
Abstract
The successful establishment of an exotic species is based on its resource-based habitat, and thus depends on its encountering the interaction partners and environmental conditions it requires to complete its life cycle. We examine factors that define the resource-based habitat of the European common blue butterfly (Polyommatus icarus Rottemburg, [1775] (Lepidoptera: Lycaenidae)) recently introduced in Quebec, testing the role of exotic host plants and of short vegetation structure. Field studies surveyed adult butterfly abundance in lightly managed urban sites and related it to host plant cover and vegetation structure. Oviposition behavior was also recorded and linked to individual plant traits, namely height, species and anatomical structures. Results show that adult butterflies maintain the same host plant associations as in the native range, suggesting a role for naturalized plants in facilitating the establishment of this exotic herbivore. However, relative oviposition rate was not higher on the most strongly associated host plant (birdsfoot trefoil, Lotus corniculatus, Fabaceae) suggesting no clear behavioral preference for this plant. Butterflies also showed the same association with short vegetation documented in the native range, although the basis for this preference remains unclear. P. icarus uses different biotopes in its invaded range (lightly managed urban sites) than in its native range (unimproved grasslands), and our results show the role of host plants and of short vegetation structure in defining the resource-based habitat that can predict its future expansion. Our results raise interesting questions about why this butterfly uses cities more in its invaded than its native range. The role of cities in insect conservation is increasingly recognized; we show how, paradoxically, a butterfly whose native biotopes are declining has found suitable conditions to thrive in far-removed urban centers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plant and animal species are being transported across the globe at increasing rates. Whether an new arrival establishes successfully or even becomes invasive depends less on the biotopes and more on the biotic interactions and environmental conditions it encounters in the new location (Wisz et al. 2013). Indeed, a recently arrived exotic species needs to form associations with interaction partners and establish itself within the community structure in its new location (Agosta 2006). Potential distribution range is thus best modelled, not based only the biotopes occupied by the species in its native range, but based on where all the resources it needs to complete its lifecycle can be found—this is known as the resource-based habitat definition (Dennis et al. 2003). This can include novel biotopes or vegetation types (ones that can be either unused or unavailable in the native range) if all resources necessary to complete the life-cycle are present (Vanreusel and Van Dyck 2007; Dennis et al. 2014). Biotopes occupied in the invasion range can thus differ from those used in the native region, depending on availability of interaction partners.
For herbivorous insects, host plants are often the most significant interaction partner, defining habitat quality (Dennis et al. 2003). Herbivorous insects can only persist in areas in which they can complete their life cycle, i.e., where larval host plants are present. However, herbivorous insects are known to rapidly colonize new host plants when an opportunity arises via ecological fitting between their traits and those of the plants (Agosta 2006). Introduced plants, including crops, rapidly accrue a suite of native herbivores, and many introduced herbivorous insects form associations feeding on native plants. All that is required is that the insect recognize the plant as a suitable substrate for feeding or oviposition, and that it survives to produce some offspring, achieving some realized fitness (Agosta 2006). These novel interactions can lead invading insects to occupy new biotopes in their invasion ranges.
However, presence of acceptable host plant is not sufficient to define resource-based habitat, since insects need other resources during their life cycle. Most of these remain ill-defined such that, even for butterflies, some of the best studied insects, habitat quality is often measured only in terms of host plant availability (Dennis et al. 2014). However, vegetation structure has been shown to be an important element of resource-based habitat for butterflies (Berg et al. 2013; Habel et al. 2016; Aguirre-Gutiérrez et al. 2017). It can directly impact utilities-category resources (e.g. roosting, pupation or overwintering sites), or have indirect effects mediated either by quality of consumable resources (e.g. host plant defensive chemistry, flowering phenology etc.) (Dennis et al. 2014) or by interaction partners at other trophic levels (e.g. predators, parasitoids, mutualists) (Valdés and Ehrlén 2018). In general, higher vegetation hosts higher butterfly species diversity, but specialists often prefer low vegetation (Pöyry et al. 2006). Predicting species distribution based on resource-based habitat combined with ecological fitting thus implies that introduced herbivorous insects may use different biotopes and even different host plants in the invaded range than in the native one and that both availability of potential host species and vegetation structure can play a role in defining a herbivore’s habitat in its new range.
