
Abstract
We explored the temporal and spatial variations in airborne Alternaria spore quantitative and phenological features in Europe using 23 sites with annual time series between 3 and 15 years. The study covers seven countries and four of the main biogeographical regions in Europe. The observations were obtained with Hirst-type spore traps providing time series with daily records. Site locations extend from Spain in the south to Denmark in the north and from England in the West to Poland in the East. The study is therefore the largest assessment ever carried out for Europe concerning Alternaria. Aerobiological data were investigated for temporal and spatial patterns in their start and peak season dates and their spore indices. Moreover, the effects of climate were checked using meteorological data for the same period, using a crop growth model. We found that local climate, vegetation patterns and management of landscape are governing parameters for the overall spore concentration, while the annual variations caused by weather are of secondary importance but should not be neglected. The start of the Alternaria spore season varies by several months in Europe, but the peak of the season is more synchronised in central-northern Europe in the middle of the summer, while many southern sites have peak dates either earlier or later than northern Europe. The use of a crop growth model to explain the start and peak of season suggests that such methods could be useful to describe Alternaria seasonality in areas with no available observations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Alternaria is associated with more than 1100 common names (Toth et al. 2011) and consists of more than 300 species (Seifert et al. 2011). Spores from Alternaria are known to cause allergic problems worldwide (Burbach et al. 2009; Gioulekas et al. 2004; Mari et al. 2003). Fungal spores in general, and particularly those of Alternaria, induce respiratory allergies, most commonly among children and other sensitive individuals (Behbod et al. 2015; Knutsen et al. 2012). Fungal spores also cause acute respiratory failure, thus being frequently responsible for hospital admissions (Bush and Prochnau 2004; Dales et al. 2000). Nonetheless, fungi are reported as a neglected and underestimated source of respiratory allergy (Crameri et al. 2014).
A number of Alternaria species are also important plant pathogens (Gravesen et al. 1994; Oerke and Dehne 2004; Simmons 2007). Some species frequently infect important crops such as potatoes (Escuredo et al. 2011; Iglesias et al. 2007) and cereals (Skjøth et al. 2012). Alternaria mainly grows on vegetation and is considered to be related to agricultural landscapes (Sadyś et al. 2015; Skjøth et al. 2012). Many species of Alternaria are known to be host specific, such as Alternaria brassicae that affects specific Brassica species (Su’udi et al. 2013). The abundance of Alternaria in crops is related to the development and decomposition of biomass of their hosts (Escuredo et al. 2011; Iglesias et al. 2007). It has been calculated that Alternaria alternata can produce up to 3.2 × 105 spores per cm3 of nutrient medium, tested in two different media, rich one (PDA: potato-dextrose agar) and a poor one (PCA: potato-carrot agar) and at a variety of air temperatures (Damialis et al. 2015a). As nutrient availability significantly varies among different crop systems and vegetation types across different areas of Europe, it is expected that the total amount of Alternaria spores detected in the air in any given season (seasonal fungal spore index, SFI) will vary as well.
Spatial and temporal studies on Alternaria in Europe are rare. Most of the temporal studies focus on just one or two sites (Corden et al. 2003; Damialis et al. 2015b; Kasprzyk and Worek 2006). Continental-scale studies are non-existent to date. A recent study that focused on the spatial aspect in central and eastern Europe only covered one season and four countries (Kasprzyk et al. 2015). This scarcity of information on both temporal and spatial patterns on fungal spores has caused expression of concerns from both the biodiversity perspective and the health perspective (Berman 2011). There is therefore a general need for a Europe-wide assessment of fungal spore concentrations and how they vary in time and space.
The SFI has been shown to vary with more than a factor of four during a 10-year period (Corden et al. 2003; Damialis et al. 2015b; Skjøth et al. 2012). In Europe, local climate and vegetation are relatively stable over a 10-year period. Major land cover classes hardly change, and these classes include olive and grapes or areas with crops under rotation (European Commission 2005). The annual changes must therefore be attributed to changes in weather and management of the landscape. On the contrary, the amount of hosts varies spatially to a large degree. The abundance of crops, agricultural land and other vegetation patterns that are hosts of Alternaria can vary by a factor of 10 or more on both a 5 km × 5 km scale (Sadyś et al. 2014, 2015) or a 50 km × 50 km scale (Skjøth et al. 2010). Local management (e.g. harvesting and use of pesticides) of agricultural areas and vegetation pattern is therefore likely to have a larger impact on the SFI than the annual variations in weather.
The start of the Alternaria season in each area of Europe is likely related to the development of biomass of early crops. The development of biomass in crops can be simulated with simple crop growth models using accumulation of heat units (e.g. Deen et al. 2001; McMaster and Wilhelm 1997). The development of pathogens is also mainly determined by temperature (e.g. Agrios 1997). These two aspects—development of biomass and the effect of temperature on pathogens—make it likely that the start of the Alternaria season can be simulated with a simple crop growth model that responds to temperature, e.g. by accumulating the heat units above a specific threshold, the so-called growing degree days (GDD). Each area of Europe has its own specific crops (e.g. Balkovic et al. 2013). As a consequence, the simulation of crop growth will require different parameters between the areas. Generally, it is expected that temperature will comprise a crucial factor in the start of the Alternaria spore season. Nonetheless, temperature can be non-significant dependent on the species under study, on biological requirements and nutrient availability, among areas and sampling years etc. (e.g. Damialis et al. 2015a).
Peak concentrations of Alternaria are important as the exceedance of thresholds usually governs when sensitive patients react to spore exposure. The threshold for Alternaria, as reviewed by Gravesen (1979), has been estimated to be 100 spores per cubic metre of air (s m−3). This suggests that knowledge of local peak concentrations is important information and relevant to forecast. The annual peak concentrations of Alternaria have been shown to be related to specific agricultural operations like harvesting of mature crops (Friesen et al. 2001; Skjøth et al. 2012). The development of crop growth and maturation can also be simulated with simple crop growth models (McMaster and Wilhelm 1997). However, Alternaria peak concentrations can also be related to long-distance transport episodes (Burshtein et al. 2011; Sadyś et al. 2015). A consequence is that peak concentrations of Alternaria are not necessarily related to local crop development, but can also depend on atmospheric transport from areas with mature crops and harvesting (Fernández-Rodríguez et al. 2015; Skjøth et al. 2012). Alternaria concentrations can also decrease in the middle of the crop growth season as observed in the Mediterranean region (Escuredo et al. 2011; Maya-Manzano et al. 2012). Most likely the drop is caused by high temperatures which are known to cause a decrease in spore production in many species (e.g. Hauptman et al. 2013). This is particularly true in the case of Alternaria alternata, which decreased its spore production under experimentally elevated air temperature (Damialis et al. 2015a). Overall, since the peak concentration of any air particle is in fact a daily value and as such it is subject to high randomness (because of meteorological factors, anthropogenic influence and unpredictable parameters), Alternaria peak spore season and relationships with accumulated temperatures above a threshold (GDD) are considered only of secondary importance.
