
Abstract
Objective
The effects of monosaccharide constituents of lignocellulosic materials on exopolysaccharide (EPS) production by Mesorhizobium sp. Semia 816 were studied.
Results
According to the results, by using sugars commonly found in lignocellulosic biomass as carbon sources (glucose, arabinose and xylose), no significant differences were observed in the production of EPS, reaching 3.39 g/L, 3.33 g/L and 3.27 g/L, respectively. Differences were observed in monosaccharide composition, mainly in relation to rhamnose and glucuronic acid contents (1.8 times higher when arabinose was compared with xylose). However, the biopolymers showed no differences in relation to rheological properties, with EPS aqueous-based suspensions (1.0% w/v) presenting pseudoplastic behavior, and a slight difference in degradation temperatures. Using soybean hulls hydrolysate as carbon source, slightly higher values were obtained (3.93 g/L).
Conclusion
The results indicate the potential of the use of lignocellulosic hydrolysates containing these sugars as a source of carbon in the cultivation of Mesorhizobium sp. Semia 816 for the production of EPS with potential industrial applications.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Rhizobia are Gram-negative bacteria capable of establishing a nitrogen fixation symbiosis with leguminous plants (Senthil Kumar et al. 2017). These diazotrophic bacteria synthetize extracellular polymeric substances (EPS), consisting of polysaccharides, as well as proteins, nucleic acids, lipids, and other compounds such as humic acids. (Wingender et al. 1999). The secretion of EPS by rhizobia is associated with the invasion process and bacteroid and nodule development, as well as a response to environmental stress. They have fundamental ecological functions, such as the cohesion of microorganisms and adhesion of biofilms to surfaces and protection against abiotic and biotic factors, i.e., temperature, pH and salinity (Senthil Kumar et al. 2017; Costa et al. 2018).
The polysaccharides are the most studied component of EPS produced by rhizobia. These high molecular weight microbial polymers show diversity in terms of their chemical structures, implying different properties (Senthil Kumar et al. 2017). Current studies are focused on the discovery of new biopolymers of commercial interest and the optimization of industrial processes. In order to reduce production costs, alternative carbon sources from industrial byproducts and wastes have been used for the production of EPS, such as rice bran hydrolysate (Saranya Devi et al. 2012), crude glycerol (Oliveira et al. 2018), soy molasses (Oliveira et al. 2020) and whey (Zhou et al. 2014).
In this context, lignocellulosic biomass is a promising alternative for application in biotechnological processes because it is abundant, low in cost and renewable. The three-dimensional lignocellulosic complex is formed by three macromolecules, cellulose, hemicellulose, and lignin, that need to be converted into assimilable carbon sources for microbial conversion. While cellulose is a homopolymer of glucose, the hemicellulosic portion consists of a mixture of heteropolysaccharides containing hexoses and pentoses (Alfenore and Molina-Jouve 2016). However, there is little information about the capacity of rhizobial species to use mainly pentoses for EPS production and the impact on their properties.
Therefore, the aim of this work was to evaluate the ability of Mesorhizobium sp. Semia 816 to assimilate and convert some monosaccharide components of lignocellulosic residues into EPS, as well as to characterize the structural components and verify some properties in order to establish potential technological applications. In addition, soybean hulls hydrolysate was evaluated as an alternative carbon source in the cultivation.
Materials and methods
Soybean hulls hydrolysate
Soybean hulls were kindly provided by Bianchini S/A (Rio Grande, RS, Brazil). They were ground, sieved (1 mm diameter) and treated in autoclave (121 °C, 15 min) according to Ibbett et al. (2011), with modifications. The hydrolysis was performed at pH 5.3 (50 mM citrate buffer), 50 °C for 72 h, using Celluclast® 1.5L (20 FPU/g), based on previous assays (unpublished data).
Microorganism
Mesorhizobium sp. Semia 816 was provided by the Diagnostic and Agricultural Research Department (DDPA) of the Secretariat of Agriculture, Livestock and Rural Development (SEAP) of Rio Grande do Sul State (Porto Alegre, Brazil).
