increased significantly only in Podbabské skály and Baba Nature Reserves. Generally, it can be said that species diversity values, over the course of a six-year monitoring period, increased with statistical significance in two nature reserves (Prokopské údolí and Podbabské skály), in the other nature reserves, no statistically significant changes were observed. None of the monitored areas showed a decline in species diversity values or a drop in the number of species in the monitored period.
Changes in vegetation characteristics
Differences in the presence of plant species groups that provide summary information about the overall state of vegetation are listed in Table 3 and described in Fig. 2. Results show that throughout 2000–2006, total species cover of the Czech red listed vascular plants that are subject to nature conservation essentially did not change. The cover of Festuco-Brometea species was varying, and for that reason it is close to the vertical axis in Fig. 2; in 2006, however, it was statistically higher than in 2000. As a result of grazing, cover of woody species declined in a desirable way that is shown in Fig. 2 by the location that is found relatively far from the vertical axis in the direction opposite to the direction of time vector. Decline in cover, however, was not statistically significant. On the contrary, presence of nitrophilous and ruderal species (a statistically significant increase in number and cover—see Table 3, location in the direction of time vector in Fig. 2) increased. Also the cover of all species in the monitored areas has increased (Fig. 2).

A result of direct gradient analysis (CCA) of all studied areas shows differences in the presence of cover of species groups indicating an overall state of vegetation from 2000 to 2006. Groups of species in the left part of the graph have increased, groups of species in the right part of the graph have declined. The effect of year explained 7.6% of the cover variability of species groups on all localities. The test of the effect of year was significant (P = 0.005)
However, some differences between the individual nature reserves are visible in numerical values expressing the presence of individual groups of species. These differences however, apart from the Podbabské skály Nature Reserve where, with exception of woody species, a significant increase in the number and cover of all monitored species was observed, are mostly not significant with regard to statistical evaluation (see the Table 4). The number of species also did not significantly increase at any of the nature reserves.
Assessment of the impact of grazing on the changes in cover of individual plant species is included in Table 3 and Fig. 3. Monitoring in 2006 confirmed a significant long-term declining trend in the case of Arrhenatherum elatius. Results of statistical evaluation (Table 3) and direct gradient analysis (Fig. 3) show that a decline in the growth of woody species, especially Ligustrum vulgare, and to a smaller, non-significant extent, Cornus sanguinea and Prunus spinosa was also observed. From 2000 to 2006 a statistically significant decline in the cover of Pimpinella saxifraga and Allium senescens was also observed. In the case of Allium senescens, however, the changes were rather oscillations. On the contrary, a certain significant trend in the increase of cover was registered for the following species: Achillea millefolium, Centaurea stoebe, Securigera varia, Elytrigia repens, Erysimum crepidifolium, Falcaria vulgaris, Fallopia convolvulus and Verbascum lychnitis. On the areas where a decline in the cover of Arrhenatherum elatius was registered, the cover of the species Fallopia convolvulus increased.

A result of direct gradient analysis (CCA) of all studied areas shows the species with a significant growth or decline from 2000 to 2006. The species in the right part of the graph have increased, the species in the left part of the graph have declined. Effect of year explained 1.9% of the species cover variability in all localities. The test of the effect of year was significant (P = 0.005)
Results of direct gradient analysis—CCA (Fig. 3) confirm the results of statistical evaluation included in Table 3. Some species that in the resulting ordination diagram are found relatively far from the vertical axis occur only in one or few studied plots, thus the changes in their cover had no statistic significance in the overall evaluation.
Differences in the changes of cover of individual species between individual nature reserves are shown in Table 4. Compared with the other monitored nature reserves, a large decline of Arrhenatherum elatius in the localities Podbabské skály and Baba was registered. In these localities, Arrhenatherum elatius belonged to the predominant species so that impact of grazing on its cover could manifest more significantly. The most significant changes took place in the Podbabské skály Nature Reserve where the cover of seven species changed significantly with regard to statistical evaluation. The cover of Achillea millefolium, Centaurea stoebe, Securigera varia, Dianthus carthusianorum, Erysimum crepidifolium and Verbascum lychnitis has increased. Relatively prominent changes were also observed in the Homolka Nature Reserve where the cover of Securigera varia, Potentilla reptans and Scabiosa ochroleuca has increased. In the localities Baba, Jenerálka and Prokopské údolí, significant changes were observed only in the case of 1–3 species. In the Salabka and Podhoří Nature Reserves, no significant changes were observed for any of the monitored species.