The aim of this study was to investigate potential relationships of the characteristics of the overall spore season of the genus Alternaria (start, end, duration, peak and SFI) with local climate throughout Europe. To achieve this, we tested whether the start of the Alternaria season in Europe was statistically related to GDD by applying a crop growth model. At the same time, we checked whether peak concentrations had weaker statistical relationship with GDD than the start of the season. As agricultural areas are considered to be the main source of Alternaria spores, we assessed the spatial abundance of Alternaria spores in four of the main agricultural areas in Europe by exploring the SFI in the following four biogeographical regions, the Atlantic, the Continental, the Mediterranean and the Pannonian, in a similar way to Grewling et al. (2012). Finally, we investigated whether the variation in SFI in all biogeographical regions was higher spatially rather than temporally. Our assumption was that variability in spatial and temporal scales is of similar magnitude and that any differences observed in airborne spore abundances and spore seasons would be mainly due to parallel weather and land use variations over large geographical areas. We hypothesised that both spatial variation (due to larger-scale phenomena, like crop and land use changes and climatic indices) and temporal variation (like short-scale incidents of local meteorology and yearly extreme weather events) would be of similar significance.
2 Materials and methods
2.1 Aerobiological data
Daily concentrations of Alternaria spores were used. The data were obtained using 7-day volumetric samplers (Hirst 1952). This sampler manages to monitor different air particles in the atmosphere and can therefore be used for the analysis of both pollen and spore airborne concentrations (instead of using two separate air samplers for each particle type). The aerobiological data were obtained from 23 sites in Europe (Fig. 1; Table 1). Coordinates and citations to a full description of the sites and their surroundings are given in Table 1. Observations are obtained according to aerobiological recommendations, typically on the top of a tall building so that the site represents the regional concentration, 30 km from the sampler. The counting method used during spore identification was either transverse traverses (Šikoparija et al. 2011) or longitudinal transects (Käpyla and Penttinen 1981). Aerobiological sampling standards, like the sample size per year and study area, were according to the minimum requirements for aerobiological sampling, as specified by Galán et al. (2014). All counts have been converted to daily mean concentrations of fungal spores per m3 of air (s m−3). Available data during the period 2000–2014 have been used to provide descriptive statistical evaluation in tabular form in a similar way to Kasprzyk and Worek (2006) including SFI, start and end of season, and peak concentrations. Days are here expressed as calendar days. Season start and season end are based on the 90 % method (Nilsson and Persson 1981) which is the most common method in aerobiological studies, including fungal spores (e.g. Gioulekas et al. 2004). Years with data gaps of 7 or more days during the Alternaria season were not included in the analysis.

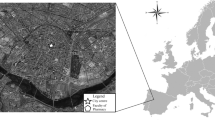
Location of meteorological stations and fungal spore sites used in the study. Names are placed to the top right, the top or the right according to location of the monitoring site, except for Vielha that is placed to the left. Names of the biogeographical regions refer to the official nomenclature given by the European Environmental Agency
2.2 Land cover and biogeographical data
The sites were grouped into the main biogeographical regions of Europe. We used the official biogeographical boundaries that are defined by European Environmental Agency and freely available as a shape file for use in GIS systems. The biogeographical regions are used in international regulations such as the Habitat Directive. Biogeographical regions can be transboundary and define areas where animal and plant distributions are having similar or shared characteristics, which means that these regions are well suited for broad continental-wide studies.
Each site was analysed with respect to nearby land cover from the Corine Land Cover data set (CLC2000). CLC2000 is the most detailed uniform international land cover data set currently available (e.g. Thibaudon et al. 2014) for this type of studies. Here, we analyse the vegetation cover within a 30-km radius from each site in a similar way to previous studies covering a large number of monitoring sites (Skjøth et al. 2010; Thibaudon et al. 2014; Karrer et al. 2015). This distance is found using the assumption that the overall pollen or spore index reflects the amount of plants within a distance of 30 km from a monitoring site (see Skjøth et al. 2010). Then, long-distance transport episodes are episodic (e.g. Sadyś et al. 2014; Smith et al. 2008) and therefore have the potential to contribute to the overall index (e.g. Fernandez-Rodrigues et al. 2015), but expectedly to a lower extent. The total area of each land cover type is calculated within the 30-km radius for each station using ArcGIS version 10 and is available as supplementary information, and together with the percentage of agricultural areas (CLC2000 codes 211–244, see supplementary material) is listed in Table 1.
2.3 Air temperature data
Meteorological parameters were obtained from nearby stations by analysing the Global Summary of the Day (GSOD) meteorological data set that is exchanged as part of the World Weather Watch Programme within the World Meteorological Organization (WMO). Each aerobiological site was then matched with the nearest meteorological site (Table 1) in order to calculate local GDD. Here we use the WMO definition of GDD, which is the daily mean temperature above a critical threshold. We use a critical threshold value of 0 °C as given in the agricultural model MV-Tool (Olesen and Plauborg 1995) and a starting date of 01 March as this combination is based on empirical calibration for common crops like spring barley and winter wheat throughout the majority of the study area (Gyldenkærne et al. 2005). We therefore use GDD as a proxy for overall crop development within a particular area. It has previously been shown that the annual peaks of Alternaria concentrations are at least partly related to harvesting of mature crops and augmentation of the spore concentration from remote sources (Skjøth et al. 2012). The long-term mean of both seasonal characteristics and GDD are then used for climatological assessments on a spatial scale throughout Europe. Data gaps in the meteorological time series shorter than a week were filled by interpolation or by using data from a nearby station if there were a very high autocorrelation and no significant bias between sites.