Reactivation of bacterial cells and preparation of the inoculum
The microorganism was reactivated at 30 °C for 48 h in YMA (Yeast Mannitol) agar. For the inoculum preparation, a tube with the reactivated microbial culture was scraped with 10 mL of 0.1% w/v peptone diluent to remove the cells. The suspension was then transferred to an Erlenmeyer flask containing 90 mL of YMA broth with pH adjusted to 7.0 before sterilization (Duta et al. 2006). The flask was kept in a rotary shaker at 30 °C and 200 rpm until an optical density (OD600) of 0.8 (Oliveira et al. 2020).
Cultivation in shake flasks
The cultivations were carried out in 500 mL Erlenmeyer flasks containing 90 mL of media adapted from Duta et al. (2006) with the following composition (g/L): carbon source (10); K2HPO4 (0.1); KH2PO4 (0.4); MgSO4·7H2O (0.2); NaCl (0.1); yeast extract (0.4); MnCl2·7H2O (0.12); CaCl2·2H2O (0.15). The broth was adjusted to pH 7.0 prior to sterilization.
The medium, with 10% v/v inoculum added, was incubated at 30 °C and 200 rpm in a rotary shaker for 96 h. Aliquots were centrifuged at 13,000×g for 30 min at 4 °C. pH and the substrate concentration were determined in the supernatant. The pellet was used to determine the biomass. At the end of the cultivation, the content of each flask was used for EPS recovery and quantification.
Analytical determinations
Biomass
The pellet was washed with distilled water, centrifuged again under the same conditions and resuspended in an appropriate volume. OD600 was measured and converted to cell dry weight (g/L) using a calibration curve for the microorganism.
Reducing sugars
The determination of reducing sugars was performed using the 3,5-dinitrosalicylic acid (DNS) spectrophotometric method (Miller 1959). Calibration curves using glucose, xylose or arabinose as standards were previously prepared.
Carbohydrates
The quantification of carbohydrates in hydrolysate was performed by High Performance Liquid Chromatography. A HPLC system (Waters, e2695, United States). equipped with an Aminex HPX-87H column (300 mm length × 7.8 mm internal diameter, Bio-Rad, United States) and a RID-10A refractive index detector. Mobile phase was composed by 0.05 M H2SO4 in ultrapure water. The analysis was performed in isocratic operation at 60 °C and 0.5 mL/min (30 min). The detector temperature was 30 °C. The sugars used as standards were cellobiose, xylose, arabinose and glucose.
EPS recovery
After cultivation, the medium was centrifugated at 13,000×g for 30 min at 4 °C for cell removal. For EPS recovery from the supernatant, ethanol 96% analytical grade was added in the proportion of 3:1 v/v (ethanol/supernatant). This suspension was stored at 4 °C for 24 h (Oliveira et al. 2020). The precipitate was dried at 50 °C until constant weight, quantified by gravimetry and the value related to the sample volume.
EPS characterization
The EPS samples were solubilized in ultrapure water and dialyzed (molecular weight cut-off 12 kDa) for 72 h at 5 °C for removal of impurities, and then frozen (− 80 °C) and lyophilized.
FT-IR
The determination of the functional groups of the EPS was performed by infrared spectrometer (Shimadzu, model IR Prestige-21, Japan). Spectra were obtained in the region of 4000–400 cm−1.
Monosaccharide composition by HPLC
Hydrolyzed EPS and monosaccharide standards (1 mg) were derivatized with 1-phenyl-3-methyl-5-pyrazolone (PMP), according to the protocol described by Castellane et al. (2014).
A Shimadzu-10A High Performance Liquid Chromatography System (Japan), which includes a SPD-M10Avp photodiode array UV–VIS detector (245 nm), a CTO-10Avp chromatography oven (30 °C) and a Restek-C18 column (150 × 4.6 mm, 5 μm) were used. The injected volume was 10 µL, using a flow rate of 0.5 mL/min. The mobile phase consisted of buffers A and B, 100 mM of ammonium acetate (pH 5.5) with 10% and 25% acetonitrile, respectively. For the calibration curve, glucose, mannose, rhamnose, galactose, glucuronic acid and galacturonic acid were used as standards (Castellane et al. 2014).
