
Abstract
Biological invasions by insects that vector plant pathogens have altered the composition of natural and urban forests. Thousand cankers disease is a new, recent example and is caused by the complex of walnut twig beetle, Pityophthorus juglandis, and the fungus, Geosmithia morbida, on susceptible hosts, notably some Juglans spp. and Pterocarya spp. Host colonization by P. juglandis may be particularly important for disease development, but the beetle’s host range is not known. In the United States and Italy, this insect has expanded its geographic range by colonizing naïve hosts. The objective of this study was to characterize limits to, and variation within, the host range of P. juglandis and infer the extent to which hosts might constrain the geographic distribution of the insect. We examined colonization and reproduction by P. juglandis in no-choice laboratory experiments with 11 Juglans spp., one Pterocarya sp., and two Carya spp. over 2 years and found that all but the Carya spp. were hosts. Reproduction was generally greater on Juglans californica, J. hindsii, and J. nigra, than on J. ailantifolia, J. cathayensis, J. cinerea, J. major, J. mandshurica, J. microcarpa, or J. regia. Escape of an insect vector into populations of evolutionary-naïve hosts can facilitate rapid range expansion by the pest and massive mortality to hosts. Multi-continental plantings of suitable species may facilitate geographic range expansion of P. juglandis and place other, native Juglans spp. at risk.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forest insects are important ecological disturbance agents (e.g., Bentz et al. 2010; Gandhi and Herms 2010; Kautz et al. 2017). Global movement of forest pests is of high concern to land managers and to the public given the landscape scale mortality that can result from establishment in naïve hosts (Anulewicz et al. 2008; Økland et al. 2011; Flø et al. 2014; Umeda et al. 2016). Mortality can be particularly severe when a forest insect vectors a plant pathogen to a susceptible host. Notable examples of transformative forest diseases in North America caused by insect-pathogen complexes comprised of at least one non-native species include beech bark disease (Ehrlich 1934), Dutch elm disease (Brasier 1991), and laurel wilt (Fraedrich et al. 2008). Range expansion by insects through colonization of naïve hosts can occur following climatic shifts (de la Giroday et al. 2012), human-mediated introductions (Hanula et al. 2008; Herms and McCullough 2014), and widespread use of non-native trees in urban or forested environments (Branco et al. 2015). Predicting which alien insects are likely to invade and where the risk to forests is greatest is a priority for federal land managers (Venette et al. 2010) because prevention is the least costly form of invasive-species management (e.g., Aukema et al. 2011; Koch et al. 2011; Yemshanov et al. 2012).
One emerging threat to forestry and agriculture is thousand cankers disease in the walnut family Juglandaceae (Kolařík et al. 2011; Tisserat et al. 2011; Seybold et al. 2013). This family is distributed across North and South America and Eurasia and contains species that are important in forest ecosystems, in cultivated nut production, and for high quality wood products (Aradhya et al. 2006; Newton and Fowler 2009). Thousand cankers disease is caused by an insect-pathogen complex. The walnut twig beetle, Pityophthorus juglandis Blackman, inoculates the phloem of hosts with a fungal pathogen, Geosmithia morbida Kolařík et al. (Kolařík et al. 2011). Males initiate colonization of a host when they leave their developmental (brood) host, locate a new host, bore through the outer bark and feed in the phloem, create a nuptial chamber, and attract females and other males via an aggregation pheromone (Wood 1982; Seybold et al. 2016). Host selection by males is influenced by several stimuli (visual, tactile, olfactory, and gustatory) from the potential host and ends with sustained feeding in the phloem (Wood 1982). One to two females join the male in the nuptial chamber, and after mating, elongate egg galleries and lay eggs individually along the gallery walls. Development time from egg through three instars to adult is approximately 12 weeks; two overlapping generations occur per year (Dallara et al. 2012; Faccoli et al. 2016). Adult offspring likely remain near the natal host or colonize a proximate new host to reproduce, as the flight capacity of adults is limited (Kees et al. 2017). Intensive feeding by P. juglandis adults and larvae and phloem necrosis from the fungal pathogen lead to thousand cankers disease on susceptible hosts (Tisserat et al. 2009). The activity and abundance of beetles in an area may be affected by the quality of host trees (Faccoli et al. 2015).
Pityophthorus juglandis is native to northern Mexico and the southwestern United States where its native host, Juglans major, occurs (Rugman-Jones et al. 2015). Pityophthorus juglandis has spread out of its native range over the last century and as of September 2015 occurs in 16 U.S. states (9 western; 7 eastern) and Italy (Montecchio and Faccoli 2014; Seybold et al. 2016). This insect may remain under the bark of cut wood from hosts, and untreated walnut wood with bark may be moved for firewood (Jacobi et al. 2012) or woodworking, for example (Newton and Fowler 2009). From field observations, host-switching has contributed to P. juglandis range expansions. This insect species is established in both adventive plantings and native stands of Juglans spp. and some Pterocarya spp. (Tisserat et al. 2009; Flint et al. 2010; Grant et al. 2011; Tisserat et al. 2011; Serdani et al. 2013; Montecchio et al. 2014; Yaghmour et al. 2014; Montecchio and Faccoli 2014; Hishinuma et al. 2016; Seybold et al. 2016), yet many species in the Juglandaceae occur in regions of the world where P. juglandis has not yet invaded (Aradhya et al. 2006). The ability of P. juglandis to feed and reproduce in these species is not known, but the susceptibility of eleven species of Juglandaceae to G. morbida has been investigated (Utley et al. 2013).
Juglans and Pterocarya are widely distributed genera, and their geographic distributions might reflect the potential range of P. juglandis worldwide. The objective of this study was to test Juglandaceae species for P. juglandis colonization and development to characterize limits to, and variation within, the insect’s potential host range. We examined the likelihood of male establishment after one introduction to potential host material in the laboratory under no-choice conditions and examined differences in reproduction among hosts.