Cities are hotspots for biological invasions (New 2015); introduced species can often establish in the novel ecosystems assembled in cities by ecological fitting even if they are unable to compete in the co-evolved networks observed in less disturbed areas (Cadotte et al. 2017). Several different explanations have been proposed for the greater success of invasive species in cities, including higher propagule pressure, invasional meltdown, and niche opportunity due to competition/predation release that decreases biotic resistance (New 2015). For herbivorous insects, the presence of exotic plants can facilitate their establishment since it provides opportunities to re-establish the same interactions as they had in their native range (Johnson et al. 2019; Mech et al. 2019). This bottom-up facilitation can fast-track invasion by herbivorous insects (Engelkes and Mills 2013). Moreover, the unique community assemblages in cities can provide niche opportunities unavailable in the absence of more established co-evolved relationships (Cadotte et al. 2017) because of differences in both bottom-up and top-down pressure on insect herbivores (Harvey et al. 2010).
We use the invasion of a common European grassland butterfly in Quebec to examine the role of plant species and vegetation structure in its establishment in urban areas. The European common blue butterfly Polyommatus icarus Rottemburg, [1775] (Lepidoptera: Lycaenidae) was introduced in the Montreal area around 2005 (Hall 2007), has since been spreading locally (Al-Rewashdy et al. 2010) and has formed a myrmecophilous relationship with at least one native ant (Dexheimer et al. 2021). In its native range, P. icarus feeds on open-area ruderal Fabaceae plants and is not particularly associated with cities (Dennis et al. 2017); its main habitats are unimproved grasslands, rough downs and coastal dunes, and it also uses heaths and woodland clearings (Emmet and Heath 1989; Habel et al. 2016). Many of P. icarus ’s host plants have long been naturalized across North America, including the Montreal region (Turkington and Franko 1980). In Canada, P. icarus is more abundant at moderate than at low disturbance, and is associated with cities at the landscape scale (Rivest and Kharouba 2021). The blue butterfly seems to have a limited presence if at all in native ecosystems since observations are scarce beyond urban areas (GBIF Secretariat 2021; eButterfly 2022). We test whether association with exotic host plants favors establishment in cities.
Like other Lycaenids (Valdés and Ehrlén 2018), P. icarus is associated with short vegetation at the larval, but not necessarily at the adult, stage (León-Cortés et al. 1999; Habel et al. 2016). Proposed explanations have been linked to microclimate, lower rates of predation, higher rates of myrmecophily, or greater availability of overwintering sites. The best predictors of P. icarus abundance have been shown to be the presence of the host plant birdsfoot trefoil (Lotus corniculatus, Fabaceae) and low vegetation height, in both the British isles (León‐Cortés et al. 1999) and Germany (Habel et al. 2016). Although mechanistic explanations remain unclear, we test if this relationship still holds in new biotopes in P. icarus’ invasion range.
We examine factors that favor the establishment of the European P. icarus in Canadian urban sites, testing whether the butterfly’s resource-based habitat is defined by specific host plants and/or by vegetation structure. We first record the use of various host plants relative to their availability to examine whether P. icarus uses the same host plants as in its native range, testing the hypothesis that presence of known host plants in cities allows the butterfly to re-establish relationships (bottom-up facilitation). We next examine the role of vegetation structure: specifically, we ask whether low vegetation height in lightly managed urban meadows provide the favourable conditions found in this species’ vanishing native grassland habitat.