2.4 Seasonal fungal index
The SFI was calculated for each year and for each site during the period 2000–2014 (Table 2), and the mean of the SFI is displayed on a map (Fig. 2). Sites with five or more years of Alternaria data were then analysed for the maximum temporal variation in the data from that particular site. This was achieved by calculating the maximum and minimum daily concentrations per year and per site and expressing them as percentages in relation to the annual mean for the respective year and site. For each site, the temporal variation is the difference between these maximum and minimum values. The total temporal variation for all relevant sites is given in Table 2 as an estimate of local changes due to weather and management using following formula: 100 × (max/average − min/average). As such, the calculation of the temporal variation is almost identical to the calculation of the coefficient of variation (given as percentage in relation to the mean value), where the focus is on the standard deviation instead of maximum and minimum values. Spatial variation has also been grouped by the main biogeographical regions in Europe as defined by the European Environment Agency (EEA 2012). The total spatial variation for all relevant years is given in Table 2. This is assumed to be an estimate of variations in SFI due to vegetation type and vegetation pattern (thus the spatial distribution), its management and large-scale climatic conditions within the area. The extent of the total temporal variation was then compared with the extent of the total spatial variation in order to test whether vegetation (and its long-term management such as choice of land cover), vegetation pattern and overall climate have a larger impact on the SFI than annual changes in weather and short-term management of the landscape (e.g. harvesting, choice of annual crops and use of pesticides). Finally, the total spatial variation in SFI was linearly correlated with respect to the fraction of agricultural land, for the entire study area and each biogeographical region with more than 5 sites.

Location of spore sites, their mean seasonal fungal spore index and the mean length of the spore season. Numbers are placed to the top right, the top or the right according to location of the monitoring site
2.5 Start and peak dates of fungal season
The relationships between cumulative temperature above the threshold (GDD) and starting days as well as GDD and peak days were first analysed for each site with hierarchical clustering on data sets with five or more years available (Table 3). The calculations were done with the pvclust, which is an add-on package for R statistical software to assess the uncertainty in hierarchical cluster analysis (Suzuki and Shimodaira 2006; Suzuki 2014). Pvclust calculates probability values (p values) for each cluster using bootstrap resampling techniques. Two types of p values are calculated: approximately unbiased (AU) p value and bootstrap probability (BP) value. Clusters that are highly supported by the data will have large p values. Average clustering agglomerative method and the default number of bootstrap replications that were equal to 1000 were used in this study to obtain clusters. For each site, maximally two clusters were further considered for the analysis. If the clustering revealed outliers, they were not taken into account for the creation of mean values (centres of the clusters) of starting day and peak day (Fig. 3). Centres of the clusters were calculated with the k-means function available in the R package stats library (R Core Team 2015). The method to get cluster centres aims to the minimization of sum of squares from points to the assigned cluster centres. The k-means function has an nstart option that attempts multiple initial configurations and reports on the best one. The recommended number for nstart of 25 was used here.

Fungal spore sites and the associated start of season and peak day. Numbers are placed to the top right, the top or the right according to location of the monitoring site
3 Results
Spore data sets were available from 23 sites providing annual summaries from daily data that passed the quality criterion (no gaps of considerable size). These data varied in length from 3 years (Badajoz and Thessaloniki) to 15 years (Barcelona, Bordeaux, Girona, Kraków and Tarragona). Corresponding meteorological data were available for each year for 22 out of the 23 sites (Table 1). The spatial pattern in the start of the Alternaria season shows a gradient from the south to the north of about 3 months (Fig. 3) with the earliest start in Spain and Greece in April–May and the latest in Denmark in mid-June to mid-July. Similarly, the length of the season is up to 4 months longer in the south than in the north generally ranging from 2–3 months in Denmark to 6–7 months in Greece and even more in Spain. The annual peak dates of Alternaria concentrations show hardly any gradient from south to north (Fig. 3; Table 2) with respect to the mean value. However, Fig. 5 shows that in the continental and Atlantic regions, the peak dates are usually between calendar days 180 and 240, with mean values as in Fig. 3. In contrast, the Pannonian and the Mediterranean regions have hardly any peak values in the middle or late summer with very few values between 220 and 250. The peak values are instead clustered before day 220 (early summer) and after day 250 (autumn), thus suggesting a bimodal pattern. In the Continental and Atlantic regions, the peak dates are found in July–August (Fig. 2; Table 3), while there is much larger variation between years in the Mediterranean region and few peak values in August.
The highest SFIs were exhibited scattered across Europe, like in Hungary and the UK and a single site in NE Spain; little direct connection between SFI and season length was observed (Fig. 2). Therefore, in England the longest season is in Worcester, but the highest SFIs are in Leicester and Derby. Likewise, in France one of the largest SFIs occurred in Paris, where, nonetheless, the spore season had a marked short duration. Within the four main biogeographical regions, the highest SFIs (Fig. 2; Table 2) are found in the Pannonian region (up to 59,587 conidia), Spain (up to 59,875 conidia) and England (up to 30,281 conidia), while the lowest SFIs are found in Mediterranean region (6005 conidia in Greece) and the northern part of the Continental region (2282 conidia in Denmark). It should be noted that the remote site Vielha in the alpine region of the Pyrenees is even lower with an SFI as low as 815 conidia. Overall, the spatial variation in SFI per year (calculated as the mean of yearly SFIs per site) is more than 1–2 factors larger than the temporal variation in SFIs (calculated as the mean SFI of all sites per year) (p < 0.05). In fact 14 out of 15 years have a larger spatial variation than any recorded temporal variation (Table 2). The fraction of agricultural land varied from 5.7 % at Vielha to 89.5 % at Lleida, and 12 out of 23 stations have this fraction above 70 % (Table 1). The majority originates from non-irrigated arable land, with various contributions from other categories such as olive groves, fruit trees and berry plantations and vine yards (supplementary information). The R 2 values with the overall seasonal fungal spore index are 0.10, 0.68, 0.00 and 0.53 for the entire area, the Mediterranean region, the Atlantic Region and the Continental region, respectively.
A statistically significant relationship was observed between both the start and the peak of the spore season and GDD. This was true for all sites, except for Krasne in Poland (Table 3). In the Atlantic and Continental regions, the GDD for peak dates have a tendency to more clustering than the start of the season (Figs. 4, 5). The Atlantic and the Continental regions also have a higher tendency to clustering than the Pannonian and Mediterranean regions (Figs. 4, 5). Most sites have the GDD values in two main clusters (Table 3). For the start of the season, the clusters correspond to calendar dates in April–June and May–July. The GDD values range from 968 to 1274 °C for 11 out of 20 GDD values. The remaining values range from 9.2 to 1489 °C. The annual season peaks have clusters that are mainly in July–August. For the peak, 11 out of 20 cluster values lie in the range 1880 to 2102 °C, while the rest of the GDD values are scattered from 1393 to 5277 °C.

