
Abstract
Although farmers have managed west African parkland savanna systems for 1,000 of years, concerns have been raised about the sustainability of these agro-ecosystems due to human population growth, shortening of fallow periods, droughts, desertification and new orientations towards cash generation away from subsistence farming. We conducted a tree diversity survey in 16 villages from Burkina Faso, Mali, Niger and Senegal, recording total species composition for 300 quadrats (mainly 50 × 20 m2) that were randomly sampled from the main landuse categories of parklands of village fields (VF), bush fields (BF), sylvopastoral zone (SP) and forest reserves (FR). About 110 tree species were encountered, including 100 indigenous species. The results from balanced species accumulation curves (based on randomized subsampling of the same number of quadrats from each village) showed that the accumulated number of species was smallest in VF, largest in FR and almost equal in BF and SP, whereas classical (unbalanced) species accumulation curves yielded different results. Although there was a significant within-village reduction in species richness with increasing diameter class [averaging 8.5–13.1 more species in the smallest (<5 cm) compared to the largest diameter class (>80 cm)] for the different landuse categories, new species were also encountered in larger diameter classes (2.7–7.2 species). The evidence for tree regeneration problems (including problems in FR) suggest that farmer-managed tree regeneration should be further explored and that advances in domestication and marketing of indigenous tree species may be crucial to tree conservation in parkland systems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Farmers have managed west African savanna systems for 1,000 of years (White 1983; Boffa 1999; Kristensen and Balslev 2003). As an example of the anthropogenic modification of the landscapes of dry west Africa, research into one village in Burkina Faso revealed that farmers did not consider trees in their fields to be natural but an effect of active selection and protection during land clearing, although only two species out of the total 101 species had been planted (Augusseau et al. 2006). Although parkland savanna landscapes are often dominated by one particular economic species, they contain a selection of other useful woody species, whereas inedible species are usually confined to hills, rocky outcrops or protected areas (Maranz and Wiesman 2003).
Concerns have been raised about the conservation of tree species within these managed agro-ecosystems due to human population growth resulting in the shortening of fallow periods (e.g., Wezel and Boecker 1998; Augusseau et al. 2006), due to severe droughts and desertification (e.g., Kristensen and Balslev 2003), as a result of an orientation towards cash generation away from subsistence farming (e.g., Maranz and Wiesman 2003; Augusseau et al. 2006) and likely due to combined influences of human and climatic influences (e.g., Gonzalez 2001; Wezel and Lykke 2006).
Here we report results on tree diversity patterns in 16 villages from Burkina Faso, Mali, Niger and Senegal. The specific aim of the study was to investigate current patterns of tree diversity within distinctive patterns of landuse. Two main questions were investigated: (i) whether substantial differences in tree species richness existed between different types of landuse; and (ii) whether there was any indication for problems with regeneration of tree species. As the study was based on a complete inventory of tree species that currently occur, it was also possible to compare current species composition with historical species lists.
Material and methods
Study area
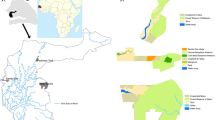
Tree diversity was characterised in 16 villages (Table 1, Fig. 1), including 14 villages that belonged to the benchmark sites of the Desert Margins Programme (DMP) in Burkina Faso, Niger and Senegal, and two villages in Mali belonging to the Tree Biodiversity Project (TBP). The DMP aims at arresting land degradation in Africa’s desert margins and addresses the loss of biodiversity, reduced sequestration of carbon, and increased soil erosion and sedimentation in these arid and semi-arid ecozones. Selected benchmark sites satisfied the criterion of having annual rainfall between 100 and 600 mm, which characterizes Sahelian drylands at risk of desertification (GEF 2002). Villages were randomly selected from the list of villages for each benchmark site, excluding those villages that had part of their territory outside the benchmark (Kalinganire et al. 2005). The TBP aims to improve the livelihoods of resource-poor farmers through the conservation of Sahelian traditional agroforestry systems, enrichment of agrobiodiversity and enhancement of the ecological functions of trees within these systems. The two selected villages had rainfall less than 600 mm and belonged to the cereal farming system.

Location of surveyed villages in Burkina Faso, Mali, Niger and Senegal with respect to the UNESCO/AETFAT/UNSO vegetation map of Africa (White 1983), country boundaries and geographical position [latitude and longitude (−15° E = 15° W)]
The vegetation type of villages (Table 1, Fig. 1) was determined from the UNESCO/AETFAT/UNSO vegetation map of Africa (White 1983, available in digital format from URL http://geodata.grid.unep.ch/). The Sudanian undifferentiated woodland of the majority of villages corresponds to the West Sudanian savanna terrestrial ecoregion of Africa (Burgess et al. 2004). Whereas White (1983) follows a strict definition to differentiate between woodland (cover percentage of at least 40%) and wooded grassland or savanna (10–40% cover), the description of the terrestrial ecoregion describes woodland (>40% cover) in the southern portion and grasslands (<10% cover) in the northern part. The West Sudanian terrestrial ecoregion includes all the Sudanian woodland vegetation types differentiated on the basis of the dominance of Isoberlinia (White 1983, Fig. 1). The Sahel wooded grassland and bushland corresponds to the southern part of the Sahelian Acacia savanna terrestrial ecoregion (Burgess et al. 2004, Fig. 1). The ecoregion also includes White’s northern Sahel semi-desert grassland and scrubland vegetation type, characterized by rainfall less than 250 mm per year (White 1983, Fig. 1). The current species composition of quadrats with the same original vegetation type was compared with the species composition of the vegetation types described in White (1983) and the listed habitats for the encountered species available from Arbonnier (2004).