Materials and methods
Colonization and offspring (i.e., brood) production were assessed in no-choice laboratory experiments across 11 Juglans spp., one Pterocarya sp., and two Carya spp. These species were collected from multiple locations across the United States (Table 1). Specific accession numbers for the plants are provided in Supplemental Appendix 1. Multiple cultivars of Juglans nigra were included as positive controls. Carya illinoensis and C. ovata were included as putative negative controls. Individual trees from germplasm repositories were selected to avoid clones or trees from the same seed source and to ensure a high degree of genetic and geographic diversity in our experimental material.
A tree species was defined as a host if adult offspring were produced (i.e., complete growth and development) (Hodkinson and Hughes 1982) and the number of female offspring met or exceeded the number of female parents (i.e., maternal replacement). The primary metric of host suitability was the mean total number of offspring produced per female parent in branch sections of each tree species.
Cut branches were used in assays instead of whole trees as host material for two reasons. First, there are Juglandaceae that either do not occur or are rare within the current geographic range of P. juglandis. Several known hosts and other potential hosts were screened simultaneously under uniform conditions. Second, a reliable method to study P. juglandis reproduction in cut branches has recently been developed (Hefty et al. 2016). Host effects have been detected in previous bark beetle assays by using cut plant material (Švihra and Volney 1983; Lee et al. 2008; Walter et al. 2010; Mayfield et al. 2013; McKee et al. 2013).
Insects
Naturally infested hybrid black walnut branch sections [Juglans hindsii × (J. nigra × J. hindsii/J. californica] from a commercial seed orchard in Sutter Co., CA, USA (39°03.681′N, 121°36.818′W, 19.2 m elevation) were shipped to a Biosafety Level-2 facility in St. Paul, MN in February, March, April, and May 2014 to provide a source of parent beetles for experiments conducted in 2014. Adult P. juglandis emerged from these cut branch sections between July and September 2014. For experiments conducted in 2015, parent beetles were sourced from infested Juglans californica branch sections from the Juglans collection of the USDA Agricultural Research Service (ARS) National Clonal Germplasm Repository (NCGR) in Winters, California (38°30′10.7″N 121°58′51.5″W) and shipped to St. Paul, MN in July, August, September, and October 2015. Adult P. juglandis emerged daily from those branch sections between September and October 2015. All branch sections, in 2014 and 2015, were held on a laboratory benchtop (20–22 °C, 30–50% RH, 14:10 L:D) in 3.8 L plastic jars (ULINE, Pleasant Prairie, WI) with a top of modified micro-mesh (“No-see-um” mesh; 96–112 holes per cm2; Quest Outfitters, Sarasota, FL) that allowed air exchange. Emerged beetles were collected daily, and the sexes were separated (Bright 1981) and held in sealed Petri dishes with moist Kimwipes (Kimberly Clark, Roswell, GA) for 2 d before introduction to branch sections.
Host range assays
When available, ten trees were sampled per species by removing branches that were approx. 3–5 cm in diameter. For some species, more than one branch was sampled from the same tree because 10 individual trees were not available (Table 1). The lowest replication occurred for C. ovata, which had four trees. Pityophthorus juglandis preferentially colonizes branches larger than 1.5 cm in the field (Seybold et al. 2016). All samples (i.e., branch sections) were placed in plastic bags to reduce desiccation and kept in a cooler. Material from Texas, New Mexico, Missouri, and California was sent via overnight courier to St. Paul, MN. Samples collected from Indiana and locally in Minnesota were driven to St. Paul within 3 d after collection. Upon arrival, branches were cut to 25.4 cm, dipped in paraffin wax to seal cut ends, and placed in cold storage (≈ 4 °C) until beetles were available.
Cut branches from California were held in a − 80 °C freezer for 24 h to kill subcortical insects because these samples originated from counties (Solano and Yolo) known to have P. juglandis. Once branch sections were thawed, they were dipped in 2–5% bleach solution and allowed to dry to inhibit fungal growth at beetle entry sites (see below). California branch sections were sealed with Tree Bandage (Forestry Suppliers, Inc., Jackson, MS) because melting paraffin wax was not permitted in the quarantine facility. For the remainder of the text, when a tree species is discussed the state from which the material originated is designated within parentheses.
Methods developed by Hefty et al. (2016) were followed to infest branch sections with P. juglandis. Surface area was calculated from the length and diameter of each branch section and entrance holes were drilled at the density of two holes/100 cm2 of surface area. Entrance holes were drilled at a 30° angle to allow easier access to phloem and to avoid direct insertion of beetles into the xylem. The holes were approximately 2 mm in diameter, extending no further than to the sapwood surface.
Male P. juglandis were placed on bark for 10–15 s to confirm normal ambulatory behavior. Males that did not walk were discarded. One male was placed in each drilled hole with a fine paint brush. Holes were covered with modelling clay (Craft Smart natural clay, Irving, TX) to prevent males from escaping per Hefty et al. (2016). Males were checked daily and replaced up to three times until signs of boring were visible (i.e., frass accumulation around the hole from feeding and excavation in the underlying phloem). The number of male introductions was recorded for each hole on each cut branch section. If the first male accepted the branch after one introduction, the event was recorded as a 1. If a second or third male was introduced to a hole due to inactivity or death, the event was recorded as a 0.
Males were introduced to branch sections by using a randomized block design of 10–12 host species per block per day. If there were not enough male parents for a full block in 1 day, then that day was skipped. In 2014, branch sections from California were received in late summer after ten other tree species were infested (early summer), so in this instance, material was infested in sets of ten cut branches as they arrived in St. Paul.
After males showed signs of boring, the first step of host selection, one female was introduced to each hole with a fine paint brush. Females were also tested for normal ambulatory behavior before experimentation. Holes were re-sealed with modelling clay until feeding was evident. The time from placement of a male or female beetle into a drilled hole to signs of boring varied from 1 to 5 days. These procedures resulted in colonization densities of two to six mating pairs per cut branch section. After all mating pairs were established on a cut branch, the branch was placed in a 3.8 L plastic jar with modified lid (as described above), and jars were placed in a growth chamber (21 °C, 50% RH, 14:10 L:D).