Annual start of season versus annual growing degree days (GDD) at individual sites and grouped into the four major biogeographical regions: a Atlantic, b Continental, c Pannonian and d Mediterranean. Data for each site are clustered in either one or two groups. Clustering values and the corresponding p value are found in Table 2

Annual peak of season versus annual growing degree days (GDD) at individual sites and grouped into the four major biogeographical regions: a Atlantic, b Continental, c Pannonian and d Mediterranean. Clustering values and the corresponding p value are found in Table 2
4 Discussion
Our results support the hypothesis that the SFI of Alternaria is more affected by local land use management, vegetation structure and overall climate than year-to-year variations in weather. If this hypothesis is correct, then it would mean that airborne concentrations of Alternaria spores exhibit higher variation spatially rather than temporally. The results also support the close relationship of Alternaria season with climate expressed as GDD, given that the results show large variations in GDD values between the biogeographical regions. However, the annual peak concentration of Alternaria was found to have a weaker statistical relationship with GDD than the start of the spore season when treating Europe as a whole. Contrary to this, the peak dates had a stronger relationship with GDD than the starting dates in both central and northern Europe and weaker relationship in southern Europe. Different climatic conditions seem to play an important role in this phenomenon. The Mediterranean, with mild winters and hot summers and with a more prolonged spore season in all cases (Mediterranean sites vs. all other sites), exhibits an unpredictable occurrence of the daily peak concentration of Alternaria conidia compared to the short respective period of north and central Europe. If one also takes into account the increased biodiversity of the associated crops, grasslands and other sources of Alternaria conidia to the south, this phenomenon is even more probable. We found no linear correlation between agricultural land cover over the entire study area and SFI. However, the strong correlations between agricultural land and SFI in both the Mediterranean and Continental region suggest that in large parts of Europe, the land cover is a very important parameter for the prediction of the overall SFI. It should here be mentioned that the observed high R 2 values are based on a small data set. In any case, this suggest that spatially robust predictions of SFI should take into account both climate and vegetation cover.
For all sites, we found a statistical clustering of GDD values in relation to both the start and the peak of the season. In the northern part of the area studied, this clustering was stronger than the southern part for the peak of the season. This may be related to several things. Firstly, the GDD is directly related to temperatures. The higher the temperatures, the higher the GDD, with inevitably growing values by definition. As Alternaria spore season presents a more or less seasonal pattern, spore abundances will start decreasing after a specific time point. After this point (up to which parallel increases in GDD and spore concentrations take place), GDD inevitably continues to rise, whereas spore abundances start decreasing. This may be due to the threshold values required for each Alternaria species to sporulate, in each area, habitat, site, year and day. Overall, higher temperatures of south Europe lead to faster achievement of Alternaria sporulation thresholds, for the same species, which potentially leads to weaker clustering in south Europe. This matches well with the findings that June is the month with the highest number of peak concentrations (Fig. 5). This is particularly true in the southern parts of the study area, while in colder north areas, the relatively short season and low maximum temperatures do not exceed the required abovementioned threshold value. Secondly, it could be related to the harvesting of cereals, as two major crops in northern Europe are wheat and barley. According to our hypothesis, harvesting was expected to affect spore concentrations through long-distance transport (LDT). However, harvesting of cereals generally happens after several days of good sunny and dry weather (Skjøth et al. 2012), which is a synoptic scale meteorological phenomenon that simultaneously can cover large areas in central-northern Europe. When this happens, then LDT from one geographical area will have a smaller tendency to affect another area and increasing the otherwise low concentration through LDT. Instead, the LDT episodes will augment the already high concentrations within central-northern Europe. It is therefore unclear whether the measured Alternaria spores on high days are mainly due to either local sources or more remote sources and how atmospheric transport can affect the annual spore index in each area. This hypothesis is supported by Fig. 3 and Table 3 showing that the peak days are generally much more synchronised in central-northern Europe than the start dates, where there is a clear south–north gradient. Despite significant variations in actual starting dates, GDD values for each site (Table 3; Figs. 4, 5) are clustered around one or two values, in central and northern Europe, in particular. Several GDD values at one site can be related to the crop rotation system in Europe. The annual changes in the crop rotation areas will cause annual changes in the maturation of the crops. This must be expected to cause changes in Alternaria spore production and, as in this study, also in the values of required GDD values. As a consequence, there will be several GDD values for each site. These values are then interlinked with a strong south–north gradient in starting date with a difference of more than 2 months (Figs. 3, 4). It should be noted that both Figs. 4 and 5 could indicate a strong correlation between GDD and starting date of the fungal spore season (e.g. Figure 5c will then have an R 2 value of 0.97). This is partly true, because there is a very high autocorrelation between GDD and calendar date throughout most of Europe. Regardless of the length of time series, the SFI is relatively stable for a large number of sites and varies with less than 100 % from the mean values, mainly seen in the shorter time series (Table 2). The series with data covering 13–15 years show a temporal variation from 100 to 150 %, which suggests that 5 years are not sufficient to cover annual variability at a specific site. This is contrasted with the spatial variation as this is always above 100 %, even exceeding 200 % in 14 out of 15 cases. All these findings show that the seasonality of Alternaria is strongly linked with both overall climate for the area and to a smaller degree, but nevertheless important, with the weather of a particular season.
