
Abstract
Australopiths are a group of early human ancestors that lived approximately 4 to 2 million years ago and are considered a key transitional form between apes and humans. Studying australopiths can help to understand the evolutionary processes that led to the emergence of humans and gain insights into the unique adaptations and characteristics that set humans apart from other primates. A bibliometric-based review of publications on australopiths contained in the Scopus database was conducted, analyzing approximately 2000 of them. The main authors, institutions, and countries researching this subject were identified, as well as their future development. The connections between authors, countries, and research topics were also analyzed through the detection of communities. The more frequent keywords in this subject are hominid, animal, human, South Africa, and Australopithecus afarensis. Four main research clusters were identified in the field of australopiths: palaeobiology, cranial evolution, locomotion, and mandible evolution and morphometry. The most important countries in terms of collaboration networks are South Africa, the UK, France, and Germany. Research on australopiths is ongoing, and new research clusters are expected to emerge, such as those focused on pre-australopiths and the molecular evolution and taxonomy of australopiths. Overall, this work provides a comprehensive overview of the state of research on australopiths and offers insights into the current direction of the field.
Résumé
Les australopithes sont un groupe de premiers ancêtres humains qui vivaient il y a environ 4 à 2 millions d'années et sont considérés comme une forme de transition essentielle entre les singes et les humains. L'étude des australopithes peut aider à comprendre les processus évolutifs qui ont conduit à l'émergence de l'homme et à mieux comprendre les adaptations et les caractéristiques uniques qui distinguent l'homme des autres primates. Une étude bibliométrique des publications sur les australopithes contenues dans la base de données Scopus a été réalisée, analysant environ 2000 d'entre elles. Les principaux auteurs, institutions et pays menant des recherches sur ce sujet ont été identifiés, ainsi que leur évolution future. Les liens entre les auteurs, les pays et les sujets de recherche ont également été analysés par la détection de communautés. Les mots-clés les plus fréquents dans ce domaine sont hominid, animal, human, South Africa et Australopithecus afarensis. Quatre groupes de recherche principaux ont été identifiés dans le domaine des australopithes: paléobiologie, évolution crânienne, locomotion, et évolution et morphométrie de la mandibule. Les pays les plus importants en termes de réseaux de collaboration sont l'Afrique du Sud, le Royaume-Uni, la France et l'Allemagne. Les recherches sur les australopithes se poursuivent et de nouveaux groupes de recherche devraient voir le jour, comme ceux qui se concentrent sur les pré-australopithes et sur l'évolution moléculaire et la taxonomie des australopithes. Dans l'ensemble, ce travail fournit une vue d'ensemble de l'état de la recherche sur les australopithes et donne un aperçu de l'orientation actuelle.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The family Hominidae (hominids, great apes) contains the subfamily Ponginae (Asian hominids, which has only one extant genus, Pongo -orangutans-), and the subfamily Homininae (hominines), to which belongs the tribe Gorillini (gorillas, chimpanzees, bonobos, and their evolutionary ancestors), the tribe Panini (the chimpanzee lineage), and the tribe Hominini (hominins) (Almecija et al., 2021). The tribe Hominini includes the subtribe Hominina (hominians), and this included the genus Homo, which is the only genus having a current species, and the questioned subtribe Australopithecina (australopiths). Within this last group included the genera Australopithecus, Paranthropus, and Kenyanthropus (Wood, 2010), as well as the earlier Ardipithecus, Orrorin, and Sahelanthropus. All these closely related species are now sometimes collectively termed australopiths or homininians (Mongle et al., 2019; Strait, 2010). The term australopithecine is now underused because this refers to a formal taxonomic group, the subfamily Australopithecinae, which is taxonomically unrecognized. In contrast, the term australopith does not refer to a taxonomic group and is a more appropriate term for all these species (Strait, 2010; Wood & Richmond, 2000). They are close relatives to current humans and, with the extant genus Homo, comprise the hominin clade. Members of Paranthropus are called the “robust australopiths,” while the term “gracile australopiths” is used to refer to other hominins such as Australopithecus, Kenyanthropus, or Homo (Wynn et al., 2020). Evaluating the hominin status of the pre-australopiths seems to be a very complex task, given that hominid fossils predating the emergence of australopiths are scarce. Ardipithecus ramidus, whose fossils were recovered from Ethiopia’s Afar Rift, has been considered its probable ancestor until recently, but today, this hypothesis is controversial. This hominin combined arboreal climbing with a bipedalism more primitive than that of Australopithecus (White et al., 2009).