After 12 weeks, all adults that had emerged into the rearing jars were counted. Cut branches were transferred into separate 3.8 L freezer bags and placed in a refrigerator (≈ 3 °C) until branch sections could be sampled. Insect reproduction is temperature dependent, and reproduction by P. juglandis at this temperature is unlikely. Outer bark was removed carefully with a #22 blade X-ACTO knife (Elmer’s Products, Inc., Westerville, OH) to expose egg galleries and larval mines. Larvae, pupae, and adults from galleries were removed with a fine paint brush and tallied. The total number of parents introduced to each branch section was subtracted from the number of adults found in the jar and branch.
In 2015, the same host species were screened (Table 1), with a few modifications. Reduced availability of parent beetles limited protocol to one entrance hole/100 cm2 of surface area, which limited intraspecific competition beneath the bark (Hefty et al. 2016). Males were replaced up to five times to ensure that reproduction could take place at a lower colonization density. Tree species from California were included with other species in the randomized block design. The focus with the California material in 2015 was the putative ancestral host (J. major) and a susceptible host from the invaded range (J. californica; Table 1). In 2014, at the end of 12 weeks, we had observed that the branch sections appeared to dry out, so in 2015, we measured percent moisture with a moisture meter (approximately 1 cm pin length, Delmhorst J-2000, Towaco, NJ) when the first male was introduced and at the end of 12 weeks. The sexes in a random subsample of adults from each host were separated (n = 20) to determine sex ratio.
Data analyses
Generalized linear mixed effects models with a binomial data distribution were used to examine the effect of tree species on the probability of male establishment after one introduction event, where species was treated as a fixed effect and source tree was treated as a random effect to help account for instances where more than one branch was taken from a single tree. After progeny emerged, species were assigned a host status (i.e., host or non-host). Generalized linear mixed effects models also were used to examine the effect of host status on the probability of male activity after one introduction where host status was treated as a fixed effect and source tree was treated as a random effect. Data were analyzed in R v3.2.3 (R Core Team 2015) with the packages ‘lme4’ and ‘lmerTest’ to fit and evaluate mixed-effects models (Kuznetsova et al. 2013; Bates et al. 2015). Separate analyses were performed for tree species infested in 2014 and 2015.
Analyses of the effect of tree species on reproduction by P. juglandis only included species that could be considered hosts. Generalized linear models were used to determine the effect of host on immatures (larvae and pupae) per female, adult brood per female, and total brood per female. For each measure of reproduction, when two branches were collected per tree (rare), values were averaged to provide a single measure for the tree and avoid pseudoreplication. The potential effects of host species on reproduction per female were analyzed in SAS ver 9.4 (Proc GLM; SAS, Cary, NC) with Tukey’s HSD test for post hoc comparisons among means. In 2014, cut branches collected from California arrived later in the summer and were infested at a later time, so these data were analyzed separately from data collected from branch sections infested earlier in the summer. In 2015, data from all cut branches were analyzed together. Square root transformations were used to satisfy model assumptions (normality of errors and homoscedasticity of variances) for analyses of 2014 and 2015 reproduction data. For 2015 data, initial analyses in R indicated no effect of change in percent moisture (starting—final percent moisture) on total brood per female (F1,72 = 1.25, P = 0.27), adults per female (F1,72 = 1.66, P = 0.20), or immatures per female (F1,70.22 = 0.54, P = 0.46). There was also no effect of the interaction between host and change in percent moisture (F9,72 ≤ 2.82, P ≥ 0.52), so model results are reported without a term for change in percent moisture of the branch sections.
To determine if the proportion of male to female adult offspring varied among Juglans hosts, logistic ANOVA and a binomial data distribution was used in R.
Results
Male establishment
The probability of male establishment immediately after one introduction to a branch section did not vary across host species tested in 2014 (χ 216 = 23.24, P = 0.10, Fig. 1a) but did in 2015 (χ 211 = 27.75, P = 0.003, Fig. 1b). In general, we found that males were less likely to colonize material that would not support further brood development, although almost half of them would readily chew exposed phloem at first presentation (Fig. 1). This pattern was consistent across both years. In 2014, the likelihood of male establishment after one introduction was 77–94% for all Juglans spp. and P. stenoptera. Male establishment after one introduction was 69% for C. illinoensis (MO) and 64% for C. ovata (IN), even though no brood developed in these hosts (see below). In 2015, the likelihood of male establishment after one introduction ranged from 50 to 89% for Juglans spp., and was ≤ 50% for C. illinoensis (MO) and C. ovata (IN).

Likelihood (± SE) of male Pityophthorus juglandis establishment after one introduction in branch sections from species of Carya, Juglans, and Pterocarya in a 2014 and b 2015. The left-hand grouping distinguishes species later confirmed to be hosts from non-hosts. The number of test branch sections is noted at the bottom of each histogram bar in parentheses. State abbreviations (USA) designate where the trees were grown. Asterisk, the likelihood of male establishment after one introduction was significantly greater for hosts collectively than non-hosts in 2014 (χ 21 = 13.67, P < 0.001) and 2015 (χ 21 = 9.43, P = 0.002)
Brood production
For tree species assayed in early summer 2014, the total number of brood per female from P. juglandis varied by species (F8,71 = 8.53, P < 0.001; Table 2). Again, if the number of brood exceeded the number of parents in a tree species, we considered it to be a host. All tested Juglans species and P. stenoptera were hosts (Fig. 2), and the two Carya species were not hosts (Fig. 2; Table 3). Among hosts, we found that per capita reproduction varied nearly eightfold (Fig. 2), from 5.0 ± 2.4 total brood per female (mean ± SE) in J. microcarpa (TX) to 39.3 ± 6.2 in J. nigra (IN). We found that J. nigra (IN) and J. cinerea × ailantifolia (IN) had the greatest mean number of adult progeny per female (24.3 ± 4.9 and 20.8 ± 4.7, respectively). Among species that could be considered hosts, J. microcarpa from southwestern Texas had the lowest mean adult progeny per female (1.7 ± 1.2). No adult offspring developed in C. illinoensis or C. ovata. One early instar developed in C. illinoensis, which was successfully colonized by only one male and female.