This study also finds large north–south variations in the length of the season as well as in the annual peak concentration. In the north, the annual peak concentrations always appear in July–August, while they can be either in May–June or September–October at the Mediterranean sites (Maya-Manzano et al. 2012). The reason for this variation is most likely related to the sensitivity of spore production to temperature as found for Alternaria alternata by Hatzipapas et al. (2002), suggesting that 25 °C was the optimal range for spore production. Certain experimental designs on Alternaria alternata have shown that spore production can decrease dramatically even within a relatively short temperature range (temperature difference of the magnitude of 0.9–1.6 °C) and especially under increased nutrient availability (Damialis et al. 2015a). Such high temperatures are frequently observed in the Mediterranean region, mainly during July–August. Hence, peak concentrations do not always occur in July–August (Fig. 5d). In the northern sites, with the short and more well-defined summer periods, peak concentrations are in the middle of the summer (Fig. 5b). The largest mean seasonal spore index is in Budapest, in the Pannonian region, but large indices are also found in Lleida, Spain, in the Mediterranean region, and in Leicester and Derby, in the Atlantic region. In the area of Catalonia within the Mediterranean region, the large index might be related to the fact that Lleida is a rural site (De Linares et al. 2010) and therefore has abundant sources nearby, while most of the other sites in this study are urban locations. The spore index of nearby site in Worcester, in the Atlantic region, is among the lowest recorded. Despite this, the season in Worcester is slightly longer or similar to Derby and Leicester. Both climatic differences as well as annual changes in weather between Worcester and Leicester can be considered negligible; they are both located in the Midlands so the main difference between them must be related to the abundance of local sources. One explanation could be related to the difference in crops. Worcester is known to contain large areas of permanent crops, such as orchards, while the area around Leicester, according to a map by Sadyś et al. (2015), contains large areas with crops under rotation. The importance of crops and the type of crops in each biogeographical region are underlined by the clustering of the GDD values. Areas with crops under rotation must by definition contain several types of crops, such as winter wheat or spring barley, that mature and are ready for harvesting at different times of the year (Cooter et al. 2012; Gyldenkærne et al. 2005). Consequently, different biogeographical regions often have specific phenotypes of crops, such as slow- or fast-growing species (e.g. Poorter et al. 1995), where these phenotypes are adapted to the local environment. Fungal mycelia, and consequent spore production, are related to harvesting and decomposition of vegetation, growing on living or dead tissue, and hence, the different crop types expectedly cause different peak dates in spore production due to maturation of the crops (and thus the associated GDD value). Similar or even larger differences in the SFI were found in the Spanish part of the Mediterranean region. It is also necessary to consider the use of fungicides on crops to interpret differences between places, as they may affect Alternaria abundance in the air. Overall, these findings suggest that despite the high randomness that one would expect when examining such diverse data sets across Europe and for a variety of geoclimatic and vegetation zones, over time, there is a key factor masking out all other factors examined, temperature, but also all others that have not been probably incorporated in this study. Surely the climatic element plays a role, but primarily a locally important parameter affects spore production within an area. This adds further support to the hypothesis that the SFI of Alternaria is more affected by local landscape management, vegetation structure and overall climate than year-to-year variations in weather. As many of the Alternaria species comprise endophytic organisms, they acquire the necessary nutrients by the plant hosts in which fungi grow. Damialis et al. (2015a) have also experimentally proved that Alternaria alternata’s spore production and mycelium growth are strongly correlated with the amount of available nutrients provided by their plant hosts. In fact, they are suggesting that various fungal species (among which Alternaria spp.) prefer, under climate change conditions to change their life strategy by, that is, elongating their growth phase and delaying their sporulation stage, until they exhaust the available nutrients. Gundel et al. (2013) have also documented that such changes in life strategies exist and that these influence the interactions between plants and fungi in an extremely complex and species-specific manner.
Globally, primary biogenic aerosols (PBA), such as pollen, bacteria and fungal spores, have a number of very important functions. Firstly, PBA are efficient as ice nuclei and their emission and concentrations are directly affected by meteorological factors (Hoose and Möhler 2012). According to the Intergovernmental Panel on Climate Change (IPCC), there has been limited research on these effects, so the associated feedback mechanism between PBA and climate is poorly understood (Kirtman et al. 2013). Secondly, a number of PBA, in particular fungal spores, are also important in relation to soil and vegetation in agriculture (e.g. Agrios 1997), to forestry (e.g. Sadyś et al. 2014) and to human health (e.g. Denning et al. 2014), but there are major knowledge gaps that have recently been identified by IPCC (Kirtman et al. 2013). Thirdly, according to an editorial in the Lancet (2008), the impact of PBA on human health in relation to allergies does not receive the scientific attention it needs. In response to the need for collecting and assessing large aerobiological data sets (Beggs 2004; Cecchi et al. 2010) to fill the identified knowledge gaps, a number of large-scale pollen studies have been recently published (e.g. Smith et al. 2014; Zhang et al. 2015), but studies on fungal spores remain sparse. This large-scale assessment is our attempt towards improving the existing knowledge on Alternaria, a major fungal genus that is important both as a phytopathogen and a potential aeroallergen.
Up to now, very little research in Europe has been conducted on fungal spore year-to-year variations, within-year variations and the magnitude of these changes to the SFI (e.g. Damialis et al. 2015b; Fernández-Rodríguez et al. 2015). Either very few years have been examined or few fungal taxa, whereas the spatial feature has never been addressed at the European scale. One study in Europe (Canaries Islands, Catalonia and Andalusia in Spain) has previously investigated Alternaria fungal spore concentrations in more than three sites (De Linares et al. 2010) and more than one season. The authors found large variations in Alternaria dynamics in the two biogeographical regions within Spain extending a previous regional study by Maya-Manzano (2012) in the same country. In our study, even larger variations were found in the main biogeographical regions of Europe. Damialis et al. (2015b) searched for long-term trends in a time series for a wide range of fungal taxa, 15 in total including Alternaria, during 1987–2005 from one site, Thessaloniki, Greece; they observed both a later onset and peak of the main spore season of Alternaria and a shorter overall season length. This corresponds well with our study, as peak concentrations are frequently absent during the middle of the summer in the Mediterranean region, where 17 out of the 85 annual peaks in that biogeographical region are in July/August and 54 are found in June, September and October (Fig. 5).
Kasprzyk et al. (2015) worked on an Alternaria-specific aerobiological data set including data from a single year but of a wide spatial distribution, from 12 monitoring sites of four countries across central and eastern Europe. They observed that there is a north–south shift mainly of daily peak spore concentrations depending on the latitude. The findings in our study considerably extend the study by Kasprzyk et al. (2015) by covering a larger geographical area and also by taking the temporal aspect into account. Kasprzyk et al. (2015) also find highest seasonal spore index in the most intense agricultural area and more than a factor of ten lower seasonal spore index in the Boreal biogeographical region where large areas are dominated by forests. These results, combined with the results from both the agricultural intense areas and the low seasonal spore index from the site at Vielha could suggest that the concentrations of Alternaria in large parts of Europe are due to both non-agricultural and agricultural sources, where the agricultural sources are dominating.
Our research exemplifies the strong effects of environmental factors on both the temporal and spatial characteristics of Alternaria fungal spore season. It combines both long-term data sets (frequently of more than a decade), for the most recent records (after 2000), for the most allergenic fungal genus, Alternaria (e.g. Gioulekas et al. 2004), and, above all, in the largest spatial scale to date, across Europe. The results obtained from our study are in agreement with relevant ones from studies of far smaller geographical or temporal scale (Damialis et al. 2015b; De Linares et al. 2010; Kasprzyk et al. 2015). In addition, they elucidate on the observed patterns and on the driving factors. Indeed, increased temperatures with decreased latitude (i.e. Mediterranean) lead to a later onset of spore season but also to a decreased overall abundance of spores, in terms either of annual concentration or daily peak concentration, at many sites. Despite this general pattern, large variations in the overall seasonal spore index are detected within a few hundred kilometres and within areas with similar climate like in north Spain (Catalonia) or the entire UK (Fig. 1). It seems that a high regional variation is associated with specific micrometeorological conditions and features of the local vegetation. Yearly variations as in Damialis et al. (2015b) are evident at most sites in Europe (Table 2). However, these are of secondary importance when compared to the effects of temperature, either directly, in terms of required GDDs, or indirectly, as simulated by latitude.