Discussion
This study has shown that extensive grazing by sheep and goats is sufficient to control spontaneous succession after abandonment of dry grassland. After introducing this management method, cover of woody species, especially Ligustrum vulgare, and to a smaller extent, Cornus sanguinea and Prunus spinosa decreased. Barbaro et al. (2001) also observed a decline in the cover of woody species after the introduction of grazing. A long-term trend of decline in the cover of the expansively spreading Arrhenatherum elatius, that is sensitive to grazing, was a positive finding with regard to pasture management as was also reported by Poissonet et al. (1981) and Hewett (1985). This phenomenon is caused probably by a combination of two factors, grazing and treading. Grazing similar to vegetation mowing as was found by Wilson and Clark (2001) can cause decline in the presence of this species. However, habitat treading also has a considerable impact, adversely influencing the prosperity of this species.
Our observations also showed that no significant changes had occurred in the number and cover of the Red List species. An increase in the presence of character species of dry grassland (Festuco-Brometea) was even observed. Our findings thus confirm the conclusions of the study of Klimek et al. (2007) who found that grazing at a low stocking rate can support biodiversity conservation. Poschlod et al. (2002), Riecken et al. (1998), Wallis De Vries (1999), and Vera (2000) also pointed out the importance of low-intensity grazing for biodiversity conservation.
Krahulec et al. (2001) point out that the main problem associated with sheep grazing is the increasing presence of nitrophilous tall herbs and grasses. In our observation, under a low-grazing intensity, the presence of nitrophilous and ruderal species significantly increased (especially in the first year after the introduction of grazing), however, it did not exceed a tolerable level.
The total number of species and values of species diversity increased, which may also be attributable to the cutting of woody species before the introduction of grazing as is documented in the publications of Bąba (2003), and Dzwonko and Loster (1998). In our experiment, however, the most important factor causing these changes was grazing as can be seen by the significant increase in nitrophilous and ruderal species in the first year after its introduction.
Species diversity in general increased between 2000 and 2006. A significant increase in the diversity value occurred especially in the first year after the introduction of grazing. Between 2004 and 2006, however, stagnation was registered. An increase in species richness of the monitored vegetation was caused especially by the increase in the number of synanthropic species, mainly annual weedy species, whose development is associated with a disturbance of the soil substratum. The occurrence rate of these species, however, does not currently have an adverse impact on the communities of xerothermic vegetation that are the subject of nature conservation. On the contrary, a reasonably disturbed vegetation of xerothermic vegetation can become a refuge of rare segetal (weedy) species.
Two reasons for the decreasing trend values that show the extent of total vegetation changes are the following:
-
1)
The effect of grazing on communities is highest in the first years
-
2)
Between-year oscillations are cumulated, so the longer-term to short-term change ratio decreased with the duration of observation.
The rate of changes in communities indicates certain differences between individual nature reserves. Mean values of percentage dissimilarity and trend can be used to generally divide the nature reserves into four groups depending on the extent of changes of communities during 2000–2006 and the ratio of short-term (oscillations) and longer-term (trend) changes:
-
1)
Small changes with relatively small ratio of oscillations were found in the Salabka Nature Reserve. In this locality, succession played a large role in the small vegetation changes indicated.
-
2)
Small changes with large ratio of oscillations were found in the Podhoří, Jenerálka and Homolka Nature Reserves. These localities, characterized by a very shallow soil layer, were affected only little by grazing, but cover of some species showed between-year oscillations.
-
3)
Large changes with relatively small ratio of oscillations were found in the Prokopské údolí Nature Reserve. In this locality the impact of grazing was relatively big. Changes in communities were not only oscillations but some were of longer-term importance.
-
4)
Large changes with large ratio of oscillations were found in the Podbabské skály and Baba Nature Reserves. The soils of these localities are less stabilized, relatively loose, and were significantly synanthropically influenced in the previous period. Vegetation in these habitats is probably more responsive to disturbances of various types. The most important change in these localities is the considerable decrease of cover of Arrhenatherum elatius.
In general, it can be stated that conserved vegetation on the managed areas is in a relatively good condition. It is obvious that low-intensity grazing management in the monitored areas has a positive impact with regard to nature conservation. However, during pasture management it is necessary to pay attention to the possibility of heightened erosion as a consequence of gathering of goats and sheep at exposed habitats, especially on steep slopes and in the vicinity of rock outcrops. According to our experience, it would be efficient to combine grazing with ongoing clearing of shrubs and grass remains. Barbaro et al. (2001) confirmed the favorable impact of a shrub-clearing and grazing combination.