Mean (+ SE) total offspring (= brood) of Pityophthorus juglandis per female, partitioned into mean numbers of adults and immatures (larvae and pupae) per female for branch sections of potential host species of Carya and Juglans collected in 2014. Means with different letters indicate significant differences at α = 0.05 for total combined brood. State abbreviations (USA) designate where the trees were grown
When we assayed additional species from California in late summer 2014, we again found differences in reproduction among hosts (F5,53 = 5.45, P < 0.001; Table 2). Mean total brood per female ranged from 3.71 ± 0.91 in P. stenoptera to 23.6 ± 4.48 in J. hindsii (Fig. 3). Pterocarya stenoptera and J. hindsii also produced the lowest (0.6 ± 0.4) and greatest (10.6 ± 2.6) mean number of adult progeny per female, respectively (Fig. 3).

Mean (+ SE) total offspring (= brood) of Pityophthorus juglandis per female, partitioned into mean numbers of adults and immatures (larvae and pupae) per female for branch sections of potential host species of Juglans and Pterocarya collected in California in 2014. Means with different letters indicate significant differences (α = 0.05) for total combined brood
For tree species assayed in 2015, again, strong species-level differences in reproduction emerged (F9,79 = 7.67, P < 0.001; Table 2). Among hosts, we found that suitability varied approximately ten-fold (Fig. 4). Mean total brood per female ranged from 4.3 ± 2.5 in J. microcarpa (NM) to 42.9 ± 6.0 in J. nigra (MO). Juglans nigra from Missouri and J. californica from northern California supported the highest mean adult progeny per female (14.5 ± 5.0 and 13.8 ± 4.4, respectively). Juglans microcarpa (NM) and J. ailantifolia (IN and MN) yielded the lowest mean adult progeny per female, 0.6 ± 0.4 and 0.8 ± 0.5. Similar to 2014, no adult offspring developed in C. illinoensis and C. ovata. In 2015, mean adult progeny per female was lower than in 2014 for all hosts except J. californica (CA) and J. nigra (MO). Juglans californica (CA) had a higher mean adult progeny per female in 2015 than in 2014 and J. nigra (MO) had a similar level of adult development in both years. In 2015, all Juglans spp. tested and P. stenoptera met the two conditions for host status; Carya spp. did not meet either condition (Table 3).

Mean (+SE) total offspring (= brood) of Pityophthorus juglandis per female, partitioned into mean numbers of adults and immatures (larvae and pupae) per female for branch sections of potential host species collected in 2015. Means with different letters indicate significant differences at α = 0.05 for total combined brood
In Juglans hosts, P. juglandis females produced approximately one male offspring to two female offspring (M:F ratio was 0.53 ± 0.02). Sex ratio did not vary by host (χ 212 = 18.83, P = 0.09).
Discussion
Successful, but varied, reproduction of P. juglandis in branch sections of P. stenoptera and all Juglans spp. tested suggest that host may mediate and limit the geographic range expansion of P. juglandis through native and adventive Juglans and Pterocarya plantings in the United States, Europe, and elsewhere. One species that is native to the eastern United States, J. nigra, and two species that are native to the western United States, J. californica, and J. hindsii, generally supported the greatest levels of reproduction. Results for J. nigra span the range of possible outcomes; in 2015, while the per capita rate of reproduction was greatest on cut branches from Missouri, it was among the least on cut branches from Indiana. For the western species, especially J. californica, these observations are consistent with high levels of thousand cankers disease infection and tree mortality witnessed at the USDA ARS NCGR collection (Solano Co., California) and concomitant decline of tree vigor observed in the native range (Seybold et al. 2016; Hishinuma 2017). Less suitable hosts in our experiments included native southwestern United States species (J. major and J. microcarpa), Eurasian species (J. regia), Asian butternuts (J. ailantifolia, J. mandshurica, and J. cathayensis), a second native eastern United States species (J. cinerea), and one of its hybrids (J. ailantifolia × cinerea). Less suitable hosts may reduce spread rates by producing fewer adults, whereas more suitable hosts, given appropriate environmental conditions, would lead to greater spread rates if the dispersal behavior of the insect is unaffected.
Because J. major in the southwestern United States and Mexico has been reported as the putative native host of P. juglandis (Wood and Bright 1992; Seybold et al. 2016), our observations of modest levels of reproduction in this host suggest a co-evolved resistance to P. juglandis (Rugman-Jones et al. 2015). Thousand cankers disease may be another emerging example where escape of an insect herbivore into populations of evolutionary-naïve hosts (e.g., J. nigra, J. californica, J. hindsii) can facilitate rapid range expansion and widespread mortality to hosts (Haack et al. 2010; Cullingham et al. 2011; Herms and McCullough 2014).
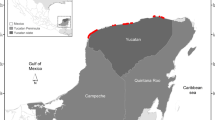
From our assays, P. juglandis appears to be a generalist within the genus Juglans and may be able to reproduce in other Juglans that we have not yet screened. We confirmed that some members of each of the four major taxonomic sections of genus Juglans are hosts, which strongly suggests the existence of other host species. The section Rhysocaryon is monophlyetic and comprised of J. major, J. microcarpa, J. nigra, J. californica, J. hindsii, and the somewhat more distantly related J. australis, J. boliviana, J. neotropica, J. guatemalensis, and J. olanchana; section Cardiocaryon includes J. mandshurica, J. ailantifolia, and J. cathayensis; section Trachycaryon only includes J. cinerea; and section Dioscaryon is comprised only of J. regia (Stanford et al. 2000). The phylogenetic relationships among species are generally supported by Orel et al. (2003). Examples of Juglans spp. not included in our assays have distributions in Central Asia (J. sigillata), Central America (J. mollis and J. olanchana), and South America (J. australis and J. neotropica), which suggests that these regions could be vulnerable to invasion by this insect (Fig. 5).