We observed P. icarus in the Montreal area in sites where the butterfly has been recorded by naturalists and related adult butterfly abundance to host plant cover and vegetation structure. Oviposition was also recorded and linked to individual plant traits, namely height, species and anatomical structures. This field study examines whether P. icarus associates with either exotic or native Fabaceae plants and tests the prediction that it prefers sites with short vegetation. These associations are further investigated with regards to oviposition.
Methods
Field sites
We selected field sites in the Montreal area by choosing locations where P. icarus had been observed by naturalists, based on records on the eButterfly and iNaturalist community science applications. Sites were chosen more than 1 km apart for statistical independence, based on data on the butterfly’s mobility in Europe (Gutiérrez et al. 2001). Sites were visited to confirm the presence of plants in the Fabaceae family. All Fabaceae were considered as potential hosts, and their actual use by P. icarus was evaluated as part of this study.
Remotely sensed images were used to measure area of different land use types within a 1 km buffer surrounding each field sites. We obtained data at GeoIndex accessed by Concordia University and used QGIS v3.10.5 to extract the proportion cover of land use types. Residential and industrial areas were dominant around the selected field sites, accounting for 40 and 21% of the total surface of the buffer areas respectively. Subsequent categories were conservation (11%) and recreational (10%) green spaces.
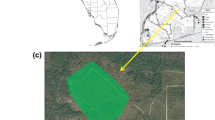
We visited each of the 20 selected sites once per month between 9:30 AM and 6:00 PM on a sunny calm day between June 16th and August 31st of 2020; there were 2–3 visits to each site (see Fig. 1) and a total of 56 visits. Percent cover by host plant species, vegetation height, abundance of adult butterflies and positioning of eggs were recorded at each visit.

Map showing location of 26 field sites in the Montreal region, with photographs taken at 3 of the sites to show the vegetation. Yellow flowers visible in one photo are L. corniculatus
Plant survey
The following Fabaceae plants were observed in at least one of the field sites: Lotus corniculatus, Medicago lupulina, Medicago sativa, Melilotus alba, Trifolium pratense, Trifolium repens, and Vicia cracca. All of these plants are native to Europe, naturalized in Canada and have been previously recorded as hosts for P. icarus in its native range (Emmet and Heath 1989; Robinson et al. 2010).
Linear transects of 100 m were established in each field site. If a field site was too small or irregular to contain a linear 100 m transect, the transect instead followed the edge of the site. Surfaces such as pavement or open water were excluded from the transect. These transects were used for evaluation of both plant cover and butterfly abundance.
Ten quadrats of 50 cm by 50 cm in size were drawn along the transects spaced 10 m apart. The percentage of surface area occupied by each host plant species was visually estimated to the nearest 5% within each quadrat. We used this information to relate butterfly abundance and oviposition behaviour to host plant abundance.
Vegetation height was measured in order to investigate its effect on habitat selection, since it has been suggested to be a predictor of P. icarus habitat use (Emmet and Heath 1989; León-Cortés et al. 1999; Habel et al. 2016). Three indices of vegetation were measured with a tape measure, from the ground up: maximum height reached by any plant in the quadrat, maximum height reached by a host plant from the list above, and average height of the vegetation (i.e. “canopy”). All three height indices were included in models, but proved to be highly correlated, such that only one was retained.
Butterfly survey
We estimated adult abundance with modified Pollard walks (Pollard and Yates 1994; Rivest and Kharouba 2021): the observer walked the 100 m transect at the pace of ~ 50 cm per second, such that the transect took about 5 min to complete. All butterflies observed within 5 m on either side of the transect were recorded and their sex noted. The transect was done twice per site visit, and the average value was used in analyses.
Egg survey
At each site, we searched all Fabaceae plants present for P. icarus eggs. No other Lycaenid species were observed as adults in the field sites, so Lycaenid eggs found on host plants were assumed to belong to P. icarus. When possible, female P. icarus were followed to observe oviposition events directly (see Fig. 2), after which the measurements for the egg were made immediately. When an egg was found, the following variables were measured: host plant species, anatomical structure on which the egg was found (young leaf, mature leaf, or flower), height above the ground at which the egg was found (cm), total height of the plant (cm).