Because of the lack of previous knowledge, concerns have been expressed regarding this variability in fungal spores with respect to allergic sensitisation and biodiversity perspectives (Berman 2011). As sporulation of fungi is influenced by several meteorological factors, such as rainfall, air temperature and relative humidity (e.g. Cecchi et al. 2010; Sadyś et al. 2014, 2015), the ongoing climate change is expected to greatly influence the temporal and quantitative characteristics of fungal seasons worldwide. Therefore, specific studies are needed, as already suggested by Helfer (2014), to further investigate the interaction effects of fungal spore abundances with climate change temporally and spatially. This study suggests that the integrated effect of land management should also be included (e.g. land cover type such as woodland or agriculture, type of crops, use of pesticides). These studies should be conducted for a wide range of fungal taxa, chosen on the basis of their allergenic properties (e.g. Alternaria, Cladosporium, Aspergillus and Penicillium) or their phytopathogenic potential (e.g. Botrytis, Fusarium and Ganoderma). Whenever possible, these studies should include the use of atmospheric transport models such as HYSPLIT (Draxler et al. 2014) and actual quantification of the foot print area (Skjøth et al. 2015; Makra et al. 2010). Ideally, these studies should take into account existing climate as well as the expected effects of climate change. This will be a direct response to the suggestions by the Lancet (The Lancet 2008) and Cecchi et al. (2010) by focusing on the fungal spores as well as Berman (2011) by focusing on biodiversity. This study is the first step towards this objective.
5 Conclusion
We conclude that temporal and spatial variations in SFI and spore season start and peak derive from different sources: local climate and vegetation (including management agricultural landscape) are the governing parameters for the yearly spore concentration, while the annual variations are most likely caused by weather fluctuations. We find that the start of the spore season varies by several months across Europe, but the peak of the season is rather synchronised in central-northern Europe, in the middle of the summer, while southern sites in general have peak dates either earlier or later than northern Europe. We did not find a direct correlation between spore index on European scale and amount of agricultural land. However, the data suggest that within a region with similar climate and vegetation, as defined by a biogeographical region, a strong correlation between abundance of agricultural land and spore index may be present. Understanding the temporal and spatial dynamics of Alternaria requires knowledge of how spore production and release are affected by weather (short-term effects) and climate (decadal effects) and how the main habitats and the agricultural areas are affected by management. The results with the simple crop growth model to explain start and peak of season were encouraging due to the substantial clustering. This suggests that such methods could be useful to describe Alternaria seasonality in areas that are not covered by observational sites.
References
Agrios, G. N. (1997). Plant pathology. San Diego: Academic Press.
Balkovic, J., van der Velde, M., Schmid, E., Skalsky, R., Khabarov, N., Obersteiner, M., et al. (2013). Pan-European crop modelling with EPIC: Implementation, up-scaling and regional crop yield validation. Agricultural Systems, 120, 61–75.
Beggs, P. J. (2004). Impacts of climate change on aeroallergens: Past and future. Clinical and Experimental Allergy, 34, 1507–1513.
Behbod, B., Sordillo, J. E., Hoffman, E. B., Datta, S., Webb, T. E., Kwan, D. L., et al. (2015). Asthma and allergy development: Contrasting influences of yeasts and other fungal exposures. Clinical and Experimental Allergy, 45, 154–163.
Berman, D. (2011). Climate change and aeroallergens in South Africa. Current Allergy and Clinical Immunology, 24, 65–71.
Burbach, G. J., Heinzerling, L. M., Edenharter, G., Bachert, C., Bindslev-Jensen, C., Bonini, S., et al. (2009). GA(2)LEN skin test study II: clinical relevance of inhalant allergen sensitizations in Europe. Allergy, 64, 1507–1515.
Burshtein, N., Lang-Yona, N., & Rudich, Y. (2011). Ergosterol, arabitol and mannitol as tracers for biogenic aerosols in the eastern Mediterranean. Atmospheric Chemistry and Physics, 11, 829–839.
Bush, R. K., & Prochnau, J. J. (2004). Alternaria-induced asthma. Journal of Allergy and Clinical Immunology, 113, 227–234.
Cecchi, L., D’amato, G., Ayres, J. G., Galan, C., Forastiere, F., Forsberg, B., et al. (2010). Projections of the effects of climate change on allergic asthma: the contribution of aerobiology. Allergy, 65, 1073–1081.
Cooter, E. J., Bash, J. O., Benson, V., & Ran, L. (2012). Linking agricultural crop management and air quality models for regional to national-scale nitrogen assessments. Biogeosciences, 9, 4023–4035.
Corden, J. M., Millington, W. M., & Mullins, J. (2003). Long-term trends and regional variation in the aeroallergen Alternaria in Cardiff and Derby UK—Are differences in climate and cereal production having an effect? Aerobiologia, 19, 191–199.
Crameri, R., Garbani, M., Rhyner, C., & Huitema, C. (2014). Fungi: The neglected allergenic sources. Allergy, 69, 176–185.
Dales, R. O. B. E., Cakmak, S. A. B. I., Burnett, R. I. C. H., Judek, S. T. A. N., Coates, F. R. A. N., & Brook, J. E. F. F. (2000). Influence of ambient fungal spores on emergency visits for asthma to a regional children’s hospital. American Journal of Respiratory and Critical Care Medicine, 162, 2087–2090.
Damialis, A., Mohammad, A., Halley, J., & Gange, A. (2015a). Fungi in a changing world: Growth rates will be elevated, but spore production may decrease in future climates. International Journal of Biometeorology, 59, 1157–1167.
Damialis, A., Vokou, D., Gioulekas, D., & Halley, J. M. (2015b). Long-term trends in airborne fungal-spore concentrations: A comparison with pollen. Fungal Ecology, 13, 150–156.
De Linares, C., Belmonte, J., Canela, M., de la Guardia, C. D., Alba-Sanchez, F., Sabariego, S. A., & Nso-Perez, S. (2010). Dispersal patterns of Alternaria conidia in Spain. Agricultural and Forest Meteorology, 150, 1491–1500.
Deen, W., Swanton, C. J., & Hunt, L. A. (2001). A mechanistic growth and development model of common ragweed. Weed Science, 49, 723–731.
Denning, D. W., Pashley, C. H., Hartl, D., Wardlaw, A., Godet, C., Giacco, S. D., et al. (2014). Fungal allergy in asthma—state of the art and research needs. Clinical Biochemistry, 4, 1–23.
Draxler, R., Stunder, B., Rolph, G., & Stein, A., & Taylor, A. (2014). Hysplit4 users guide. Revision September 2014. http://www.arl.noaa.gov/documents/reports/hysplit_user_guide.pdf.