Potential distribution of Pityophthorus juglandis based on native ranges of Juglans hosts in North America, South America, and Eurasia (map reproduced with permission from Aradhya et al. 2006)
Additional hosts outside Juglans may occur. Our finding of P. stenoptera as a host is consistent with field observations of colonization and reproduction by P. juglandis in this and two other Pterocarya spp. (Hishinuma et al. 2016). We agree with Hishinuma et al. (2016) that the genera Alfaroa, Annamocarya, Carya, Cyclocarya, Engelhardia, and Platycarya (all Juglandaceae) merit attention in future host range assays. Cyclocarya falls within the subtribe Juglandinae, as do Juglans and Pterocarya (Manos and Stone 2001). Our finding that Carya is unlikely to be a host suggests that other genera within the subtribe Caryinae, such as Annamocarya, may not be hosts. Platycarya (tribe Platycaryeae) is basal within the subfamily Juglandoideae (Manos and Stone 2001), so knowledge of its host status would be useful to determine if suitability for P. juglandis is an ancestral or derived state within the clade. Alfaroa, Engelhardia, and Oreomunnea fall within the subfamily Engelhardioideae for which we have tested no representatives, so any projection of host usage by P. juglandis extrapolated from known hosts would be highly speculative.
Planting of susceptible species may be expanding the geographic area at risk from invasion by P. juglandis. Juglans regia is the most widely planted nut tree in the world (Stanford et al. 2000; Martinez et al. 2010). Pityophthorus juglandis has colonized trees in commercial J. regia orchards throughout California, USA (Flint et al. 2010; Yaghmour et al. 2014; Rugman-Jones et al. 2015; Seybold et al. 2016) and northern Italy (Montecchio et al. 2014; Faccoli et al. 2016). Pityophthorus juglandis has not been reported in the Southern Hemisphere, but commercial plantings of J. regia have been increasing in Australia (RIRDC 2017) and South America (Ramirez 2015), especially in Chile and Argentina (Martinez et al. 2010). In Chile, walnuts typically have been grown on rootstock from J. regia, J. hindsii, or their hybrids (Sotomayor and Castro 2004; S.S. Silva, personal communication). Planting non-native cultivars may also serve as a bridge to native walnut species. However, this risk may be low if the climate or other biotic factors are not favorable for P. juglandis to survive (Hefty et al. 2017).
Some Juglans species native to Eurasia and Asia are planted as ornamentals and small-scale nut trees in the United States. Juglans ailantifolia, for example, produces “heartnuts” and was introduced from Japan to the eastern United States as an ornamental tree (Manning 1978) in the late 19th century. Its introduced range overlaps with the native North American range of butternut, J. cinerea, and the two can hybridize (Zhao and Woeste 2011). There are no records of P. juglandis colonizing J. ailantifolia, J. cinerea, or hybrids of J. cinerea × J. ailantifolia in the eastern United States, though branch sections of J. cinerea baited with an attractant were colonized in Tennessee, USA (Hefty et al. 2016) and live trees of both species and perhaps the hybrid have been colonized with successful reproduction at the USDA ARS NCGR Juglans Collection (SJS, unpublished data). If introductions of P. juglandis continue to occur, however, these species could putatively support P. juglandis populations on the landscape in eastern North America or Asia (Table 3).
Before our study, it was unknown if P. juglandis could develop in Asian butternuts or North American hickories (Carya). Juglans ailantifolia, J. mandshurica, and J. cathayensis occur in portions of China, Taiwan, Japan, and Korea (Bai et al. 2016). Our data suggest that range expansion of P. juglandis via two Carya spp. is very unlikely. Pecan, C. illinoensis, is native to the southeastern United States and grown widely for nut production (Newton and Fowler 2009). Shagbark hickory, C. ovata, also native to eastern North America, is an important food source for wildlife (Grauke 2016). Although we have found that male P. juglandis will attempt to feed on Carya spp. under no-choice conditions and, in theory, that pathogen transmission could occur, Utley et al. (2013) reported that these tree species are not hosts for G. morbida. Wood (1982) describes bark beetle host selection as beginning with “the response to host stimuli prior to (and/or after) landing on the tree” and ending with “sustained feeding in the phloem.” Our assays skipped several steps in this process by placing the males and females directly into the phloem of the Carya spp. branch sections. Under these conditions, males appeared capable of completing the process of host selection at a relatively low frequency. It is not clear if they could complete the initial steps under natural conditions, and furthermore, given the low frequency of completion of host selection and near absence of larval development in our assays, populations would be unlikely to build if Carya species were the only Juglandaceae present.
The long-term environmental consequences of thousand cankers disease on the Juglandaceae resource remain unknown. Of all species tested, J. nigra is the most susceptible to G. morbida (Utley et al. 2013). In 2002, the estimated value of J. nigra growing stock in the eastern United States was over half a trillion dollars (Newton and Fowler 2009). Aggressive feeding by larvae and adults on J. nigra, combined with high susceptibility to the pathogen, and potential host stress induced by local growing conditions, all likely contributed to the rapid decline of J. nigra throughout the western United States (Tisserat et al. 2011). Despite this impact in the West, J. nigra in its native range does not appear to be as susceptible to the disease. Initial widespread mortality in the western United States prompted grave concern for urban forest managers and nut growers (Newton and Fowler 2009), but in the eastern United States where J. nigra is native, thousand cankers disease appears to progress at a slower rate. In some cases, trees have been able to recover, although recovery varies with site characteristics (Griffin 2015). More research is needed to determine host-selection characteristics of P. juglandis on live hosts, with an emphasis on varying vigor and pathogen interactions across hosts.
Specific mechanisms of host resistance to P. juglandis, such as induced and constitutive defenses, also require further study in this system. In our assays, J. microcarpa, J. ailantifolia, and P. stenoptera exhibited some of the lowest brood production per female, suggesting that potential host resistance to beetle reproduction could be exploited through development of hybrids with these species. The absence of any records of P. juglandis from native stands of J. microcarpa (Graves et al. 2011) coupled with our data on reproduction suggest that this species, in particular, may be particularly useful in a breeding program targeting resistance to P. juglandis.
The impacts of thousand cankers disease in the United States and Europe vary considerably, and the consequences of future spread by P. juglandis in South America, into Eurasia, or the Asian Far East and Australia are difficult to project, a difficulty common to many biological invasions (Venette et al. 2010). Certainly, the high incidence of human-mediated movement of P. juglandis in infested wood increases the risk of future introductions from the United States and Europe to the other continents (Hefty 2016). Our study supports the importance of monitoring in native and adventive stands of Juglans and Pterocarya to minimize future spread and impacts from P. juglandis.