Female P. icarus butterfly ovipositing in a Trifolium repens inflorescence in June 2020 on a mixed lawn patch adjacent to Saint Jacques street, Montreal. Photo: Eric Dexheimer
Statistical analysis
Counts of both adult butterflies and eggs were related to vegetation characteristics, including percent cover of the 7 host plants and the three measures of vegetation height (canopy, max. host plant height and max. plant height). The unit of replication was the observation (N = 56). Site and time period were included in the analysis as random factors: the time period factor was obtained by dividing the growth season into three periods: early (Julian date 161–192), mid- (JD 193–217) and late (JD 218–239) summer. Cut-off dates were placed to correspond to troughs in butterfly abundance.
Adult butterfly abundance was analyzed first with a correlation matrix and next with a generalized mixed model (based on Poisson distribution) including cover of all 7 host plants and 3 vegetation height indices as fixed factors and site and time period as random factors. Model simplification with AIC was used to identify which of these factors are significant predictors of adult abundance.
Host plant choice for oviposition was analyzed using a general linear model with plant species and time period as fixed factors and host plant cover as an offset to control for variation in proportion cover of different host plants on the different sites (Hosmer and Lemeshow 2000). A quasipoisson distribution was used to account for data overdispersion.
Height of oviposition site relative to total plant height was analyzed with another mixed model including plant species as a random factor. Finally, selection of plant structure for oviposition was examined with a chi-square on the four most commonly used host plants. All analyses were done in R (v. 3.6.2).
Results
A total of 365 butterflies were observed, and 213 eggs were counted over the course of the field season.
Butterfly abundance
The correlation matrix in Fig. 3 suggests that adult abundance is favored by L. corniculatus availability, and to a lesser extent by Medicago lupulina, Trifolium pratense and T. repens. Butterfly abundance is also negatively associated with all three measures of vegetation height. This matrix also suggests vertical structuring of the habitat by different plants: vegetation height is positively related to V. cracca abundance but negatively related to T. repens (and to a lesser extent T. pretense and M. lupulina) cover.

Spearman correlation matrix between butterfly abundance per observation and percent cover of each Fabaceae species, maximum vegetation height, average vegetation height (canopy) and maximum host plant height, all in cm
Model simplification using AIC retained a model with three terms: canopy height and percent cover of L. corniculatus and T. pratense (AIC of full 9-factor model: 118; 3-factor model: 112). A model with the same three fixed factors but excluding the random factors performed even better (AIC: 109) and gave very similar results: butterfly abundance increased with L. corniculatus cover (coefficient: 0.0238 (S.E. 0.005), p < 0.00001) and with T. pratense cover (coefficient: 0.047 (S.E. 0.0211), p = 0.031) but decreased with canopy height (coefficient: − 0.01 (S.E. 0.006), p = 0.0027).
Oviposition
Eggs were observed on all Fabaceae plants except T. repens which was removed from these analyses.
A general linear model with quasipoisson distribution showed that M. sativa was used more often relative to its abundance than any other plant, and significantly more so than L. corniculatus (main effect coefficient = 24.11 (S.E. 8.56), p = 0.0064). However, this effect decreases as the season progresses (interaction coefficient = − 2.80 (S.E. 1.25), p = 0.028). No other significant difference in the rate of host plant use relative to cover was detected (see Fig. 4), suggesting that P. icarus generally lays its eggs on whichever plants are available.

Average proportion of eggs on each host plant per site against average proportion host cover. The red line represents the f(y) = x function, the null expectation if no preference is shown: plant species above the red line are used more often than expected given their abundance, whereas those below the line are avoided
The height at which eggs are positioned above the ground increased with the height of the receiving plant (Fig. 5). A linear model showed a strong relationship between total plant height and height at which the egg is placed (main effect coefficient = 0.734 (S.E. 0.026), p < 0.001), with no improvement of the model when plant species is included as a random effect (AIC 1208 for the mixed model compared with 1206 for the simplest model).