Rheological behavior
The freeze-dried EPS samples were suspended in ultrapure water (1% w/v), homogenized using a vortex mixer for 5 min and heated at 60 °C for 40 min. Samples were stored at 25 °C for 24 h to ensure complete hydration. Flow curves at 25 °C were obtained using a cone-plate Brookfield rotational rheometer (model DV-III, Brookfield Engineering Laboratories Inc., United States) equipped with a CP51 cone spindle (1.565° angle and 1.2 cm radius) and an adapter for small volumes (0.5 mL). The flow curves were obtained ranging the shear rate (\(\dot{\gamma }\)) between 0 and 100 s−1.
The adjustment of the experimental data was performed using the Herschel–Bulkley model, according to Eq. 1, to determine the rheological parameters. The apparent viscosity was determined by adjusting the data according to Eq. 2.
where σ0 is the yield stress (Pa), σ is the shear stress (Pa), \(\dot{\gamma }\) is the shear rate (s−1), η is the apparent viscosity (Pa s), k is the consistency index parameter (Pa sn) and n the flow behavior index (dimensionless).
Thermal analysis
DSC curves were obtained by a differential scanning calorimeter (DSC-60, Shimadzu, Japan), under a dynamic nitrogen atmosphere at 50 mL/min. The initial temperature was 50 °C, which was raised to 250 °C at 10 °C/min. TGA curves were obtained by a thermogravimetric analyzer (TGA-60, Shimadzu, Japan), under a nitrogen atmosphere at 30 mL/min, increasing the temperature to 500 °C at 10 °C/min.
Statistical analysis
All experiments as well as analytical determinations were performed in triplicate. The Tukey test was used at a 95% confidence interval (p < 0.05) to verify the existence of significant differences using Statistica 5.0 software (StatSoft Inc., USA).
Results and discussion
Cultivation parameters
Figure 1 shows biomass, pH and reducing sugars during Mesorhizobium sp. Semia 816 cultivation using xylose, glucose and arabinose.

Biomass (a), pH (b) and reducing sugars (c) during the cultivation of Mesorhizobium sp. Semia 816 using different carbon sources. (open circle) Xylose; (filled square) glucose; (open triangle) arabinose
The biomass profile for the three different carbon sources shows a typical increase over time (Fig. 1a). At 96 h cultivation, the biomass concentration reached the maximum values, with significant differences (p < 0.05) between the carbon sources (Table 1). In the cultivation of Rhizobium tropici Semia 4080, maximum biomass concentration was observed in the range of 0.77–1.44 g/L according to the carbon source (Castellane et al. 2017). In the work of Moretto et al. (2015), biomass production ranged from 0.76 to 3.88 g/L in the cultivation of Rhizobium isolates with sucrose.
In relation to EPS production, significant differences were not observed between the carbon sources (Table 1). In comparison with other diazotrophic bacteria, the results were relevant. Rhizobium sp. LBMP-C04 (6.63 g/L), followed by R. tropici LBMP-C01 (3.48 g/L), Ensifer sp. LBMP-C03 (2.86 g/L) and E. meliloti LBMP-C02 (2.75 g/L) appeared to be good producers of EPS when sucrose was used as the carbon source (Moretto et al. 2015).
Regarding the pH (Fig. 1b), a tendency towards acidification of the medium was observed. This fact can be associated with the preferential utilization of sugars that results in the formation of byproducts of total or partial digestion of the substrates as well as the excretion of acidic substances from rhizobia (Tan and Broughton 1981). This typical behavior was also observed by Jida and Assefa (2012) for Mesorhizobium isolates.
As shown in Table 1, the maximum specific growth rate (µmax), the maximum biomass concentration (Xmax) and the substrate to biomass conversion factor (YX/S) obtained with glucose were significantly different (p < 0.05) in comparison with pentoses, resulting in higher values. On the other hand, the biomass to product conversion factor (YP/X) presented higher values for the pentoses in relation to glucose, with arabinose differing from the other sugars (p < 0.05). These values indicate that glucose promotes cell growth (higher values of Xmax, µmax and YX/S). On the other hand, the pentoses stimulate a greater production of EPS by the cells (higher YP/X values).