References
Bąba W (2003) Changes in the structure and floristic composition of the limestone grasslands after cutting trees and shrubs and mowing. Acta Soc Bot Pol 72:61–69
Bąba W (2004) The species composition and dynamics in well-preserved and restored calcareous xerothermic grassland (South Poland). Biologia, Bratislava 59:447–456
Baier A, Tischew S (2004) Naturschutz-Management auf Xerothermrasenstandorten in Sachsen-Anhalt-Gefährdungsanalyse und Entwicklungsstrategien am Beispiel des Naturschutzgebietes“Lämmerberg und Vockenwinkel”. Hercynia N F 37:201–230
Barbaro L, Dutoit T, Cozic P (2001) A six-year experimental restoration of biodiversity by shrub-clearing and grazing in calcareous grassland of the French Prealps. Biodivers Conserv 10:119–135
Bornkamm R (2006) Fifty years vegetation development of a xerothermic calcareous grassland in Central Europe after heavy disturbance. Flora 201:249–267
Bullock JM, Franklin J, Stevenson MJ, Silvertown J, Coulson SJ, Gregory SJ, Tofts R (2001) A plant trait analysis of responses to grazing in a long-term experiment. J Appl Ecol 38:253–267
Chytrý M, Kučera T, Kočí T (eds) (2001) Katalog biotopů České republiky [Habitat Catalogue of the Czech Republic]. Agentura ochrany přírody a krajiny, Praha (in Czech, English summary)
Chytrý M, Tichý L (2003) Diagnostic, constant and dominant species of vegetation classes and alliences of the Czech Republic: a statistical revision. Folia Fac Sci Nat Univ Masaryk Brun 108:1–231
Dolek M, Geyer A (2002) Conserving biodiversity on calcareous grasslands in the Franconian Jura by grazing: a comprehensive approach. Biol Conserv 104:351–360
Dzwonko Z, Loster S (1998) Dynamics of species richness and composition in a limestone grassland restored after tree cutting. J Veg Sci 9:387–394
Ellenberg H (1996) Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht. 5. stark veränderte u. verbesserte Aufl. Ulmer, Stuttgart
Gauch HG (1982) Multivariate analysis in community ecology. Cambridge University Press, Cambridge
Grime JP (1979) Plant strategies and vegetation processes. J Wiley & Sons, Chichester
Hadar L, Noy-Meir I, Perevolotsky A (1999) The effect of shrub clearing and grazing on the composition of a Mediterranean plant community: functional groups versus species. J Veg Sci 10:673–682
Heil GW, Werger MJA, De Mol W, Van Dam D, Heijne B (1988) Capture of atmospheric ammonium by grassland canopies. Science 239:764–765
Hewett DG (1985) Grazing and mowing as management tools on dunes. Plant Ecol 62:441–447
Hodgson J (1979) Nomenclature and definitions in grazing studies. Grass Forage Sci 34:11–18
Holub J, Procházka F (2000) Red list of vascular plants of the Czech Republic—2000. Preslia 72:187–230
Hroudová Z, Prach K (1994) Dlouhodobé změny reliktního stepního porostu v Českém krasu [Long-term changes in relict steppe community in Bohemia Karst, Czech Republic]. Příroda 1:63–72 (in Czech with English abstract)
Jäger C, Mahn EG (2001) Die Halbtrockenrasen im Raum Questenberg (Südharz) in Beziehungen zu ihrer Nutzungsgeschichte. Hercynia N F 34:213–235
Kahmen S, Poschlod P, Schreiber KF (2002) Conservation management of calcareous grasslands. Changes in plant species composition and response of functional traits during 25 years. Biol Conserv 104:319–328
Klimek S, Richter gen. Kemmermann A, Hofmann M, Isselstein J (2007) Plant species richness and composition in managed grasslands: the relative importance of field management and environmental factors. Biol Conserv 134:559–570
Kolbek J (1978) Die Festucetalia valesiacae-Gesellschaften im Ostteil des Gebirges České středohoří (Böhmisches Mittelgebirge) 2. Synökologie, Sukzession, und syntaxonomische Ergänzungen. Folia Geobotanica et Phytotaxonomica 13:235–303
Krahulec F, Skálová H, Herben T, Hadincová V, Wildová R, Pecháčková S (2001) Vegetation changes following sheep grazing in abandoned mountain meadows. Appl Veg Sci 4:97–102
Kubát K, Hrouda L, Chrtek J jun., Kaplan Z, Kirschner J, Štěpánek J (eds) (2002) Klíč ke květeně České republiky [Key to the Flora of the Czech Republic]. Academia, Praha
Kubíková J (1976) Geobotanické vyhodnocení chráněných území na severovýchodě Prahy [Geobotanische Erschätzung von Naturschutzgebieten im nordöstlichen Teil von Prag]. Bohemia Centralis 5:61–105
Kubíková J (1999) Xerotermní trávníky až semixerotermní lemy [Xerothermic grasslands and subxerothermic margins]. In: Petříček V (ed) Péče o chráněná území I. [Nature Reserves Management I.]. Agentura ochrany přírody a krajiny ČR, Praha, pp 213–236 (in Czech with German summary)