Egg height plotted against height of the host plant across all 6 Fabaceae species used. No eggs were observed on T. repens
Patterns of egg positions on different plant structures appears to vary between the four most frequently used host plants (chi square p = 0.0004; see Table 1.) suggesting differences in choice of oviposition substrate between host plants.
Discussion
The only Fabaceae plants observed in our study sites were European species (Taggar et al. 2021), naturalized in Canada, that are known as host plants of P. icarus in its native range (Emmet and Heath 1989; Burghardt and Fiedler 1996; Janz et al. 2005; Robinson et al. 2010; Habel et al. 2016); all but one were used as oviposition sites in our study. The bottom-up facilitation hypothesis is therefore supported, suggesting that the presence of co-evolved host plants has facilitated the establishment of this herbivore in its invasion range (Engelkes and Mills 2013; Johnson et al. 2019; Mech et al. 2019).
Butterfly presence was best predicted by the abundance of two known host plants, namely L. corniculatus and T. pratense. Examination of oviposition showed preference for M. sativa over L. corniculatus, especially early in the season. The well-established association with L. corniculatus, observed in both the native (León-Cortés et al. 1999; Gutiérrez et al. 2001; Janz et al. 2005; Habel et al. 2016) and invasion (Rivest and Kharouba 2021) ranges, is, at least in the case of its novel Canadian range, linked to the abundance of this plant in disturbed sites with short vegetation rather than true behavioral preference. Indeed, Fig. 3 shows that L. corniculatus, T. pratense and M. lupulina are associated with short vegetation, likely increasing their use by the butterfly; in contrast, M. sativa is associated with tall vegetation and therefore might be less frequently used in the field despite being a preferred host. Furthermore, P. icarus oviposit more on plants with than without flowers, and often oviposit after nectaring, suggesting that flowering period could play a role in host selection (Janz et al. 2005). It must also be noted that work on P. icarus host preference has mostly been done in Western Europe, notably in the British isles, only part of a vast native range spanning Europe and parts of Asia, and that the provenance of the Canadian invaders is unknown.
L. corniculatus is abundant on roadsides and disturbed areas in Eastern Canada; it is very tolerant of dry, infertile and acidic soils and of salt contamination and is used in land reclamation (Turkinton and Franko 1980). All the plants used as hosts in this study are common in disturbed areas (Turkinton and Franko 1980), and our results support the view that this butterfly will expand its range in North America using roads as corridors (Al-Rewashdy et al. 2010), but remain limited to urban and disturbed settings with little effects on native ecosystems (Rivest and Kharouba 2021).
Our findings also support the role of vegetation structure in defining habitat quality for P. icarus in its invasion range. A similar preference for short vegetation has also been recorded in the British isles (León-Cortés et al. 1999) and in Germany (Habel et al. 2016), where this short-vegetation meadow biotope is typically found in low intensity agricultural land, such as traditionally grazed pastures. These plant communities are becoming increasingly rare and P. icarus is declining in Europe (Cowley et al. 1999; León‐Cortés et al. 1999; Maes and Van Dyck 2001). This suggests a discrepancy in biotope use between the invasion range, where the butterfly is concentrated in cities (GBIF Secretariat 2021; Rivest and Kharouba 2021) and the native range where it is not listed among common urban species (Dennis et al. 2017) and where mown urban sites dominated by short L. corniculatus are considered unsuitable (Gutiérrez et al. 2001).
The P. icarus thus uses different biotopes in the invasion than in the native range, but in both cases it is associated with the presence of Fabaceae plants, particularly L. corniculatus, and with short vegetation. These features are found in unimproved grasslands in the native range, but in the invasion range are provided by the novel plant communities found in cities. As per the resource-based habitat model (Dennis et al. 2003), it is the presence of specific resources, namely host plants and short vegetation, rather than that of particular plant communities, that promotes the establishment of this butterfly in its invasion range.