Survey methodology
Rectangular quadrats for which all living trees (defined as woody perennial species) were counted and measured were established in all the survey villages, but survey methods differed between the countries. Surveyors in most countries used a quadrat size of 50 × 20 m2, with exceptions of quadrat sizes of 50 × 40 m2 in Niger and 25 × 20 m2 in FR in Senegal. The location of quadrats was established by transect surveys in the four cardinal compass directions from the village centre in Niger and during surveys outside FR in Senegal (locating quadrats at intervals of 500 m). Stratified-random sampling from available maps was the survey method for the other inventories. Village maps for stratified-random sampling were compiled by initial surveys with Global Positioning System receivers to obtain geographical coordinates for the boundaries of major landuse categories.
The landuse of a quadrat was categorised as village fields (VF; ‘champs de case’), bush fields (BF; ‘champs de brousse’), sylvopastoral (‘zone sylvopastorale’) and protected areas, which are the most common landuse categories of Sahelian parklands. VF surround the homesteads, are usually fertile and are continuously under cereal (mainly pearl millet or sorghum) or vegetable cultivation (Abegg et al. 2006). BF are located further away from the homesteads, are less fertile and larger than VF and accommodate cereal and cash crop (mainly cotton and tobacco) production (Kalinganire et al. 2005; Abegg et al. 2006). The sylvopastoral category consists of fallows which are left for grazing and harvesting of timber, fuelwood and other tree products (Kalinganire et al. 2005).
These three types of landuse categories form part of the parkland systems, which are farmed landscapes in which certain trees are scattered in cultivated or recently fallowed fields and livestock production is a significant component. Parklands are extremely dynamic systems which may develop over many generations, reflecting changes in the physical and socio-economic environment (Boffa 1999). To harmonize the definitions used in the different countries, the ‘pleins champs’ of Senegal (categorised in Senegal as a category of cultivated fields at distances from the village centre in between those of bush and VF) were reclassified as BF. Protected areas were state forest lands (‘forêts classées’) in Senegal and traditionally protected areas (‘bois sacrés’) in Mali.
Diameters were measured at 1.3 m above ground for all trees with diameters larger than 5 cm, except within the traditionally protected reserves in Mali since the protection allowed the surveyors to only estimate but not actually measure tree diameters within this type of landuse. For trees that branched below 1.3 m or had been coppiced earlier below 1.3 m, diameters were measured at 0.3 m above ground. The species identity of each tree was established in the field. Arbonnier (2004) was used to harmonize species names between countries as sometimes different synonyms were used for the same species in the different countries.
Tree richness and abundance calculations
Coupled datasets containing information on tree communities (tree abundance for a crosstabulation of quadrats × species) and environmental descriptors (quadrats × explanatory variables) were analysed with the freely available Biodiversity.R statistical software (Kindt and Coe 2005) that was developed for the R 2.1.1 statistical language and environment (R Development Core Team 2005) and often uses the vegan community ecology package (Oksanen et al. 2005).
The software calculated total and average species richness for each landuse category, each village and all possible combinations of villages and landuse categories. The same approach was used for average tree abundance. The hypothesis that average species richness was larger within particular landuse categories was tested with generalised linear regression models (GLM) with logit link and negative binomial variance functions (Hastie and Pregion 1993; Kindt and Coe 2005), using landuse and village as categorical explanatory variables. The abundance of trees in the quadrat was included as an additional continuous explanatory variable in another GLM model with the same other parameters.
Species accumulation curves
An exact calculation method for site-based species accumulation has been developed recently by various authors (Ugland et al. 2003; Colwell et al. 2004; Kindt et al. 2006), whereas the traditional method was based on a permutation procedure that calculated numerous random sequences of sites (Gotelli and Colwell 2001). Exact site-based species accumulation curves were estimated by the specaccum function of the vegan package (Oksanen et al. 2005) for the entire dataset and for subsets of the data of quadrats belonging to the same landuse category. Species accumulation curves were scaled by number of quadrats and by average number of trees per accumulated quadrats (Gotelli and Colwell 2001).