Dupuy, N. (2007). Lecture de spores fongiques. Technical Report, Reseau National de Surveillance Aerobiolique, Lyon.
Escuredo, O., Seijo, M., Fernández-González, M., & Iglesias, I. (2011). Effects of meteorological factors on the levels of Alternaria spores on a potato crop. International Journal of Biometeorology, 55, 243–252.
European Commission. (2005). Image2000 and CLC2000 products and methods. European Commission, Joint Research Center (DG JRC), Institute for Environment and Sustainability, Land Management Unit, I-21020 Ispra, VA.
Fernández-Rodríguez, S., Sadyś, M., Smith, M., Tormo-Molina, R., Skjøth, C. A., Maya-Manzano, J. M., et al. (2015). Potential sources of airborne Alternaria spp. spores in South-west Spain. Science of the Total Environment, 533, 165–176.
Friesen, T. L., De Wolf, E. D., & Francl, L. J. (2001). Source strength of wheat pathogens during combine harvest. Aerobiologia, 17, 293–299.
Galán, C., Smith, M., Thibaudon, M., Frenguelli, G., Oteros, J., Gehrig, R., et al. (2014). Pollen monitoring: Minimum requirements and reproducibility of analysis. Aerobiologia, 30, 385–395.
Gioulekas, D., Damialis, A., Papakosta, D., Spieksma, F., Giouleka, P., & Patakas, D. (2004). Allergenic fungi spore records (15 years) and sensitization in patients with respiratory allergy in Thessaloniki-Greece. J Invest Allergo Clin Imm, 14, 225–231.
Gravesen, S. (1979). Fungi as a cause of allergic disease. Allergy, 34, 135–154.
Gravesen, S., Frisvad, J. C., & Samson, R. A. (1994). Microfungi: Munksgaard. Copenhagen: Denmark. ISBN 9788716114365.
Grewling, Ł., Šikoparija, B., Skjøth, C. A., Radišić, P., Apatini, D., Magyar, D., et al. (2012). Variation in Artemisia pollen seasons in Central and Eastern Europe. Agricultural and Forest Meteorology, 160, 48–59.
Grinn-Gofron, A., & Strzelczak, A. (2008). Artificial neural network models of relationships between Alternaria spores and meteorological factors in Szczecin (Poland). International Journal of Biometeorology, 52, 859–868.
Gundel, P. E., Garibaldi, L. A., Helander, M., & Saikkonen, K. (2013). Symbiotic interactions as drivers of trade-offs in plants: Effects of fungal endophytes on tall fescue. Fungal Diversity, 60, 5–14.
Gyldenkærne, S., Ambelas Skjøth, C., Hertel, O., & Ellermann, T. (2005). A dynamical ammonia emission parameterization for use in air pollution models. Journal Geophysical Research, 110, 1–14. doi:10.1029/2004JD005459.
Hatzipapas, P., Kaloskak, K., Dara, A., & Christias, C. (2002). Spore germination and appressorium formation in the entomopathogenic Alternaria alternata. Mycological Research, 106(11), 1349–1359.
Hauptman, T., Pitcairn, C. E. R., de Groot, M., Ogris, N., Ferlan, M., & Jurc, D. (2013). Temperature effect on Chalara fraxinea: Heat treatment of saplings as a possible disease control method. Forest Pathology, 43, 360–370.
Helfer, S. (2014). Rust fungi and global change. New Phytologist, 201, 770–780.
Hirst, J. M. (1952). An automatic volumetric spore trap. Annals of Applied Biology, 39, 257–265.
Hoose, C., & Möhler, O. (2012). Heterogeneous ice nucleation on atmospheric aerosols: A review of results from laboratory experiments. Atmospheric Chemistry and Physics Discussions, 12, 12531–12621.
Iglesias, I., Rodríguez-Rajo, F., & Méndez, J. (2007). Evaluation of the different Alternaria prediction models on a potato crop in A Limia (NW of Spain). Aerobiologia, 23, 27–34.
Käpyla, M., & Penttinen, A. (1981). An evaluation of the microscopial counting methods of the tape in Hirst–Burkard pollen and spore trap. Grana, 20, 131–141.
Karrer, G., Skjøth, C. A., Šikoparija, B., Smith, M., Berger, U., & Essl, F. (2015). Ragweed (Ambrosia) pollen source inventory for Austria. Science of the Total Environment, 523, 120–128.
Kasprzyk, I., Rodinkova, V., Sauliene, I., Ritenberga, O., Grinn-Gofron, A., Nowak, M., et al. (2015). Air pollution by allergenic spores of the genus Alternaria in the air of central and eastern Europe. Environmental Science and Pollution Research, 22, 9260–9274.
Kasprzyk, I., & Worek, M. (2006). Airborne fungal spores in urban and rural environments in Poland. Aerobiologia, 22, 169–176.
Kirtman, B., Power, S. B., Adedovin, J. A., Boer, G. J., Bojarju, R., Camiloni, I., et al. (2013). Near-term climate change: Projections and predictability. In T. F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S. K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex, & P. M. Midgley (Eds.), Climate Change 2013: The physical science basis. Contribution of Working Group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
Knutsen, A. P., Bush, R. K., Demain, J. G., Denning, D. W., Dixit, A., Fairs, A., et al. (2012). Fungi and allergic lower respiratory tract diseases. Journal of Allergy and Clinical Immunology, 129, 280–291.
Makra, L., Santa, T., Matyasovszky, I., Damialis, A., Karatzas, K., Bergmann, K. C., et al. (2010). Airborne pollen in three European cities: Detection of atmospheric circulation pathways by applying three-dimensional clustering of backward trajectories. Journal Geophysical Research. doi:10.1029/2010JD014743.
Mari, A., Schneider, P., Wally, V., Breitenbach, M., & Simon-Nobbe, B. (2003). Sensitization to fungi: Epidemiology, comparative skin tests, and IgE reactivity of fungal extracts. Clinical and Experimental Allergy, 33, 1429–1438.
Maya-Manzano, J., Fernández-Rodriguez, S., Hernández-Trejo, F., Díaz-Peres, G., Gonzalo-Garijo, Á., Silva-Palacios, I., et al. (2012). Seasonal Mediterranean pattern for airborne spores of Alternaria. Aerobiologia, 28, 515–525.
McMaster, G. S., & Wilhelm, W. W. (1997). Growing degree-days: One equation, two interpretations. Agricultural and Forest Meteorology, 87, 291–300.
Nilsson, S., & Persson, S. (1981). Tree pollen spectra in the Stockholm region (Sweden), 1973–1980. Grana, 20, 179–182.