The preference observed for short vegetation lacks a mechanistic explanation. Possibilities include the role of vegetation structure for microclimate, for host-finding, for refuges for overwintering larvae, for roosting (Dover et al. 1997) as well as possible effects on top-down forces from predators or parasitoids, or on tending by ants (Dexheimer et al. 2021). Our results show that eggs are laid toward the top of plants, on different anatomical structures on different host species. Previous work in Sweden has shown another myrmecophilous lycaenid (Phengaris alcon) to oviposit preferentially at the top of plants surrounded by short vegetation, suggesting that host choice depends both on plant traits and vegetation structure (Valdés and Ehrlén 2018). The authors further conclude that plant-insect interactions are context dependent, because small-scale environmental factors, including physical structure of the habitat, influence both insect behavior and expression of plant traits, as well as behavior of other interaction partners like tending ants (Valdés and Ehrlén 2018).
It is not clear why Canadian cities should provide more suitable resource-based habitat than European ones, but it would be worth investigating the theory of niche opportunities which proposes that the an invasive species might face less biotic resistance in the form of competitors or natural enemies in its invasion range (Cadotte et al. 2017). Our results show that the P. icarus in the invasion range has re-established pre-existing relationships with host plants from its native range. Previous work shows how this species has forged a novel relationship with another interaction partner, a tending ant (Dexheimer et al. 2021). Indeed, the caterpillar is known to be myrmecophilous and is tended by a native North American ant (Lasius neoniger) in the Montreal area (Dexheimer et al. 2021). Adaptive values of these different relationships are unknown: it is not clear how larval growth, overwinter survival, predator/parasitoid pressure or protection by myrmecophily differ between the native and invasion range. All these relationships could be involved in defining niche opportunities in the novel range. It is also possible that given differences between Eastern Canada and Western Europe, for instance in the relationship between climate and photoperiod, P. icarus uses resources differently in these two places (Dennis et al. 2003). The high density of Canadian P. icarus populations, compared to the species’ decline in Europe, supports the idea that high quality resource-based habitat is found in the invasion range.
The role of cities in insect conservation is increasingly recognized (New 2015; Theodorou 2022), especially as many other habitats disappear. In Europe, unimproved grazed grasslands are decreasing as traditional extensive agriculture is abandoned, and common grassland butterflies, including P. icarus, are becoming rarer (Cowley et al. 1999; León-Cortés et al. 1999; Maes and Van Dyck 2001). Some butterflies find new habitats in cities, and thus cities can be important conservation resources for these species (Öckinger et al. 2009). Overall, cities tend to show lower overall butterfly diversity than less disturbed habitats, but some species, known as urban exploiters, do thrive (Öckinger et al. 2009; Theodorou 2022). P. icarus shows some of the traits (multivoltinism, phenological plasticity, feeding on ruderal plants) expected for an urban exploiter. Evidence suggests that European cities are not very good habitat for P. icarus (Hardy and Dennis 1999; Gutiérrez et al. 2001; Konvicka and Kadlec 2011; Dennis et al. 2017), but that Canadian cities might provide better quality resource-based habitats for this species. This raises the intriguing question of the role of populations in the invasion range for conservation of a species declining in its native range.