References
Anulewicz AC, McCullough DG, Cappaert DL, Poland TM (2008) Host range of the emerald ash borer (Agrilus planipennis Fairmaire) (Coleoptera: Buprestidae) in North America: results of multiple-choice field experiments. Environ Entomol 37:230–241. https://doi.org/10.1603/0046-225x(2008)37[230:hrotea]2.0.co;2
Aradhya MK, Potter D, Simon CJ (2006) Cladistic biogeography of Juglans (Juglandaceae) based on chloroplast DNA intergenic spacer sequences. In: Motley TJ, Zerega N, Cross H (eds) Darwin’s harvest: new approaches to the origins, evolution and conservation of crops. Columbia University Press, New York, pp 143–170
Aukema JE, Leung B, Kovacs K et al (2011) Economic impacts of non-native forest insects in the continental United States. PLoS One 6:1–7. https://doi.org/10.1371/journal.pone.0024587
Bai WN, Wang WT, Zhang DY (2016) Phylogeographic breaks within Asian butternuts indicate the existence of a phytogeographic divide in East Asia. New Phytol 209:1757–1772. https://doi.org/10.1111/nph.13711
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bentz BJ, Régnière J, Fettig CJ et al (2010) Climate change and bark beetles of the western US and Canada: direct and indirect effects. BioScience 60:602–613
Branco M, Brockerhoff EG, Castagneyrol B et al (2015) Host range expansion of native insects to exotic trees increases with area of introduction and the presence of congeneric native trees. J Appl Ecol 52:69–77. https://doi.org/10.1111/1365-2664.12362
Brasier CM (1991) Ophiostoma novo-ulmi sp. nov., caustive agent of current Dutch elm disease pandemics. Mycopathlogia 115:151–161
Bright DE (1981) Taxonomic monograph of the genus Pityophthorus Eichhoff in North and Central America (Coleoptera: Scolytidae). Mem Entomol Soc Canada 113:1–378
Cullingham CI, Cooke JEK, Dang S et al (2011) Mountain pine beetle host-range expansion threatens the boreal forest. Mol Ecol 20:2157–2171. https://doi.org/10.1111/j.1365-94x.2011.05086.x
Dallara PL, Flint ML, Seybold SJ (2012) An analysis of the larval instars of the walnut twig beetle, Pityophthorus juglandis Blackman (Coleoptera: Scolytidae), in northern California black walnut, Juglans hindsii, and a new host record for Hylocurus hirtellus. Pan-Pac Entomol 88:248–266. https://doi.org/10.3956/2012-16.1
de la Giroday HMC, Carroll AL, Aukema BH (2012) Breach of the northern Rocky Mountain geoclimatic barrier: initiation of range expansion by the mountain pine beetle. J Biogeogr 39:1112–1123. https://doi.org/10.1111/j.1365-2699.2011.02673.x
Ehrlich J (1934) The beech bark disease: a Nectria disease of Fagus, following Cryptococcus fagi (Baer.). Can J For Res 10:593–692
Faccoli M, Favaro R, Concheri G et al (2015) Tree colonization by the Asian longhorn beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae): effect of habitat and tree suitability. Insect Sci 2:288–296. https://doi.org/10.1111/1744-7917.12192
Faccoli M, Simonato M, Rassati D (2016) Life history and geographical distribution of the walnut twig beetle, Pityophthorus juglandis (Coleoptera: Scolytinae), in southern Europe. J Appl Entomol 140:697–705. https://doi.org/10.1111/jen.12299
Flint ML, Graves AD, Seybold SJ (2010) Thousand cankers disease of walnuts spreads in California. CAPCA Advisor Mag 8:36–39. http://entomology.ucdavis.edu/files/201360.pdf. Accessed 1 Sept 2016
Flø D, Krokene P, Økland B (2014) Importing deciduous wood chips from North America to northern Europe—the risk of introducing bark- and wood-boring insects. Scand J For Res 29:77–89. https://doi.org/10.1080/02827581.2013.863380
Fraedrich SW, Harrington TC, Rabaglia RJ et al (2008) A fungal symbiont of the redbay ambrosia beetle causes a lethal wilt in redbay and other Lauraceae in the southeastern United States. Plant Dis 92:215–224. https://doi.org/10.1094/pdis-92-2-0215
Gandhi KJK, Herms DA (2010) Direct and indirect effects of alien insect herbivores on ecological processes and interactions in forests of eastern North America. Biol Invasions 12:389–405. https://doi.org/10.1007/s10530-009-9627-9
Grant JF, Windham MT, Haun WG et al (2011) Initial assessment of thousand cankers disease on black walnut, Juglans nigra, in eastern Tennessee. Forests 2:741–748. https://doi.org/10.3390/f2030741
Grauke LJ (2016) Hickories: Carya ovata. http://aggie-horticulture.tamu.edu/carya/species/ovata/ovata.htm. Accessed 5 July 2016
Graves AD, Coleman TW, Seybold SJ (2011) Monitoring walnut health and decline in response to thousand cankers disease and infestation by the walnut twig beetle, Pityophthorus juglandis, in southern California and New Mexico. Year one Progress Report for USDA Forest Service Forest Health Monitoring Project INT-EM-B-11-03 http://fhm.fs.fed.us/em/funded/12/INT-EM-B-11-03.pdf. Accessed 24 Mar 2016
Griffin GJ (2015) Status of thousand cankers disease on eastern black walnut in the eastern United States at two locations over 3 years. For Pathol 45:203–214. https://doi.org/10.1111/efp.12154
Haack R, Hérard F, Sun J, Turgeon JJ (2010) Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: a worldwide perspective. Annu Rev Entomol 55:521–546. https://doi.org/10.1146/annurev-ento-112408-085427
Hanula JL, Mayfield AE, Fraedrich SW, Rabaglia RJ (2008) Biology and host associations of redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae), exotic vector of laurel wilt killing redbay trees in the southeastern United States. J Econ Entomol 101:1276–1286. https://doi.org/10.1603/0022-0493(2008)101[1276:bahaor]2.0.co;2