Exact and random site-based species accumulation curves show the result of random selection of sites from the entire population of available sites. Since different numbers of quadrats were sampled from the different villages (Table 3), villages with higher numbers of quadrats had larger influence on the obtained species accumulation curves. To balance the influence of each village on the species accumulation curves, we restricted species accumulation to the same number of quadrats from each village. A randomisation approach was used by which the first village was randomly selected from the available villages, a fixed number of quadrats (the subsample size) were randomly selected from the village, a second village was randomly selected from the remaining villages and the same number of quadrats was randomly selected from the second village. The procedure continued by randomly selecting new villages and quadrats from these villages until all villages were selected. At the end of the selection process, the sequence of all quadrats was randomized. Similar to the randomisation procedure of site-based species accumulation, a species accumulation curve was obtained by counting the total number of species in the pooled quadrats and the entire procedure was repeated many times (in this case 1,000 permutations) to obtain the average number of accumulated species and a standard deviation for accumulated species richness. For a particular subsample size, only villages with total numbers of quadrats equal to or larger than the subsample size were included, thus avoiding the need to sample the same quadrat more than once. Village-balanced species accumulation curves were calculated for fixed subsample sizes of one and five quadrats per village in the reported results. The method for balanced species accumulation was integrated in the BiodiversityR package (balanced.specaccum function; Kindt and Coe 2005).
Influence of diameter classes on species richness
Every inventoried tree was included in one of the following diameter classes: class 1 with diameters smaller than 5 cm, class 2 (5–10 cm, excluding trees with exactly 10 cm diameter), class 3 (10–20), class 4 (20–40), class 5 (40–80) and class 6 (diameters of 80 cm or larger). A different set of accumulated diameter classes contained all trees smaller than 5, 10, 20, 40 and 80 cm, which allowed for the construction of species accumulation graphs (see below). Community datasets were obtained for each diameter class, which allowed the calculation of total species richness for each combination of landuse category, village and diameter class.
Two curves were constructed for each combination of landuse category and village. A first distinct diameter–species curve (DDS) linked the total species richness for each of the distinct diameter classes 1–6. A second accumulated diameter–species (ADS) curve showed the species accumulation patterns from the smallest to the largest diameters by combining the results for the accumulated diameter classes with the results for all trees. Standardisation to the same sample size was not required since the shape of the two curves was the topic of investigation and not whether richness was larger in some villages or for some landuse categories. It was therefore possible to superimpose graphs from several villages onto the same plot, but the same plot should not be used to make comparisons of richness between villages.
Our approach of studying diameter distribution was different from a species-specific approach of looking at size class distribution of trees of the same species to infer vegetation dynamics (e.g., Lykke 1998). Whereas the species-specific approach identifies species that are not susceptible to extinction by identifying significantly higher numbers of trees in the smallest diameter class, our approach uses a more acute identification of extinction dangers by investigating how many species have no (and not just the same or fewer) trees with smaller diameters.
The shape of the DSD and ASD curves was analyzed graphically and by a generalised linear model with quasi-Poisson variance and logarithmic link functions, using species richness of a diameter class as response variable, village as categorical explanatory variable and diameter code [1–6] as continuous explanatory variable (Hastie and Pregion 1993; Kindt and Coe 2005). Statistical evidence for a regression coefficient for the diameter code to be significantly different from zero was interpreted as proof of a general ascending (positive coefficient) or descending (negative coefficient) shape of the curves.
Results
Tree species
About 110 tree species were encountered in the 300 quadrats and 16 villages (Tables 2 and 3). The total number of trees was 13 633. On average, a quadrat contained 45 trees and 6.1 species (Table 3). Twelve species were found in the four surveyed countries, 13 species were encountered in three countries, whereas half of all species were only encountered in one country (Table 2). Abundances over 100 trees were observed for 20 species, whereas 50 species had fewer than 10 individuals (Table 2).
The majority of tree species were indigenous, as only ten exotic species were encountered (Table 2). Only four species were not listed by Arbonnier (2004), which is an indication that the majority of encountered species are relatively common and widespread species that have frequently been used traditionally in the dry zones of west Africa, since Arbonnier used these criteria to select the 350–400 species described in his book. The four species that were not listed by Arbonnier included one exotic species (Eucalyptus alba) and one species that is known to be relatively common, although we only encountered one tree of this species (Senna occidentalis). The described habitats for the surveyed indigenous species included Sahelo-Sudanian or Sudano-Sahelian vegetation types (54 species), Sudanese vegetation (57 species) and the Sahel (24 species) (Arbonnier 2004, Table 2). The aforementioned habitats were not provided for seven indigenous species with Sudano-Guinean habitats (Table 2), two species with Guinean habitats (Newbouldia laevis and Terminalia laxiflora) and one species with a pantropical distribution (Tamarindus indica).
The 88 indigenous species listed by White (1983) included 55 species that are characteristic for the Sudanian regional centre of endemism and 46 species listed for the Sahel regional transition zone. Most of the Sudanian species were categorised by White (1983) as characteristic for both drier and wetter parts (25 species) or the drier northern part (16 species) of Sudanian woodland, whereas no species was typical for the wetter southern part of this vegetation type. All seven species listed by White (1983) as main woody species for the Sahel wooded grassland were encountered, with exception of Leptadenia pyrotechnica. Seven species were listed by White to belong to other phytochoria, mainly the Guineo-Congolian (Cissus quandrangularis, Holarrhena floribunda and Newbouldia laevis) and the Lake Victoria (Capparis tomentosa and Grewia bicolor) phytochoria.