Australopiths comprise ~ 10 Plio-Pleistocene hominin species who inhabited eastern, central, and southern Africa from ~ 4.2 million to ~ 1.0 million years ago (mya), and fossils have been found in Ethiopia, Chad, Kenya, Tanzania, and South Africa. They were highly diverse, having a large occurrence area, and they developed during periods of pronounced climate change (Strait, 2010). Although responses of hominins to climate change have been difficult to test, there are some well-documented cases. For instance, through high-resolution pollen data from Hadar, Ethiopia, it has been shown that Australopithecus afarensis successfully adapted to strong environmental variability between 3.4 and 2.9 mya. Such environmental change consisted of a large biome shift: up to 5 °C cooling and a 200- to 300-mm/year rainfall increase (Bonnefille et al., 2004).
Although australopiths do not belong to the genus Homo, they are hominins that share a suite of morphologies with Homo and several others with apes. Undoubtedly, they lacked several features that characterize current Homo, i.e., large brain and sophisticated tool use (Strait, 2010). Today, australopiths are considered an adaptive radiation of hominins that were terrestrial bipeds and able to better process hard-to-chew foods than their predecessors. Such conjunction of features allowed them to be successful and could have provided an appropriate setting for the emergence of Homo (Ward, 2018). Then, the evolutionary history of australopiths was influenced by their dietary adaptations (Paine & Daegling, 2023). Overall, australopiths comprise a grade (i.e., they show similar adaptations) rather than a clade of early hominin, as they do not constitute the descendant of a single common ancestor, while all these features induce to consider australopiths more related to Homo than with earlier apes or hominins. For instance, they are bipedal apes that show, to a greater or lesser degree, large molar and premolar teeth compared to the size of the incisors and canines (hyper-megadontia), and other associated modifications to their feeding apparatuses (Strait, 2010).
The genus Australopithecus was first identified in 1925, with the discovery of the fossil known as the Taung skull, which took place in South Africa. This species was named Australopithecus africanus, or “southern ape from Africa” by Dart (1925), and the name Australopithecus integrates the words austral, “from the south” and pithekos, “ape”. The Taung skull was identified as belonging to a new genus, Australopithecus, given its distinctive characteristics, including a foramen magnum (the hole through which the spinal cord exits the skull) that was tucked underneath the cranium. This suggested that this ape was able to stand and move upright on two feet (Tobias, 1965). The Taung skull also had molar teeth, reduced anterior teeth, and a shorter face compared to those of apes. These features led researchers to conclude that the Taung skull belonged to a hominin. Nevertheless, it shows a much smaller brain and larger postcanine teeth in comparison to that of Homo, as is for this that Dart classified it into a new genus (Ward, 2015). Australopiths were quickly credited with new abilities compared to those displayed by apes. For instance, the ability to throw was probably achieved at this stage of human evolution. While this ability is poorly developed in apes, anatomical studies suggest that the hand of A. afarensis was adapted to throw with precision and force (Isaac, 1987).