Oerke, E. C., & Dehne, H. W. (2004). Safeguarding production-losses in major crops and the role of crop protection. Crop Protection, 23, 275–285.
Olesen, J. E., & Plauborg, F. (1995). MVTOOL version 1.10 for developing MARKVAND. SP Rep. 27, Danish Institute of Plant and Soil Science, Tjele.
Paldy, A., Bobvos, J., Fazekas, B., Manyoki, G., Malnasi, T., & Magyar, D. (2014). Characterisation of the pollen season by using climate specific pollen indicators. Central European Journal of Occupational and Environmental Medicine, 20, 199–214.
Pashley, C., Fairs, A., Edwards, R., Bailey, J., Corden, J., & Wardlaw, A. (2009). Reproducibility between counts of airborne allergenic pollen from two cities in the East Midlands, UK. Aerobiologia, 25, 249–263.
Poorter, H., van de Vijver, C. A. D. M., Boot, R. G. A., & Lambers, H. (1995). Growth and carbon economy of a fast-growing and a slow-growing grass species as dependent on nitrate supply. Pland and Soil, 171(2), 217–227.
Sadyś, M., Skjøth, C. A., & Kennedy, R. (2014). Back-trajectories show export of airborne fungal spores (Ganoderma sp.) from forests to agricultural and urban areas in England. Atmospheric Environment, 84, 88–99.
Sadyś, M., Skjøth, C. A., & Kennedy, R. (2015). Determination of Alternaria spp. habitats using 7-day volumetric spore trap. Hybrid Single Particle Lagrangian Integrated Trajectory model and geographic information system. Urban Climate, 14, 429–440.
Seifert, K., Morgan-Jones, G., Gams, W., & Kendrick, B. (2011). The genera of hyphomycetes. CBS Biodiversity Series no. 9: 1–997, CBS-KNAW Fungal Biodiversity Centre, Utrecht.
Šikoparija, B., Pejak-Šikoparija, T., Radišić, P., Smith, M., & Soldevilla, C. G. (2011). The effect of changes to the method of estimating the pollen count from aerobiological samples. Journal of Environmental Monitoring, 13, 384–390.
Simmons, E. G. (2007). Alternaria. An identification manual (1st ed.). CBS Biodiversity Series. Utrecht
Skjøth, C. A., Baker, P., Sadyś, M., & Adams-Groom, B. (2015). Adams-Groom B. (2015). Pollen from alder (Alnus sp.), birch (Betula sp.) and oak (Quercus sp.) in the UK originate from small woodlands. Urban Climate, 14, 414–428.
Skjøth, C. A., Smith, M., Šikoparija, B., Stach, A., Myszkowska, D., Kasprzyk, I., et al. (2010). A method for producing airborne pollen source inventories: An example of Ambrosia (ragweed) on the Pannonian Plain. Agricultural and Forest Meteorology, 150, 1203–1210.
Skjøth, C. A., Sommer, J., Frederiksen, L., & Gosewinkel Karlson, U. (2012). Crop harvest in Denmark and Central Europe contributes to the local load of airborne Alternaria spore concentrations in Copenhagen. Atmospheric Chemistry and Physics, 12, 11107–11123.
Smith, M., Jäger, S., Berger, U., Šikoparija, B., Hallsdottir, M., Sauliene, I., et al. (2014). Geographic and temporal variations in pollen exposure across Europe. Allergy, 69, 913–923.
Smith, M., Skjøth, C. A., Myszkowska, D., Uruska, A., Malgorzata, P., Stach, A., et al. (2008). Long-range transport of Ambrosia pollen to Poland. Agricultural and Forest Meteorology, 148, 1402–1411.
Stepalska, D., & Wolek, J. (2009). Intradiurnal periodicity of fungal spore concentrations (Alternaria, Botrytis, Cladosporium, Didymella, Ganoderma) in Cracow, Poland. Aerobiologia, 25, 333–340.
Su’udi, M., J-M, Park, Park, S. R., Hwang, D. J., Bae, D. J., Kim, S., & Ahn, I. P. (2013). Quantification of Alternaria brassicicola infection in the Arabidopsis thaliana and Brassica rapa subsp. pekinensis. Microbiology, 159, 1946–1955.
Suzuki, R. (2014) Hierarchical clustreing with p values via multiscale bootstrap resampling. CRAN.
Suzuki, R., & Shimodaira, H. (2006). Pvclust: an R package for assessing the uncertainty in hierarchical clustering. Bioinformatics, 22, 1540–1542.
R Core Team and Contributors Worldwide. (2015). The R Stats Package. https://stat.ethz.ch/R-manual/R-patched/library/stats/html/00Index.html.
TheLancet. (2008). Allergic rhinitis: Common, costly, and neglected. The Lancet, 371, 2057.
Thibaudon, M., Šikoparija, B., Oliver, G., Smith, M., & Skjøth, C. A. (2014). Ragweed pollen source inventory for France—the second largest centre of Ambrosia in Europe. Atmospheric Environment, 83, 62–71.
Toth, B., Csosz, M., Szabo-Hever, A., Simmons, E. G., Samson, R. A., & Varga, J. (2011). Alternaria hungarica sp., a minor foliar pathogen of wheat in Hungary. Mycologia, 103, 94–100.
Zhang, Y., Bielory, L., Cai, T., Mi, Z., & Georgopoulos, P. (2015). Predicting onset and duration of airborne allergenic pollen season in the United States. Atmospheric Environment, 103, 297–306.
Acknowledgments
Dr. C. A. Skjøth is supported by European Commission through a Marie Curie Career Integration Grant (Project ID CIG631745 and Acronym SUPREME). Dr. C. H. Pashley is supported by the Midlands Asthma and Allergy Research Association (MAARA) and the National Institute for Health Research Leicester Respiratory Biomedical Research Unit. Dr. S. Fernández-Rodríguez and Dr. R. Tormo-Molina are supported by Regional Government Science Foundation of the Junta de Extremadura through the two projects: PRI06A190, PRI BS10008. Dr. A. Damialis has been supported by the Research Committee of the Aristotle University of Thessaloniki (Excellence Fellowships of Postdoctoral Researchers, 2011). Dr. I. Kasprzyk and Dr. M. Jędryczkaare supported by National Science Centre Project No. N N305 321,737. The views expressed are those of the author(s) and not necessarily those of the European Commission, the NHS, the NIHR or the Department of Health.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Skjøth, C.A., Damialis, A., Belmonte, J. et al. Alternaria spores in the air across Europe: abundance, seasonality and relationships with climate, meteorology and local environment. Aerobiologia 32, 3–22 (2016). https://doi.org/10.1007/s10453-016-9426-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10453-016-9426-6