Distribution of species richness over villages and landuse categories
Most of the species were found in BF (84 species) and VF (70 species), although the higher species richness in these landuse categories is also partially a result from their larger sample sizes (Table 3). The consequence of lack of balance in the dataset (not having the same number of quadrats within each landuse category) is that it is not meaningful to compare the total number of species of different landuse categories within the same village (Table 3). Since differences in numbers of quadrats are likely to contribute to differences in total species richness (given the expectation that more species will be encountered on a larger number of quadrats), a more careful approach of comparing total species richness between different categories of landuse is to make inferences from species accumulation curves (see below).
The average species richness is highest in the reserves (9.3 species), smallest in the VF (5.0) and slightly above the species richness of the VF in BF and the SZ (Table 3). Regression results (GLM with negative binomial variance and log link functions) confirmed the lower species richness of VF (regression coefficient: −0.31, P = 0.0003) compared to BF (intercept: 1.28). The regression results further suggested higher species richness of the SZ (coefficient: +0.60, P < 0.0001) compared to the BF and provided limited evidence for higher species richness within reserves (coefficient: +0.18, P = 0.12).
Most of the explained deviance could be contributed to differences between villages as village differences explained 40.6% of deviance (type-II ANOVA, P < 0.0001), whereas differences in landuse explained 6.5% of deviance (P < 0.0001). Average species richness of a quadrat was significantly smaller in the third village from Niger and significantly larger in all villages from Senegal (except village S4) than compared to the first village of Burkina Faso. Especially the result from Niger was surprising, since quadrat size was twice as large in Niger than in all other countries. Including the number of trees as an additional explanatory variable in the GLM resulted in the same patterns for landuse and only one change for village differences by indicating lower species richness in the second village from Burkina Faso.
Pairwise comparisons within villages show that the general trend of higher average species richness in the BF compared to the VF was not observed in several villages, as in the first Malian village and two villages in Senegal (Table 3). Species richness was higher in the SZ compared to the bush fields for four villages out of the six villages with both these landuse categories (Table 3).
Investigation of the species accumulation patterns by balanced species accumulation curves (constructed by randomly selecting one or five quadrats from each village with a particular landuse) showed that average species richness was lowest in the VF and highest in the reserves (Fig. 2a, b). The same graphs showed that the species accumulation curves of BF and the SZ were almost identical when scaling the curves by the number of quadrats. The unbalanced species accumulation curves showed that the largest species totals observed for bush and village fields was an effect of their larger number of quadrats (Fig. 2b). Accumulated species richness in the SZ was the lowest of all landuse categories for the unbalanced species accumulation, whereas this was not the case for the balanced accumulation (Fig. 2b). This finding demonstrated a strong effect of differences between villages in numbers of quadrats for the same landuse. Two villages (N3 with 18 quadrats and B2 with 10 quadrats out of the total of 44 quadrats; Table 3) dominated the quadrats of the SZ. This lack of balance had a pronounced effect on species accumulation as it resulted in contrasting patterns.”

Species accumulation curves for different landuse categories plotting species richness against accumulated number of quadrats (top) or number of trees (bottom) for different methods of sampling (left: balanced sampling sampling of 1 quadrat per village; right: balanced scaling of 5 quadrats per village). Error bars indicate standard deviations for balanced samples for different landuse categories
When scaling the species accumulation curves by the average number of accumulated individuals (Fig. 2c, d), the relative positions for the different landuse categories were fundamentally different than those scaled by numbers of quadrats (Fig. 2a, b). Based on the same number of accumulated individuals and randomly selecting one quadrat per village, species richness was highest for VF, followed by BF, reserves and finally the SZ. When randomly selecting five quadrats per village, the species richness was lowest in reserves and remained highest for the BF and VF.
Distribution of species richness over diameter classes
The majority of species had trees in the smallest diameter class (105 species), whereas 38–54 species were encountered in the intermediate diameter classes (5–10, 10–20, 20–40 and 40–80 cm) (Table 2). Only 21 species had trees in the biggest diameter class, dominated by Adansonia digitata (76 trees, 39% of all trees with largest diameters), Faidherbia albida (36 trees, 18%) and Balanites aegyptiaca (31 trees, 16%) (Table 2). These species were also identified as priority species for domestication, whereas four other priority species had trees in the biggest diameter class (excluding Ziziphus mauritiana and Lannea microcarpa that had no trees larger than 80 cm). The colonizing species Guiera senegalensis had the largest number of trees [a pioneer species of post-cultural succession, Devineau (1999)], but no trees in the largest diameter class. Of the 21 species with trees in the largest diameter class, five species had more trees of largest diameter than trees of smallest diameter (Adansonia digitata, Celtis integrifolia, Prosopis africana, Ficus platyphylla and Ficus sycomorus) (Table 2).