References
Agosta S (2006) On ecological fitting, plant-insect associations, herbivore host shifts, and host plant selection. Oikos 114:556–565
Aguirre-Gutiérrez J, WallisDeVries MF, Marshall L et al (2017) Butterflies show different functional and species diversity in relationship to vegetation structure and land use. Glob Ecol Biogeogr 26:1126–1137. https://doi.org/10.1111/geb.12622
Al-Rewashdy H, Larrivee M, Thomson A et al (2010) The Niche of an invasive species, Polyommatus icarus
Berg Ã, Ahrné K, Öckinger E et al (2013) Butterflies in semi-natural pastures and power-line corridors—effects of flower richness, management, and structural vegetation characteristics. Insect Conserv Divers 6:639–657. https://doi.org/10.1111/icad.12019
Burghardt F, Fiedler K (1996) The influence of diet on growth and secretion behaviour of myrmecophilous Polyommatus icarus caterpillars (Lepidoptera: Lycaenidae). Ecol Entomol 21:1–8
Cadotte MW, Yasui SLE, Livingstone S, MacIvor JS (2017) Are urban systems beneficial, detrimental, or indifferent for biological invasion? Biol Invasions 19:3489–3503. https://doi.org/10.1007/s10530-017-1586-y
Cowley MJR, Thomas CD, Thomas JA, Warren MS (1999) Flight areas of British butterflies: assessing species status and decline. Proc R Soc Lond Ser B Biol Sci 266:1587–1592
Dennis RL, Dapporto L, Dover JW (2014) Ten years of the resource-based habitat paradigm: the biotope-habitat issue and implications for conserving butterfly diversity. J Insect Biodivers 2:1–32
Dennis EB, Morgan BJ, Roy DB, Brereton TM (2017) Urban indicators for UK butterflies. Ecol Ind 76:184–193
Dennis RLH, Shreeve TG, Van Dyck H (2003) Towards a functional resource-based concept for habitat: a butterfly biology viewpoint. Oikos 102:417–426
Dexheimer E, Nascimento de Araújo H, Despland E (2021) Novel mutualistic interaction in introduced Polyommatus icarus larvae in Quebec. J Ent Soc Ontario 152:29–38
Dover JW, Sparks TH, Greatorex-Davies JN (1997) The importance of shelter for butterflies in open landscapes. J Insect Conserv 1:89–97. https://doi.org/10.1023/A:1018487127174
Emmet AM, Heath J (1989) The moths and butterflies of Britain and Ireland. Harley Books, England
Engelkes T, Mills NJ (2013) A fast-track for invasion: invasive plants promote the performance of an invasive herbivore. Biol Invasions 15:101–111
eButterfly (2022) eButterfly: an online database of butterfly distribution and abundance [web application]. Available: http://www.e-butterfly.org. Accessed 2 June 2022
GBIF Secretariat (2021) Polyommatus icarus (Rottemburg, 1775). In: GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei. Accessed via https://www.gbif.org/ on 17 Nov 2022
Gutiérrez D, León-Cortés JL, Menéndez R et al (2001) Metapopulations of four lepidopteran herbivores on a single host plant, Lotus corniculatus. Ecology 82:1371–1386. https://doi.org/10.1890/0012-9658(2001)082[1371:MOFLHO]2.0.CO;2
Habel JC, Teucher M, Ulrich W et al (2016) Drones for butterfly conservation: larval habitat assessment with an unmanned aerial vehicle. Landsc Ecol 31:2385–2395
Hall P (2007) The European common blue Polyommatus icarus new alien butterfly to Canada and North America. News Lepid. Soc. 49:111
Hardy PB, Dennis RL (1999) The impact of urban development on butterflies within a city region. Biodivers Conserv 8:1261–1279
Harvey JA, Bukovinszky T, van der Putten WH (2010) Interactions between invasive plants and insect herbivores: a plea for a multitrophic perspective. Biol Conserv 143:2251–2259
Hosmer D, Lemeshow S (2000) Applied logistic regression, 2nd ed. Wiley Online Books. https://onlinelibrary.wiley.com/doi/book/10.1002/0471722146
Janz N, Bergström A, Sjögren A (2005) The role of nectar sources for oviposition decisions of the common blue butterfly Polyommatus icarus. Oikos 109:535–538. https://doi.org/10.1111/j.0030-1299.2005.13817.x