Ložek V (1971) K otázce stepí ve střední Evropě [Weitere Anmerkungen zum Problem der mitteleuropäischen Steppen]. Zpr Čs Bot Spol 6:226–232 (in Czech with German summary)
Moravec J et al (1991) Přirozená vegetace území hlavního města Prahy a její rekonstrukční mapa [Natural Vegetation of the Territory of the Capital City Prague and its Reconstruction Map]. Academia, Praha (in Czech with English summary)
Moravec J et al (1995) Rostlinná společenstva České republiky a jejich ohrožení 2. ed. [Red List of Plant Communities of the Czech Republic and their Endangerment 2.ed.], Severočeskou Přírodou, příl.1, pp 1–206
Münzbergová Z (2001) Obnova druhově bohatých xerotermních trávníků na příkladu rezervací Stráně u splavu a Stráně u Chroustova [Restoration of species rich xerophilous grasslands: a case study of two nature reserves in Central Bohemia]. Příroda 19:101–121 (in Czech with English abstract)
Pielou EC (1966) The measurement of diversity in different types of biological collections. J Theor Biol 13:131–144
Poissonet J, Poissonet P, Thiault M (1981) Development of flora, vegetation and grazing value in experimental plots of a Quercus coccifera garrigue. Vegetatio 46:93–104
Poschlod P, Kiefer S, Trankle U, Fischer S, Bonn S (1998) Plant species richness in calcareous grasslands affected by dispersability in space and time. Appl Veg Sci 1:75–90
Poschlod P, Schneider-Jacoby M, Köstermeyer H, Hill BT, Beinlich B (2002) Does large-scale, multi-species pasturing maintain high biodiversity with rare and endangered species?—The Sava floodplain case study. In: Redecker B, Finc P, Härdtle W, Riecken U, Schröder E (eds) Pasture landscapes and nature conservation. Springer-Verlag, Berlin, Heidelberg, New York, pp 367–378
Procházka F (ed) (2001) Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000) [Black and Red List of Vascular Plants of the Czech Republic—2000]. Příroda 18:1–166 (in Czech with English summary)
Richter B, Partzsch M, Hensen I (2003) Vegetation, Kultur- und Nutzungsgeschichte der xerothermen Hügellandschaft bei Mücheln/Wettin (Sachsen-Anhalt). Hercynia N F 36:91–121
Riecken U, Finc P, Klein M, Schröder E (1998) Überlegungen zu alternativen Konzepten des Naturschutzes für den Erhalt und die Entwicklung von Offenlandbiotopen. Natur und Landschaft 73:261–270
ter Braak CJF, Šmilauer P (1998) CANOCO Reference Manual and Usery’s Guide to Canoco for Windows. Software for Canonical Community Ordination (Version 4). Centre of Biometry, Wageningen
Vera FWM (2000) Grazing ecology and forest history. CABI Publishing, Wallingford Oxon, UK
Wallis De Vries MF (1999) The dilemma facing nature conservation and the role of large herbivores. In: Gerken B, Görner M (eds) The development of European landscapes with large herbivores—history, models and perspectives. Natur und Kulturlandschaft 3:24–31
Wallis De Vries MF, Poschlod P, Willems JH (2002) Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol Conserv 104:265–273
Walter H, Lieth H (1960–1967) Klimadiagram-Weltatlas, Jena
Willems JH (1983) Species composition and above ground phytomass in chalk grassland with different management. Vegetatio 52:171–180
Willems JH (2001) Problems, approaches, and results in restoration of Dutch calcareous grassland during the last 30 years. Restor Ecol 9:147–154
Willems JH, Peet RK, Bik L (1993) Changes in chalk-grasslands structure and species richness resulting from selective nutrient additions. J Veg Sci 4:203–212
Wilson MV, Clark DL (2001) Controlling invasive Arrhenatherum elatius and promoting native prairie grasses through mowing. Appl Veg Sci 4:129–138
Zobel M, Suurkask M, Rosén E, Pärtel M (1996) The dynamics of species richness in an experimentally restored calcareous grassland. J Veg Sci 7:203–210
Acknowledgements
This study has been financially supported by the administration of the Prague City Council. Zuzana Münzbergová and Pavel Špryňar collected the data in nature reserves Homolka a Prokopské údolí during 2000–2004. Janice Forry kindly improved our English and anonymous referees contributed valuable suggestions to the manuscript. This study falls under research projects nos. MZP0002707301 and AVOZ60050516. Special thanks go to Petr Slavík, Prague City Council Administration for logistical support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dostálek, J., Frantík, T. Dry grassland plant diversity conservation using low-intensity sheep and goat grazing management: case study in Prague (Czech Republic). Biodivers Conserv 17, 1439–1454 (2008). https://doi.org/10.1007/s10531-008-9352-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-008-9352-1