Most of the villages had more than five species in the smallest diameter class for the landuse categories that they contained, with exceptions for two villages (13%) for BF, two villages (17%) for VF, one village (25%) for reserves and one village (17%) for the SZ (Fig. 3). Substantially fewer species were observed in the biggest diameter class in all the villages, as no village had more than five species in this diameter class. In only one village (7%) was a larger number of species observed in another diameter class than the smallest class for bush fields, in two villages for VF (17%), no villages for reserves and three villages (50%) for the SZ (curves with closed symbols show the villages where some diameter classes have larger total species richness than the smallest diameter class in Fig. 3).

Species richness for different diameter classes, villages and landuse categories (a, b: bush fields; c, d: village fields; e, f: reserves; g, h: sylvopastoral), with species richness in separate diameter classes on the left and accumulated species richness over diameter classes on the right (see Materials and methods). Curves with open and closed symbols follow different trends that are discussed in the text
On average, species richness declined 13.1 species for BF, 8.5 species for VF, 11.7 for reserves and 11.3 species for the SZ from the smallest to the largest diameter class (Fig. 3). Regression results confirmed the decline of species richness in increasing diameter classes for BF (intercept: 2.3; diameter class: −0.39, P < 0.0001), village fields (intercept: 2.4; diameter class: −0.28, P < 0.0001), reserves (intercept: 2.01; diameter class: −0.34, P < 0.0001) and the SZ (intercept: 2.5; diameter class: −0.48, P < 0.0001).
Despite the general decline in numbers of species with increasing diameter class, in only four cases (one in BF and three in VF) were no new species encountered in larger diameter classes (curves with closed symbols show these cases on the right-hand site of Fig. 3). The average increment in accumulated number of species from the smallest to the largest diameter class was 4.5 for BF, 2.7 for VF, 7.2 for reserves and 2.8 for the SZ. The observed accumulation of species was confirmed by regression analysis for BF (intercept: 1.9; diameter class: +0.05, P < 0.0001), village fields (intercept: 2.1; diameter class: +0.05, P < 0.0001), reserves (intercept: 1.5; diameter class: +0.08, P < 0.0001) and the SZ (intercept: 1.4; diameter class: +0.04, P = 0.0026).
Discussion
Influence of balance of sampling on species accumulation patterns
Our results demonstrated a strong influence of balancing in obtaining the smallest species accumulation for the sylvopastoral zone when sampling was not balanced over villages, whereas this was not the case when sampling the same number of quadrats from each village. Researchers should be aware of influence of spatial levels of sampling when they compare species accumulation curves that are obtained from samples that are obtained from different sets of spatial samples.
The balancing approach could be expanded for the estimation of the total number of species in the area (gamma diversity). Rosenzweig et al. (2003) demonstrated that hyperdispersion of sampling resulted in better estimates of gamma diversity than random or clustered techniques. Hyperdispersion of sampling is obtained in our approach by balancing the samples from different spatial strata.
Our approach also suggests that differences in sample sizes of strata that were not measured (for example, strata related to differences in years of cultivation or strata related to differences in distance to natural ecosystems) could also have influenced the species accumulation curves. Researchers should also be aware of the potential effect of differences in the average distance between quadrats, as balanced species accumulation curves may hide some differences between different strata. Imagine a first type of landuse where on average one new species is encountered between quadrats sampled at 400 m intervals and a second type of landuse where on average one new species is encountered on quadrats at 900 m intervals. Balanced accumulation curves using sampling at intervals of 400 m for the first type of landuse and 900 m intervals for the second type of landuse would not reveal differences between these two landuses. However, when balanced sampling would be done at the same distance interval, then it would become obvious that species accumulate more rapidly in the first type of landuse.
Differences between landuse categories
We found small but significant differences in the average numbers of species in the different landuse categories. These differences became more pronounced when pooling quadrats.
In contrast to our classification system of BF, village fields, SZs and reserves, other authors have described parkland systems as consisting of crop fields, fallows of different ages and FR (e.g., Mortimore et al. 1999; Maranz and Wiesman 2003; Augusseau et al. 2006). In some villages, ‘FR’ were actually old fallows as natural vegetation was not present (Augusseau et al. 2006). The study of Augusseau et al. (2006) in Burkina Faso showed that tree richness declined with field age and increased with fallow age as a result of tree selection by farmers in cultivated fields and natural regeneration in fallows. Since SZs consist of old fallows, the similar site-based species richness of BF and the SZ that we encountered could be an artefact of the same species richness on average for fields and fallows, whereas species richness would be different when considering the time since beginning and end of the cultivation cycle, especially when comparing old fallows with old fields. A shorter fallow period for VF could also explain the lower species richness within this type of landuse if distance from the village centre was confounded with length of the fallow period.
In an analysis of the effect of agricultural intensification in Burkina Faso, Gray (2005) demonstrated that tree density was lower when fields had been cultivated for longer but also when a larger percentage of the area was under animal traction. These studies suggest to include years since cultivation, years since last fallow and method of land clearing as additional explanatory variables for tree diversity within villages besides the sylvopastoral or field landuse typology.