Johnson LR, Breger B, Drummond F (2019) Novel plant–insect interactions in an urban environment: enemies, protectors, and pollinators of invasive knotweeds. Ecosphere 10:e02885. https://doi.org/10.1002/ecs2.2885
Konvicka M, Kadlec T (2011) How to increase the value of urban areas for butterfly conservation? A lesson from Prague nature reserves and parks. Eur J Entomol 108:219
León-Cortés JL, Cowley MJR, Thomas CD (1999) Detecting decline in a formerly widespread species: how common is the common blue butterfly Polyommatus icarus? Ecography 22:643–650. https://doi.org/10.1111/j.1600-0587.1999.tb00513.x
Maes D, Van Dyck H (2001) Butterfly diversity loss in Flanders (north Belgium): Europe’s worst case scenario? Biol Conserv 99:263–276. https://doi.org/10.1016/S0006-3207(00)00182-8
Mech AM, Thomas KA, Marsico TD et al (2019) Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol Evol 9:12216–12230
New TR (2015) Alien species in Urban environments. In: New TR (ed) Insect conservation and urban environments. Springer International Publishing, Cham, pp 87–101
Öckinger E, Dannestam Ã, Smith HG (2009) The importance of fragmentation and habitat quality of urban grasslands for butterfly diversity. Landsc Urban Plan 93:31–37. https://doi.org/10.1016/j.landurbplan.2009.05.021
Pollard E, Yates TJ (1994) Monitoring butterflies for ecology and conservation: the British butterfly monitoring scheme. Springer Science & Business Media
Pöyry J, Luoto M, Paukkunen J et al (2006) Different responses of plants and herbivore insects to a gradient of vegetation height: an indicator of the vertebrate grazing intensity and successional age. Oikos 115:401–412. https://doi.org/10.1111/j.2006.0030-1299.15126.x
Rivest SA, Kharouba HM (2021) Anthropogenic disturbance promotes the abundance of a newly introduced butterfly, the European common blue (Polyommatus icarus; Lepidoptera: Lycaenidae), in Canada. Can J Zool 99:642–652
Robinson GS, Ackery PR, Kitching IJ et al (2010) HOSTS—A Database of the World’s Lepidopteran Hostplants
Taggar AK, McGrath E, Despland E (2021)& Competition between a native and introduced pollinator in unmanaged urban meadows. Biol Invasions 1–9
Theodorou P (2022) The effects of urbanisation on ecological interactions. Curr Opin Insect Sci. https://doi.org/10.1016/j.cois.2022.100922
Turkington R, Franko GD (1980) THE BIOLOGY OF CANADIAN WEEDS.: 41. Lotus corniculatus L. Can J Plant Sci 60:965–979. https://doi.org/10.4141/cjps80-140
Valdés A, Ehrlén J (2018) Direct and plant trait-mediated effects of the local environmental context on butterfly oviposition patterns. Oikos 127:825–833. https://doi.org/10.1111/oik.04909
Vanreusel W, Van Dyck H (2007) When functional habitat does not match vegetation types: a resource-based approach to map butterfly habitat. Biol Conserv 135:202–211
Wisz MS, Pottier J, Kissling WD et al (2013) The role of biotic interactions in shaping distributions and realised assemblages of species: implications for species distribution modelling. Biol Rev 88:15–30
Acknowledgements
Thanks to Henrique Nascimento de Araújo, Sebastian Caucci and Natalie Dupont for assisting with fieldwork.
Funding
This study was funded by a Natural Sciences and Engineering Research Council (NSERC) Discovery Grant to E. Despland, a Mitacs Globalink Award to HNA, and an NSERC Undergraduate Student Research Award to ND.
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception and data analysis. Fieldwork was performed by Eric Dexheimer. The first draft of the manuscript was written by Eric Dexheimer and edited by Emma Despland. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Dexheimer, E., Despland, E. Newly introduced butterfly species’ urban habitat use driven by shorter vegetation and exotic plants. Biol Invasions 25, 1767–1777 (2023). https://doi.org/10.1007/s10530-023-03009-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-023-03009-3