FT-IR
Figure 2 shows the FT-IR spectra of EPS produced by Mesorhizobium sp. Semia 816, using the three different carbon sources (xylose, glucose and arabinose). As observed in the spectra, there is a similarity regarding the functional groups found in EPS, which indicates a possible structural similarity between them.

FTIR spectra of the EPS obtained from different carbon sources. (Dark green line) xylose; (light green line) glucose; (blue line) arabinose
The 3350 cm−1 band (1) is related to the presence of O–H group, which is a typical stretching of the polysaccharide vibration (Castellane et al. 2015). The presence of hydroxyl groups is directly related to the solubility in water (Castellane et al. 2015). The C–H stretching vibration of the CH2 group at 2900 cm−1 (2) was observed, usually present in hexoses and deoxyhexoses (Castellane et al. 2015). Furthermore, according to Annal and Daniel (2017), peaks between 3300 and 3500 cm−1 may represent a stretch of the N–H and C–N groups. The range between 2500 and 2000 cm−1 (3) is indicative of the presence of C–H (Annal and Daniel 2017). There is a broad peak at 1730 cm−1 (4) that represents the presence of C = O stretching of carbonyl groups of esters, suggesting pyruvate and acetate groups (Castellane et al. 2015; Andhare et al. 2017); and the 1714 cm−1 band (5) corresponds to carboxylic acids (Castellane et al. 2015). The band observed at 1300 cm−1 (6) may be related to carboxylate ions (Andhare et al. 2017) and at 1250 cm−1 (7) the peak can be attributed to acetyl groups (Priyanka et al. 2015), and bands between 1000 and 1150 cm−1 (8) indicate the presence of uronic acids (Deepika et al. 2016). The presence of these groups in these EPS is very important because the carboxylate group when linked to the glucuronate imparts anionic charge and the rhamnosyl side chains protect the carboxylic acids, preventing cross-linking by divalent calcium ions. Wingender et al. (1999) reported that the lack of intra-chain hydrogen bonds and location of anionic pyruvic ketal along with acetyl groups on the periphery of the helical structure of the polysaccharide are responsible for the absence of an ordered conformation. EPS from Mesorhizobium sp. Semia 816 showed peaks near 900 cm−1 (9), indicating the predominance of configuration β in the glycosidic bonds (Oliveira et al. 2020).
Monosaccharide composition
EPS produced by Mesorhizobium sp. Semia 816 is a polyanionic heteropolysaccharide, the repeating unit being formed by mannose, rhamnose, glucuronic acid, galacturonic acid, D-glucose and D-galactose. As shown in Table 2, the EPS have glucose as the predominant component, with lower amounts of mannose and galactose. The EPS obtained from xylose and glucose presented similar compositions. However, EPS produced with arabinose presented a different composition, with rhamnose content 1.8 times higher than that obtained from xylose cultivation. Furthermore, the glucuronic acid content of EPS from arabinose was 1.8 times higher when compared with xylose, while it had the lowest glucose content concerning the other EPS studied. Therefore, the carbon source used in bacterial culture may influence the composition of EPS.
Apparent viscosity
As shown in Fig. 3, pseudoplastic behavior was observed for the 1% w/v EPS aqueous solutions, with viscosity decreasing as the shear rate increases. Similar behavior was observed for several EPS produced by Rhizobium genus, such as R. radiobacter (Andhare et al. 2017) and R. tropici (Moretto et al. 2015; Castellane et al. 2017). This behavior is a key factor for food applications, favoring the flow during some unit operations and food texture modification (Andhare et al. 2017; Oliveira et al. 2020).

Variation of viscosity versus shear rate of EPS obtained from different carbon sources. (open circle) Xylose; (filled square) glucose; (open triangle) arabinose
In Table 3, the rheological parameters were determined using the Herschel–Bulkley mathematical model, which describes time-independent non-Newtonian fluids, fluids that need an initial shear stress to start the flow.
The rheological parameters indicated that the carbon source did not affect the rheological behavior. The parameters did not differ for the three carbohydrates, as well as the share rate evaluated at 10 and 100 s−1. The flow behavior index of less than 1 indicates pseudoplastic behavior, typical of bacterial biopolymers.