Based on the largest species richness at the end of the fallow phase, Augusseau et al. (2006) recommend to manage tree diversity at a landscape level and to maintain fallows as areas for recovery of tree diversity within a mosaic of other forms of landuse. However, as several species are eliminated during the cultivation cycle and as there is evidence that fallow periods are shortening, it may not be possible for many species to reach maturity within fallows (see below). Information from farmers in Niger, for example, showed that fallow periods had declined from 15 years or longer to less than 5 years (Wezel and Boecker 1998). There is also some evidence that fallowing is no longer practised in some areas with high human and livestock population pressures (Teklehaimanot 2004). The policies in Mali also contribute to shortening of fallow periods as land that is left in fallow for more than 10 years returns to the protected state domain (Ashley et al. 2006).
Comparisons scaled by the average number of trees showed that species accumulation was lower in reserves and the sylvopastoral zone than in BF and VF. This finding shows that farmers select for higher diversity for the same number of trees. This pattern conforms to the anthropogenic selection of tree diversity of parkland systems (Boffa 1999; Maranz and Wiesman 2003; Larwanou and Saadou 2005; Augusseau et al. 2006).
Species diameters
A general pattern was observed in all landuse categories by which many species were encountered in the small diameter classes and few species in the largest classes. These results should not be interpreted as contradicting the results of species-specific studies that have shown a general decline of most tree species in the dry areas of western Africa (e.g., Gonzalez 2001; Wezel and Lykke 2006). In a study based on diameter distributions for a forest in Senegal with a frequent fire regime, there was evidence that many species are not rejuvenating well as they have few trees in the smaller diameter classes (Lykke 1998). We documented a more acute pattern resulting from problems with regeneration, as there was statistical evidence that several species had no trees within the smallest diameter classes. Based on the reasoning that species will only regenerate into the largest class if they have a reverse-J population structure (e.g., Lykke 1998), more species may be in danger of extinction than the species that we found accumulating in ASD curves.
Another pattern that we demonstrated (SDS curves) is that most species had no trees in the larger diameter classes. Such pattern could be an indication of another emerging problem with regeneration of species within parklands: that not enough mature trees remain to produce the next generation (Boffa 1999). Such pattern may not be prevalent yet as we did record highest species richness in the smallest diameters (and thus evidence that regeneration is still taking place, possibly mainly from the soil seed bank), but could become a problem in the future.
Theoretically, the smaller numbers of species and trees with large diameters could be an effect of small quadrat sizes. Small quadrat sizes were demonstrated to result in a humped relationship between plant biomass and species richness as there are limitations for space for larger plants (Oksanen 1996). Given that the largest numbers of species for a landuse and village was never larger than five for the largest diameter class (Fig. 3), given the dimensions of the quadrat that were used (minimum 50 × 20 m2, except 25 × 20 m2 for FR in Senegal) and given that in some cases 10 or more quadrats were sampled of the same landuse and village (Table 3), we do not expect that the low species richness for larger trees is an effect of space limitations but rather of real decline in species richness. Species-specific research by which lack of regeneration is documented at a landscape level may unequivocally confirm the decline of most species in the villages that were surveyed, although we think that the priority of research should now be on exploring ways of improving regeneration of species rather than confirmation of species’ declines.
For those that are not familiar with the realities of many protected areas in west Africa, the results of small species richness in the biggest diameter classes are very surprising. The same pattern was documented for a classified forest in Mali where the only large trees were baobabs (Adansonia digitata) and karité (Vitellaria paradoxa) (Ashley et al. 2006). Lykke (2000) mentioned that those species that are most preferred by local populations and are therefore protected in agricultural fields, are also the species that are overexploited in reserves. Although tree density was smaller, tree biomass was larger in Nigerian parklands compared to unprotected natural woodlands (Mortimore et al. 1999). Ashley et al. (2006) mention that exploitation of reserves is often by outsiders, suggesting that providing special rights and responsibilities to surrounding villages could increase conservation of remaining reserves. If regeneration and large enough population sizes of adult trees could be conserved in reserves in the future, then these could become areas of seed sources for the surrounding villages.
However, given the current degradation of reserves in most Sahelian countries, the possibility of tree conservation within farmer-managed agroforestry parklands should not be overlooked. Moreover, as various forest and land related legislations do not sufficiently take into account tree management, including the tenure of trees and land (Ly et al. 2006), agroforests would need special protection if conservation of tree diversity on-farms is targeted. If shortening and eventual abandonment of fallowing is the general trend in agroforestry parklands and if the tree diversity patterns documented by our surveys are representative for these anthropogenic savanna systems, then enrichment planting with selected high-value tree species, protection of individual trees or protection of trees in the soil seed bank [for example by farmer-managed natural regeneration, including limitations of grazing (Rinaudo 2001; Cunningham and Abasse 2005)] may be necessary within cultivated fields and regeneration during the fallow phase may not longer be sufficient to maintain tree populations. As such management practices are not a part of the traditional management of parklands (Lykke et al. 2004), new approaches to tree regeneration need to be explored further.