From the middle twentieth century, discoveries continued in southern Africa, and the australopiths range reached eastern Africa. For instance, in Olduvai Gorge, in Tanzania, a skull was discovered that undoubtedly belonged to a new genus of australopiths, which was initially classified as Zijanthropus boisiei (Leakey, 1959). This hominin is currently known as Paranthropus boisei, which has been found in locations ranging from Ethiopia to Malawi and exhibits a combination of primitive and derived features. Its anatomy is similar to that of Australopithecus, but it shows distinctive cranial and dental morphology, and for some morphological regions, such as the mandible and the mandibular dentition, the samples are relatively well typified (Richmond et al., 2020; Wood & Constantino, 2007). Fossils of this taxon show a relatively long and curved ulna, an oblique scapular spine, the absence of a third metacarpal styloid process, a gracile thumb metacarpal, and curved manual phalanges. It is thought that P. boisei had strong upper limb muscles, which allowed them to climb trees to a greater extent than earlier australopiths. The anatomy of the hand suggests that P. boisei, like earlier australopiths, had the manual dexterity to make stone tools, but lacked the robust thumb of Homo erectus (Richmond et al., 2020).
Some australopiths species are undoubtedly related to the Oldowan stone tool industry. The early tools belonging to this period were simple, usually made with one or a few flakes chipped off with another stone. Oldowan tools occurred during the Lower Paleolithic, dated to 2.9–1.7 mya in several areas of Africa. This technological industry was followed by the more sophisticated Acheulean industry, dated to 1.5–1.26 mya, which is associated with H. erectus (Semaw et al., 2020). The oldest Oldowan stone sites occurred within Ethiopia’s Afar Triangle, but recently, sites at Nyayanga (Kenya) have been found dated to 3.0–2.6 mya, and far away from this location. Hippopotamoid butchery proofs were found at such sites associated with Paranthropus sp. (Plummer et al., 2023).
Australopiths disappear after 1.4 mya. Probably, the last surviving species were P. boisei in eastern Africa and P. robustus in southern Africa (Strait, 2010). The extinction of P. boisei occurred when C4 grasslands dominated landscapes of the Eastern African Rift System. P. boisei was an herbivorous C4 specialist, and paradoxically, its demise coincided with habitats favorable to its dietary ecology. However, the long-term rise of C4 grasslands was disrupted by short periods of warmer temperatures that increased C3 vegetation by the Mid-Pleistocene Transition (1.3–0.7 mya). Such shortage of C4 grasslands increased dietary competition among abundant C4-feeders, and this fact could have influenced P. boisei demise (Quinn & Lepre, 2021).
Research on australopiths is timely since it is doubtless that some species belonging to this group constitute the ancestor of all Homo species, thus, knowing the initial stages of differentiation of the former with respect to the apes will help to understand the crucial steps of our evolution. This work aims to provide knowledge about global trends on australopiths research, the most important milestones, and the interrelationships between researchers and countries currently developing knowledge on this topic. Furthermore, bibliometric research can help to identify the most influential and highly cited works in the field, allowing researchers to focus their attention on the most impactful and relevant studies. This can be particularly useful for researchers who are new to the field, as it can help them to quickly become familiar with the most important works in the field and to understand the current state of knowledge on australopiths.