Hefty AR (2016) Risk of invasion by walnut twig beetle throughout eastern North America. Dissertation, University of Minnesota
Hefty AR, Coggeshall MV, Aukema BH et al (2016) Reproduction of walnut twig beetle in black walnut and butternut. HortTechnology 26:727–734. https://doi.org/10.21273/horttech03427-16
Hefty AR, Seybold SJ, Aukema BH, Venette RC (2017) Cold tolerance of Pityophthorus juglandis (Coleoptera: Scolytidae) from northern California. Environ Entomol 46:967–977. https://doi.org/10.1093/ee/nvx090
Herms D, McCullough DG (2014) Emerald ash borer invasion of North America: history, biology, ecology, impacts, and management. Annu Rev Entomol 59:13–30. https://doi.org/10.1146/annurev-ento-011613-162051
Hishinuma SM (2017) Interactions among the walnut twig beetle, Pityophthorus juglandis, the pathogenic fungus, Geosmithia morbida, and host species in thousand cankers disease in California. Dissertation, University of California, Davis
Hishinuma SM, Dallara PL, Yaghmour M et al (2016) Wingnut (Juglandaceae) as a new generic host for Pityophthorus juglandis (Coleoptera: Curculionidae) and the thousand cankers disease pathogen, Geosmithia morbida (Ascomycota: Hypocreales). Can Entomol 148:83–91. https://doi.org/10.4039/tce.2015.37
Hodkinson ID, Hughes MK (1982) Insect herbivory. Chapman and Hall, London
Jacobi WR, Hardin JG, Goodrich BA, Cleaver CM (2012) Retail firewood can transport live tree pests. J Econ Entomol 105:1645–1658. https://doi.org/10.1603/ec12069
Kautz M, Meddens AJH, Hall RJ, Arneth A (2017) Biotic disturbances in Northern Hemisphere forests—a synthesis of recent data, uncertainities, and implications for forest monitoring and modeling. Glob Ecol Biogeogr 26:533–552
Kees AM, Hefty AR, Venette RC et al (2017) Flight capacity of the walnut twig beetle (Coleoptera: Scolytidae) on a laboratory flight mill. Environ Entomol 46:633–641. https://doi.org/10.1093/ee/nvx055
Koch FH, Yemshanov D, Colunga-Garcia M et al (2011) Potential establishment of alien-invasive forest insect species in the United States: where and how many? Biol Invasions 13:969–985. https://doi.org/10.1007/s10530-010-9883-8
Kolařík M, Freeland E, Utley C, Tisserat N (2011) Geosmithia morbida sp nov., a new phytopathogenic species living in symbiosis with the walnut twig beetle (Pityophthorus juglandis) on Juglans in USA. Mycologia 103:325–332. https://doi.org/10.3852/10-124
Kuznetsova A, Brockhoff B, Christensen HB (2013) lmerTest: tests for random and fixed effects for linear mixed effect models (lmer objects of lme4 package). R package version 2.0-0. http://CRAN.R-project.org/package=lmerTest
Lee JC, Flint ML, Seybold SJ (2008) Suitability of pines and other conifers as hosts for the invasive Mediterranean pine engraver (Coleoptera: Scolytidae) in North America. J Econ Entomol 101:829–837. https://doi.org/10.1603/0022-0493(2008)101[829:sopaoc]2.0.co;2
Manning WE (1978) The classification within the Juglandaceae. Ann Missouri Bot Gard 65:1058–1087
Manos PS, Stone DE (2001) Evolution, phylogeny, and systematics of the Juglandaceae. Ann of the Missouri Bot Gard 88:231–269
Martinez ML, Labuckas DO, Lamarque AL, Maestri DM (2010) Walnut (Juglans regia L.): genetic resources, chemistry, by-products. J Sci Food Agric 90:1959–1967
Mayfield AE, Mackenzie M, Cannon PG et al (2013) Suitability of California bay laurel and other species as hosts for the non-native redbay ambrosia beetle and granulate ambrosia beetle. Agric For Entomol 15:227–235. https://doi.org/10.1111/afe.12009
McKee FR, Huber DPW, Aukema BH (2013) Comparisons of mountain pine beetle (Dendroctonus ponderosae Hopkins) reproduction within a novel and traditional host: effects of insect natal history, colonized host species and competitors. Agric For Entomol 15:310–320. https://doi.org/10.1111/afe.12019
Montecchio L, Faccoli M (2014) First record of thousand cankers disease Geosmithia morbida and walnut twig beetle Pityophthorus juglandis on Juglans nigra in Europe. Plant Dis 98:696. https://doi.org/10.1094/pdis-10-13-1027-pdn
Montecchio L, Fanchin G, Simonato M, Faccoli M (2014) First record of thousand cankers disease fungal pathogen Geosmithia morbida and walnut twig beetle Pityophthorus juglandis on Juglans regia in Europe. Plant Dis 98:1445
Newton L, Fowler G (2009) Pathway Assessment: Geosmithia sp. and Pityophthorus juglandis Blackman movement from the western into the eastern United States. United States Department of Agriculture Animal and Plant Health Inspection Service, 1-50, Washington, D.C., Rev. 1: 10.19.2009. http://mda.mo.gov/plants/pdf/tc_pathwayanalysis.pdf. 24 April 2016
Økland B, Erbilgin N, Skarpaas O et al (2011) Inter-species interactions and ecosystem effects of non-indigenous invasive and native tree-killing bark beetles. Biol Invasions 13:1151–1164. https://doi.org/10.1007/s10530-011-9957-2
Orel G, Marchant AD, McLeod JA, Richards GD (2003) Characterization of 11 Juglandaceae genotypes based on morphology, cpDNA, and RAPD. Hortscience 38:1178–1183
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramirez N (2015) Chile’s tree nuts annual report 2015. USDA Foreign Agricultural Service. GAIN Report Number: CI1523
RIRDC [Rural Industries Research and Development Corporation] (2017) Walnuts. Farmdiversity. http://www.farmdiversity.com.au/Plant/0a7311a0-bdba-43ff-bbb5-a43600d1f1c8
Rugman-Jones PF, Seybold SJ, Graves AD, Stouthamer R (2015) Phylogeography of the walnut twig beetle, Pityophthorus juglandis, the vector of thousand cankers disease in North American walnut trees. PLoS ONE 10:1–28. https://doi.org/10.1371/journal.pone.0118264