The majority of encountered species in the surveys were common indigenous tree species that are typical of the dry areas of west Africa, conform other studies in dry west Africa (e.g., Augusseau et al. 2006; Wezel and Lykke 2006). In an ethnobotanical study conducted in the southern part of Burkina Faso (within the undifferentiated Sudanian woodland zone of White 1983), 50 informants listed 81 useful species for edible fruit, vegetable sauce, firewood, construction and medicine (Kristensen and Balslev 2003). The same study demonstrated that several species were rejected for firewood as there were traditional prohibitions against this use. Such situations may result in decreasing population sizes for the useful species, although informants in the study only listed 16 species to become rare (Kristensen and Balslev 2003) and long-term habitation of the area had not resulted in a disappearance of these species. Our four country study showed that many species were rare (only 20 species had more than 100 trees in the total survey), which confirms to a pattern that useful species may become overexploited.
For a study in Sudanian woodland that is naturally dominated by Isoberlinia (White 1983), Devineau (1999) indicated that current tree communities are adapted to the current situation, whereas species that have become rare are typical of less disturbed vegetation types. Similarly, Lykke (1998) mentions that species composition in a protected forest in Senegal is shifting towards shrubby species that are adapted to cope with fire and other forms of long-term disturbance. Based on the dual threats of overexploitation and problems with disturbance by fire, it therefore seems likely that useful species that can not cope with fire are most susceptible to extinction in the area and that special emphasis may be required to ensure the regeneration of these species.
Augusseau et al. (2006) documented a third trend that may result in the decline of indigenous tree species as they describe a trend towards species that provide cash income. In a village in Burkina Faso, farmers indicated that only five species were important of which the two exotic species [cashew (Anacardium occidentale) and mango (Mangifera indica)] were the only ones that were planted. Documenting patterns in tree increments or decreases from Burkina Faso, Niger and Senegal, Wezel and Lykke (2006) found that the only new species in villages were exotic species. The same pattern was also observed in another study from Niger (Larwanou and Saadou 2006). As reforestation and afforestation programs in the Sahel region have mostly promoted the planting of fast growing exotic tree species such as Australian acacias, eucalypts, melina, neem and teak. (Evans and Turnbull 2004), the trends may only change if future tree promotion programs include indigenous tree species. The success of promotion campaigns for indigenous tree species could be substantially enhanced if they were accompanied by improvements of the marketing opportunities for indigenous tree products, advances in the domestication of indigenous tree species (such as the selection of more productive cultivars of Tamarindus indica, Vitellaria paradoxa and Ziziphus mauritiana as currently undertaken by the World Agroforestry Centre and its partners) and changes in policies that currently discourage farmers from planting indigenous tree species.
References
Abegg C, Bayala J, Belem M, Kalinganire A (2006) Analyse comparative de facteurs socio-économiques influençant la biodiversité ligneuse des parcs agroforestiers du plateau central du Burkina Faso. J Forest Suisse 157(1):17–23
Arbonnier M (2004) Trees, shrubs and lianas of West African dry zones. CIRAD, Margraf Publishers, MNHN, Paris
Ashley R, Russell D, Swallow B (2006) The policy terrain in protected area landscapes: challenges for agroforestry in integrated landscape conservation. Biodivers Conserv 15:663–689
Augusseau X, Nikiéma P, Torquebiau E (2006) Tree biodiversity, land dynamics and farmers’ strategies on the agricultural frontier of south-western Burkina Faso. Biodivers Conserv 15:613–630
Boffa JM (1999) Agroforestry parklands in sub-Saharan Africa. FAO Conservation Guide No 34. Food and Agriculture Organization of the United Nations, Rome
Burgess N, D’Amico Hales J, Underwood E, Dinerstein E, Olson D, Itoua I, Schipper J, Ricketts T, Newman K (2004) Terrestrial eco-regions of Africa and Madagascar. Island Press, Washington
Colwell RK, Mao CX, Chang J (2004) Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717–2727
Cunningham PJ, Abasse T (2005) Reforesting the Sahel: farmer managed natural regeneration. In: Kalinganire A, Niang A, Kone B (eds) Domestications des espèces agroforestières au sahel: situation actuelle et perspectives. ICRAF Working Paper 5. World Agroforestry Centre, Nairobi, pp 75–80
Devineau JL (1999) Seasonal rhythms and phonological plasticity of savanna woody species in a fallow farming system (south-west Burkina Faso). J Trop Ecol 15:497–513
Evans J, Turnbull JW (2004) Plantation forestry in the tropics: the role, silviculture, and use of planted forests for industrial, social, environmental, and agroforestry purposes. Oxford University Press, New York
GEF (2002) Desert Margins Programme (DMP): Project Document. United Nations Environment Programme. Global Environment Facility (GEF), Nairobi