Thermal behavior
From the TGA curves (Fig. 4a), the degradation of the different samples can be observed as a function of temperature increase. According to the TGA curves, the degradation temperatures of the EPS samples were 290 °C (xylose and arabinose) and 275 °C (glucose), respectively. Furthermore, a mass loss of 74%, 70% and 64% was observed for the EPS obtained from xylose, glucose and arabinose, respectively.

TGA and DSC curves of EPS samples. (Dark green line) xylose; (light green line) glucose; (blue line) arabinose
Bakhtiyari et al. (2015), in their study of a succinoglycan polysaccharide, obtained a degradation temperature of 249.67 °C when sucrose was used as a carbon source and 261.04 °C when molasses was used. Other biopolymers with different initial degradation temperatures were cited, such as xanthan gum (266.4 °C), arabic gum (274.6 °C) and carboxymethylcellulose (280.2 °C) (Zohuriaan and Shokrolahi 2004).
In any case, TGA confirmed that all the EPS generally possessed high thermal stability, an important feature to be considered when choosing future applications. Furthermore, the EPS with xylose and arabinose as carbon source showed similar degradation behavior compared to EPS obtained with glucose. The difference in the mass loss value of the samples can be explained by the difference in molecular weight, structure and presence of different substituents (Bakhtiyari et al. 2015).
The DSC thermograms (Fig. 4b) show an endothermic peak at about 70 °C for the three EPS obtained with different carbon sources. This behavior was similar to that observed by Bakhtiyari et al. (2015) and can be attributed to the breakdown of hydrogen bonds and the changes in conformation of the double helices of the polysaccharide, giving rise to fusion of aggregates and network break up. In the production of curdlan by Agrobacterium sp. IFO 13,140, Mangolim et al. (2017) attributed the peak between 70 and 80 °C to the effect of hydrophobic interactions between the curdlan molecules.
EPS from hydrolysate
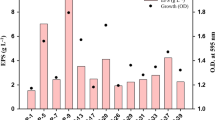
According to Fig. 5, Mesorhizobium sp. Semia 816 was able to grow and produce EPS using a soybean hulls hydrolysate-based medium. Glucose (3.7 g/L), xylose (1.3 g/L) and arabinose (1.5 g/L), among other carbohydrates (e.g., cellobiose), were detected in the hydrolysate. The amount of hydrolysate added to the medium was calculated in order to result 10 g/L reducing sugars. Maximum biomass concentration of 1.26 ± 0.02 g/L was reached in 24 h cultivation, while EPS reached a maximum value in 48 h (3.93 ± 0.06 g/L). Despite the fact that inhibitory effects are observed when lignocellulosic hydrolysates are used in microbial cultivations (Jönsson et al 2013), in this case biomass and EPS concentrations were slightly higher than those obtained with individual monosaccharides (Table 1). After hydrolysis of polysaccharides, lignin remains as a solid residue, although a minor part is degraded to phenolics and other aromatic compounds that act as inhibitors (Jönsson et al. 2013), but the soybean hulls hydrolysate used in this work presents a low level of phenolic compounds (unpublished data), reducing possible inhibitory effects. In addition, the presence of small quantities of lipids, proteins and minerals in the hydrolysate can contribute to a best performance of the microorganism.

Mesorhizobium sp. Semia 816 grown on a soybean hulls hydrolysate-based-medium. (open circle) Biomass; (filled square) pH; (open triangle) EPS
The aqueous-based suspension (1% w/v) prepared with EPS from hydrolysate presented flow behavior index lower than 1 (n = 0.44) and a significant resistance to flow, according to yield stress (σo = 32.7 ± 1.2 Pa) observed in Fig. 6, typical characteristics of Herschel–Bulkley type fluids. The shear rate (\(\dot{\gamma })\) applied in our rheological measurements, from 101 to 102 s−1, corresponds to the deformation verified in the processes of chewing and swallowing, respectively (Steffe 1996), i.e., allowing to predict the apparent viscosity perceived during ingestion of the biopolymer. Thus, this rheological behavior at low shear rate is important to impart good sensory quality, such as mouth feel, flavor release and suspension properties in food products.