Materials and Methods
There are two main scientific databases, Scopus and WoS (Web of Science). Both provide access to a wide range of scholarly literature, including journal articles, conference proceedings, and other academic publications. Both databases cover a broad range of scientific disciplines, and they both use similar methods to index and organize the literature they contain. As a result, there is a significant overlap between the two databases in terms of the content they provide access to. However, Scopus and WoS do differ in some keyways. For example, Scopus has a larger database and covers more publication sources than WoS. Additionally, Scopus offers more advanced search and discovery tools, while WoS has a stronger focus on citation analysis and impact metrics. Some authors (Cascajares et al., 2021; Mongeon & Paul-Hus, 2016) have found that Scopus overlaps or matches WoS (Web of Science) in many scientific areas by more than 90% in the indexing of published papers. Scopus has been widely used in various fields for bibliometric studies, including engineering (Zapata-Sierra et al., 2021), medicine (Garrido-Cárdenas et al., 2018; 2019), biology (Salmerón-Manzano et al., 2020), and social science (Muyor-Rodríguez et al., 2019). As a result, most bibliometric research worldwide relies on Scopus or another database, which is why it was chosen for this study. The Scopus API was utilized to retrieve the data due to the large number of results. The methodologies previously developed by Montoya et al. (2018) and Cascajares et al. (2021) were followed. The query used in this study was ((TITLE-ABS-KEY (australopit*) OR TITLE-ABS-KEY (paranthropus))). The methodology used is outlined in Fig. 1. About keywords, or the relationship between countries or between authors, this has been carried out using the VosViewer SW, which applies the analysis of scientific communities or grouping by means of clusters (Salmeron-Manzano & Manzano-Agugliaro, 2019). Vosviewer is a software tool that is used to visualize and analyze bibliometric data. It is commonly used in research to identify patterns and trends in scientific literature, such as the most influential papers, authors, and institutions in a particular field. Vosviewer allows users to create maps and diagrams that represent the relationships between different papers, authors, and institutions and to identify clusters or communities within the data. It can also be used to perform a range of analyses, such as identifying trends over time, comparing different fields or disciplines, and analyzing the impact of research. Overall, Vosviewer is a valuable tool for understanding the structure and dynamics of scientific literature and for identifying key trends and patterns in research. The software tool Vosviewer has an algorithm for detecting communities in large networks using modularity. The modularity function was first introduced by Newman and Girvan, and Newman suggested using it to detect communities through the optimization of a modularity function (Waltman & Van Eck, 2013). Community detection is the process of identifying groups or clusters of related objects within a larger dataset. In many cases, these communities represent natural divisions within the data, and detecting them can provide valuable insights into the structure and organization of the underlying dataset. One common approach to community detection is to use the optimization of a modularity function. Modularity is a measure of the strength of a division within a network, and optimization of a modularity function involves finding the division that maximizes this measure. This approach can be useful for identifying communities because it can effectively capture the underlying structure of the network and identify groups of nodes that are densely connected to each other but less densely connected to nodes in other groups. In general, the optimization of a modularity function is a powerful tool for detecting communities in complex networks and can provide valuable insights into the underlying structure of the data.

Methodology
Results
Global Evolution Trend
From 1925 to 2022, there are 2246 studies published on australopiths. The research trend for this subject is depicted in Fig. 2. Notice that there was a steady increase in published papers from 1970 to 2016, with just over 1945 papers in total. After this period, the number of published works has remained relatively stable at around 65 per year. The year in which the discoveries of the various australopiths species take place are indicated in the graphic. A. africanus and P. robustus were the earliest discovered species, in 1925 and 1942, respectively. Between 1976 and 1999, several species were dated, and the more recent one, A. sediba, was discovered in 2010.

Worldwide temporal evolution of australopiths publications
Document Types and Languages
The various document types used for disseminating knowledge on australopiths are summarized in Table 1. Notice that the more frequent document type is an article, with more than 80% of the total document types. Article, book chapter, and review modalities represent more than 90% of the total. All these publications have been written in fourteen different languages: English (96%), French (3.5%), and others under 1% as German, Italian, Spanish, Chinese, Danish, Hungarian, Japanese, Polish, Portuguese, Russian, Swedish, or Ukrainian.
Global Subject Category
After analyzing results on australopiths according to the Scopus database categories (Fig. 3), it is concluded that the Social Sciences category was the more frequent one with 27% of the total, followed closely by Agricultural and Biological Sciences, with 25%. Other significant categories having relative relevance were Medicine (13%), Earth and Planetary Sciences and Multidisciplinary (9%), Biochemistry, Genetics and Molecular Biology (8%), Arts and Humanities and Environmental Science (3%). All other categories had a representation of less than 1%, including Immunology and Microbiology, Dentistry, Engineering, and Psychology.