Gonzalez P (2001) Desertification and a shift of forest species in the West African Sahel. Clim Res 17:217–228
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Gray LC (2005) What kind of intensification? Agricultural practice, soil fertility and socio-economic differentiation in rural Burkina Faso. Geogr J 171:70–82
Hastie TJ, Pregibon D (1993) Generalised linear models. In: Chambers JM, Hastie TJ (eds) Statistical models in S. Chapman and Hall, London
Kalinganire A, Kaya B, Niang A, Kindt R, Muraya P, Coe R (eds) (2005) Caractérisation de la biodiversité ligneuse dans les sites du programme des zones en marge du désert: Manuel de procédures. Occasional Paper No 3. World Agroforestry Centre, Nairobi
Kindt R, Coe R (2005) Tree diversity analysis. A manual and software for some common statistical methods for biodiversity and ecological analysis. World Agroforestry Centre (ICRAF), Nairobi. URL http://www.worldagroforestry.org/treesandmarkets. Accessed 22 Jan 2008
Kindt R, Van Damme P, Simons AJ (2006) Patterns of species richness at varying scales in western Kenya: planning for agroecosystem diversification. Biodivers Conserv 15:3235–3249
Kristensen M, Balslev H (2003) Perceptions, use and availability of woody plants among the Gourounsi in Burkina Faso. Biodivers Conserv 12:1715–1739
Larwanou M, Saadou M (2005) Biodiversity of ligneous species in semi-arid to arid zones of southwestern Niger according to anthropogenic and natural factors. Agric Ecosyst Environ 105:267–271
Larwanou M, Saadou M (2006) Arbres dans les systèmes agraires en zone sahélienne du Niger: Mode de gestion, atouts et contraintes. Tropicultura 24:8–14
Ly I, Kalinganire A, Niang A (2006) Essai d’analyse de la prise en compte des systemes agroforestiers par les législations forestières au sahel: Cas du Burkina Faso, du Mali, du Niger et du Sénégal. ICRAF Working Paper 29. World Agroforestry Centre, Nairobi
Lykke AM (1998) Assessment of species composition change in savanna vegetation by means of woody plants’ size class distributions and local information. Biodivers Conserv 7:1261–1275
Lykke AM (2000) Local perceptions of vegetation change and priorities for conservation of woody-savanna vegetation in Senegal. J Environ Manage 59:107–120
Lykke AM, Kristensen MK, Ganaba S (2004) Valuation of local use and dynamics of 56 woody species in the Sahel. Biodivers Conserv 13:1961–1990
Maranz S, Wiesman Z (2003) Evidence for indigenous selection and distribution of the shea tree, Vitellaria paradoxa, and its potential significance to prevailing parkland savanna tree patterns in sub-Saharan Africa north of the equator. J Biogeogr 30:1505–1516
Mortimore M, Harris FMA, Turner B (1999) Implications of land use change for the production of plant biomass in densely populated Sahelo-Sudanian shrub-grasslands in north-east Nigeria. Glob Ecol Biogeogr 8:243–256
Oksanen J (1996) Is the humped relationship between species richness and biomass an artefact due to plot size? J Ecol 84:293–295
Oksanen J, Kindt R, Legendre P, O’Hara RB (2005) Vegan: community ecology package version 1.7-82. URL http://cc.oulu.fi/~jarioksa/
R Development core team (2005) R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna. URL http://www.R-project.org
Rinaudo T (2001) Utilizing the underground forest: farmer managed natural regeneration of trees. In: Pasternak D, Schlissel A (eds) Combating desertification with plants. Kluwer Academic/Plenum Publishers, New York, pp 325–336
Rosenzweig MI, Turner WR, Cox JG, Ricketts TH (2003) Estimating diversity in unsampled habitats of a biogeographical province. Conserv Biol 17:864–874
Teklehaimanot Z (2004) Exploiting the potential of indigenous agroforestry trees: Parkia biglobosa and Vitellaria paradoxa in sub-Saharan Africa. Agroforest Syst 61:207–220
Ugland KI, Gray JS, Ellingsen KE (2003) The species-accumulation curve and estimation of species richness. J Anim Ecol 72:888–897
Wezel A, Boecker R (1998) Fallow plant communities and site characteristics in semi-arid Niger, West Africa. J Arid Environ 40:269–280
Wezel A, Lykke AM (2006) Woody vegetation change in Sahelian Africa: evidence from local knowledge. Environ Dev Sustain 8:553–567
White F (1983) The vegetation of Africa. A descriptive memoir to accompany the Unesco/AETFAT/UNSO vegetation map of Africa. United Nations Educational, Scientific and Cultural Organization, Paris
Acknowledgements
We are very grateful for the collaboration provided by farmers during data collection. We thank GEF (DMP project) and IDRC (Tree Biodiversity Project) for funding data collection and analysis, the governments of Burkina Faso, Mali, Niger and Senegal for co-funding data collection (through INRA, IER, INRAN and ISRA, respectively), and Peter Muraya and Richard Coe for helping us with data preparation and initial analysis. Roeland Kindt thanks VVOB for co-funding training workshops for African researchers in methods of biodiversity analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kindt, R., Kalinganire, A., Larwanou, M. et al. Species accumulation within land use and tree diameter categories in Burkina Faso, Mali, Niger and Senegal. Biodivers Conserv 17, 1883–1905 (2008). https://doi.org/10.1007/s10531-008-9326-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-008-9326-3