Flow curve of an aqueous-based suspension (1% w/v) prepared with EPS obtained from soybean hulls hydrolysate
Conclusion
In this work, the effect of three different carbon sources was investigated, monosaccharides commonly found in lignocellulosic biomass, on EPS production by Mesorhizobium sp. Semia 816 and their characteristics. The study presented here has demonstrated that there were no significant differences (p > 0.05) in relation to EPS yield, reaching 3.39 g/L (glucose), 3.33 g/L (arabinose) and 3.27 g/L (xylose). Some structural differences were observed in relation to monosaccharide composition. However, they did not lead to significant differences in rheological behavior. In the case of soybean hulls hydrolysate used as carbon source, slightly higher values were obtained (3.93 g/L). This result indicates the potential use of lignocellulosic hydrolysates in the cultivation of this microorganism for the production of EPS of industrial interest, with possible impacts on the reduction of process costs.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Alfenore S, Molina-Jouve C (2016) Current status and future prospects of conversion of lignocellulosic resources to biofuels using yeasts and bacteria. Process Biochem 51:1747–1756. https://doi.org/10.1016/j.procbio.2016.07.028
Andhare P, Delattre C, Pierre G, Michaud P, Pathak H (2017) Characterization and rheological behaviour analysis of the succinoglycan produced by Rhizobium radiobacter strain CAS from curd sample. Food Hydrocolloid 64:1–8. https://doi.org/10.1016/j.foodhyd.2016.10.008
Annal DJM, Daniel RR (2017) Isolation and characterization of exopolysaccharides from a few selected Rhizobium species. Int J Biol Pharm Res 8:24–28. https://doi.org/10.21276/ijbpr.2017.8.1.5
Bakhtiyari M, Moosavi-Nasab M, Askari H (2015) Optimization of succinoglycan hydrocolloid production by Agrobacterium radiobacter grown in sugar beet molasses and investigation of its physicochemical characteristics. Food Hydrocolloid 45:18–29. https://doi.org/10.1016/j.foodhyd.2014.11.002
Castellane TCL, Lemos MVF, Lemos EGM (2014) Evaluation of the biotechnological potential of Rhizobium tropici strains for exopolysaccharide production. Carbohyd Polym 111:191–197. https://doi.org/10.1016/j.carbpol.2014.04.066
Castellane TLC, Otoboni AMMB, Lemos EGM (2015) Characterization of exopolysaccharides produced by Rhizobia species. R Bras Cienc Solo 39:1566–1575. https://doi.org/10.1590/01000683rbcs20150084
Castellane TCL, Campanharo JC, Colnago LA, Coutinho ID, Lopes EM, Lemos MVF, Lemos EGM (2017) Characterization of new exopolysaccharide production by Rhizobium tropici during growth on hydrocarbon substrate. Int J Biol Macromol 96:361–369. https://doi.org/10.1016/j.ijbiomac.2016.11.123
Costa OYA, Raaijmakers JM, Kuramae EE (2018) Microbial extracellular polymeric substances: ecological function and impact on soil aggregation. Front Microbiol 9:1636. https://doi.org/10.3389/fmicb.2018.01636
Deepika KV, Raghuram M, Kariali E, Bramhachari PV (2016) Biological responses of symbiotic Rhizobium radiobacter strain VBCK1062 to the arsenic contaminated rhizosphere soils of mung bean. Ecotox Environ Safe 134:1–10. https://doi.org/10.1016/j.ecoenv.2016.08.008
Duta FP, França FP, Lopes LMA (2006) Optimization of culture conditions for exopolysaccharides production in Rhizobium sp. using the response surface method. Electron J Biotechnol 9:391–399. https://doi.org/10.2225/vol9-issue4-fulltext-7
Ibbett R, Gaddipati S, Davies S, Hill S, Tucker G (2011) The mechanisms of hydrothermal deconstruction of lignocellulose: new insights from thermal–analytical and complementary studies. Bioresour Technol 102:9272–9278. https://doi.org/10.1016/j.biortech.2011.06.044
Jida M, Assefa F (2012) Phenotypic diversity and plant growth promoting characteristics of Mesorhizobium species isolated from chickpea (Cicer arietinum L.) growing areas of Ethiopia. Afr J Biotechnol 11:7483–7493. https://doi.org/10.5897/AJB11.4109