Serdani M, Vlach JJ, Wallis KL et al (2013) First report of Geosmithia morbida and Pityophthorus juglandis causing thousand cankers disease in butternut. Plant Heal Prog. https://doi.org/10.1094/php-2009-0811-01-rs.4
Seybold SJ, Haugen D, O’Brien J, Graves AD (2013) Thousand cankers disease. USDA Forest Service, Northeastern Area State and Private Forestry Pest Alert, NA-PR-02-10, 2 pp. http://www.na.fs.fed.us/pubs/detail.cfm?id=5225. Accessed 24 Mar 2016
Seybold SJ, Penrose RL, Graves AD (2016) Invasive bark and ambrosia beetles in California Mediterranean forest systems. In: Paine TD, Lieutier F (eds) Insects and diseases of Mediterranean forest systems. Springer International, Cham, pp 583–662
Sotomayor C, Castro J (2004) Rootstocks used for fruit crops in Chile: an overview. Acta Hortic 658:287–291
Stanford AM, Harden R, Parks CR (2000) Phylogeny and biogeography of Juglans (Juglandaceae) based on matK and ITS sequence data. Am J Bot 87:872–882
Švihra P, Volney WJA (1983) Effect of English, Siberian, and Chinese elms on the attack behavior and brood survival of Scolytus multistriatus (Coleoptera: Scolytidae). Can Entomol 115:513–522
Tisserat N, Cranshaw W, Leatherman D et al (2009) Black walnut mortality in Colorado caused by the walnut twig beetle and thousand cankers disease. Plant Health Prog. https://doi.org/10.1094/php-2009-0811-01-rs
Tisserat N, Cranshaw W, Putnam M et al (2011) Thousand cankers disease is widespread in black walnut in the western United States. Plant Health Prog. https://doi.org/10.1094/php-2011-0630-01-br.black
Umeda C, Eskalen A, Paine TD (2016) Polyphagous shot hole borer and Fusarium dieback in California. In: Paine TD, Lieutier F (eds) Insects and diseases of Mediterranean forest systems. Springer International, Cham, pp 757–767
Utley C, Nguyen T, Roubtsova T et al (2013) Susceptibility of walnut and hickory species to Geosmithia morbida. Plant Dis 97:601–607. https://doi.org/10.1094/pdis-07-12-0636-re
Venette RC, Kriticos DJ, Magarey R et al (2010) Pest risk maps for invasive alien species: a roadmap for improvement. BioScience 60:349–362
Walter AJ, Venette RC, Kells SA (2010) Acceptance and suitability of novel trees for Orthotomicus erosus, an exotic bark beetle in North America. Biol Invasions 12:1133–1144. https://doi.org/10.1007/s10530-009-9531-3
Wood DL (1982) The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annu Rev Entomol 27:411–446
Wood SL, Bright DE Jr (1992) A catalog of Scolytidae and Platypodidae (Coleoptera), part 2: Taxonomic Index, Volume B. Great Basin Nat 13:1553
Yaghmour MA, Nguyen TL, Roubtsova TV et al (2014) First report of Geosmithia morbida on English walnut and its Paradox rootstock in California. Plant Dis 98:1441
Yemshanov D, Koch FH, Ducey M, Koehler K (2012) Trade-associated pathways of alien forest insect entries in Canada. Biol Invasions 14:797–812. https://doi.org/10.1007/s10530-011-0117-5
Zhao P, Woeste KE (2011) DNA markers identify hybrids between butternut (Juglans cinerea L.) and Japanese walnut (Juglans ailantifolia Carr.). Tree Genet Genomes 7:511–533. https://doi.org/10.1007/s11295-010-0352-4
Acknowledgements
We thank Charles Leslie, Department of Plant Sciences, University of California, Davis, CA; John Preece, USDA ARS NCGR, Davis, CA; Andrew D. Graves and James Jacobs, USDA Forest Service Forest Health Protection, Albuquerque, NM; Minnette Marr, Lady Bird Johnson Wildflower Center, Austin, TX; and the University of Minnesota Landscape Arboretum, Chaska, MN for assistance with procuring host material for testing. We thank Cliff Beumel, Director of Product Development, Sierra Gold Nurseries, Yuba City, CA for access to a commercial walnut seed orchard for collection of walnut twig beetles; J. Preece for access to the USDA ARS NCGR for the same purpose; and University of Minnesota undergraduate research assistants, Joe Pohnan and Collin Smith. We also thank Carlos H. Crisosto and Sebastian S. Silva (both Department of Plant Sciences, University of California, Davis) for providing information on walnut production in South America. We greatly appreciate the staff at the MDA/MAES Plant Growth Facility for support with biocontainment. Funding was provided by the NSF-IGERT Risk Analysis of Introduced Species and Genotypes program at the University of Minnesota (DGE-0653827) and a USDA-Forest Service Special Technology Development Program grant (R2-2012-01) that was administered by Jeffrey Witcosky and Stephanie Stephens, USDA Forest Service, Lakewood, CO. Participation of MVC and SJS in this project was coordinated and supported by USDA National Institute of Food and Agriculture, Specialty Crops Research Initiative Program Project (11684658), “Development of Disease-resistant Walnut Rootstocks: Integration of Conventional and Genomic Approaches.”
Author information
Authors and Affiliations
Contributions
ARH, RCV, BHA, MVC, JRM, and SJS all contributed to the experimental design of host assays. ARH performed all assays. ARH, BHA, and RCV analyzed the data. SJS, MVC, and JRM provided host material. SJS provided insects. ARH, BHA, RCV, and SJS wrote the manuscript; other authors provided editorial advice.
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hefty, A.R., Aukema, B.H., Venette, R.C. et al. Reproduction and potential range expansion of walnut twig beetle across the Juglandaceae. Biol Invasions 20, 2141–2155 (2018). https://doi.org/10.1007/s10530-018-1692-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-018-1692-5