Jönsson LJ, Alriksson B, Nilvebrant N-O (2013) Bioconversion of lignocellulose: inhibitors and detoxification. Biotechnol Biofuels 6:16. https://doi.org/10.1186/1754-6834-6-16
Mangolim CS, Silva TT, Fenelon VC, Nascimento A, Sato F, Matioli G (2017) Use of FT-IR, FT-Raman and thermal analysis to evaluate the gel formation of curdlan produced by Agrobacterium sp. IFO 13140 and determination of its rheological properties with food applicability. Food Chem 232:369–378. https://doi.org/10.1016/j.foodchem.2017.04.031
Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31:426–428. https://doi.org/10.1021/ac60147a030
Moretto C, Castellane TCL, Lopes EM, Omori WP, Sacco LP, Lemos EGM (2015) Chemical and rheological properties of exopolysaccharides produced by four isolates of rhizobia. Int J Biol Macromol 81:291–298. https://doi.org/10.1016/j.ijbiomac.2015.07.056
Oliveira JM, Amaral SA, Burkert CAV (2018) Rheological, textural and emulsifying properties of an exopolysaccharide produced by Mesorhizobium loti grown on a crude glycerol-based medium. Int J Biol Macromol 120:2180–2187. https://doi.org/10.1016/j.ijbiomac.2018.06.158
Oliveira JM, Michelon M, Burkert CAV (2020) Biotechnological potential of soybean molasses for the production of extracellular polymers by diazotrophic bacteria. Biocatal Agric Biotechnol 25:101609. https://doi.org/10.1016/j.bcab.2020.101609
Priyanka P, Arun AB, Ashwini P, Rekha PD (2015) Versatile properties of an exopolysaccharide R-PS18 produced by Rhizobium sp. PRIM-18. Carbohyd Polym 126:215–221. https://doi.org/10.1016/j.carbpol.2015.03.017
Saranya Devi E, Vijayendra SVN, Shamala TR (2012) Exploration of rice bran, an agro-industry residue, for the production of intra- and extra-cellular polymers by Sinorhizobium meliloti MTCC 100. Biocatal Agric Biotechnol 1:80–84. https://doi.org/10.1016/j.bcab.2011.08.014
Senthil Kumar M, Swarnalakshmi K, Annapurna K (2017) Exopolysaccharide from Rhizobia: production and role in symbiosis. In: Hansen AP et al (eds) Rhizobium biology and biotechnology. Springer, Cham, pp 257–292
Steffe JF (1996) Rheological methods in food process engineering. Freeman Press, East Lansing
Tan IKP, Broughton WJ (1981) Rhizobia in tropical legumes XIII. Biochemical basis of acid and alkali reactions. Soil Biol Biochem 13:389–393. https://doi.org/10.1016/0038-0717(81)90083-3
Wingender J, Neu TR, Flemming H-C (1999) Microbial extracellular polymeric substances: characterization, structure, and function. Springer, Berlin
Zhou F, Wu Z, Chen C, Han J, Ai L, Guo B (2014) Exopolysaccharides produced by Rhizobium radiobacter S10 in whey and their rheological properties. Food Hydrocolloid 36:362–368. https://doi.org/10.1016/j.foodhyd.2013.08.016
Zohuriaan MJ, Shokrolahi F (2004) Thermal studies on natural and modified gums. Polym Test 23:575–579. https://doi.org/10.1016/j.polymertesting.2003.11.001
Acknowledgements
The authors thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001 and Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS).
Funding
This study was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior.
Author information
Authors and Affiliations
Contributions
CAVB, EGML and TCLC contributed to the conception, design and supervision of the study. BCSR and RGV carried out the cultivation assays. BCSR and TCLC carried out the characterization of EPS samples and data analysis. All authors contributed to the first draft of the manuscript and manuscript revision.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Research involving human participants or animals
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Roesler, B.C.S., Vaz, R.G., Castellane, T.C.L. et al. The potential of extracellular biopolymer production by Mesorhizobium sp. from monosaccharide constituents of lignocellulosic biomass. Biotechnol Lett 43, 1385–1394 (2021). https://doi.org/10.1007/s10529-021-03119-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-021-03119-9