Global subject category of australopiths publications
Distribution of Publications on Australophits by Countries
This topic has been researched by authors belonging to 63 countries. Figure 4 shows the countries that have published on this subject, and Fig. 4 shows the number of publications related to this topic. The USA stands out ~ with 1300 publications. The publications from authors of this country were focused on hominid*, *human*, animal*, Australopithecus, fossil*, evolution, A. afarensis, and morphology. The second most productive country was South Africa, with approximately 500 publications related to this subject. The most frequent keywords in these publications were hominid*, South Africa, Australopithecus, fossil*, animal*, *human, A. africanus, evolution, and Anatomy and Histology. The UK was in the third position with ~ 370 publications, and the main keywords were as in the previous country, but excluding A. africanus and South Africa and including Biological Evolution. France, with ~ 300 publications related to this subject was in the fourth position. The main keywords that French researchers used were hominid*, Australopithecus, fossil*, animal*, *human, evolution, South Africa, bone, and Pliocene. Germany holds the fifth position, with ~ 200 publications in this subject. For such works, the more frequent keywords were hominid*, animal*, fossil*, South Africa, human*, Australopithecus, Anatomy and Histology, evolution, morphology, and P. boisei. The previous countries are followed in intensity of research on this topic by Spain, Australia, Italy, and Kenya with 103, 95, 69, and 62 publications, respectively.

Worldwide australopiths number of publications by country until year 2022
The number of publications from the top 6 countries (those with more than 100 publications) from 1980 to 2022 is depicted in Fig. 5. This group includes the USA (1104), South Africa (430), the UK (337), France (268), Germany (193), and Spain (102). The USA maintained the greatest number of publications throughout this period. It is noticeable that from this group, the five remaining countries tended to stabilize at about 10–20 publications by year.

Temporal evolution on australopiths publications for top 6 countries
The analysis of the top 6 countries in accordance with the scientific categories in which their researchers have published is shown in Fig. 6. Notice that for the USA and France, Social Sciences is the main category for publishing research, followed by Agricultural and Biological Sciences, Medicine, Multidisciplinary, Biochemistry, Genetics and Molecular Biology, Earth and Planetary Sciences, Environmental Sciences, and Arts and Humanities. All these categories are shared by all researchers on the topic, but for investigators from South Africa, the UK, Germany, and Spain, the first one was Agricultural and Biological Sciences.

Distribution of publications on australopiths by scientific categories according to top 6 countries
Affiliations (Institutions)
In the previous section, the distribution of publications by country was analyzed. However, the investigations produced at Scopus-indexed institutions have not yet been discussed. Table 2 shows the 19 institutions with more than 50 publications, with 11 from the USA, 2 from France, the UK, and South Africa, and 1 from Germany and Kenya. The 4 most relevant keywords for these institutions show a close similarity in the research being conducted: hominid*, animal*, Australopithecus, and fossil*. Interestingly, South Africa was included in the top 4 main keywords only in the University of the Witwatersrand (Johannesburg, South Africa), while A. afarensis appears in the 4th position in the Arizona State University. The first position considering the intensity of research was for the University of the Witwatersrand, with 348 published papers and 4433 citations. This institution was followed by the CNRS (Centre National de la Recherche Scientifique, France), the Stony Brook University (US), and the Max-Planck-Institut für evolutionäre Anthropologie (Germany). The latter institutions have published 129, 125, and 118 documents on this topic and were cited by 2571, 3767, and 2347 papers, respectively. Interestingly, although the USA holds 11 institutions among the top 20 ones (ordered considering the number of papers related to the topic), the two first positions are for South Africa and France. The more relevant position for the UK institutions is hold by the University College (London), in 7th position, with 87 documents and 2224 citations.
Authors
Table 3 shows the researchers who accredit at least 30 papers on the subject. The lead author is L.R. Berger, from the University of the Witwatersrand (Johannesburg, South Africa), with 74 papers, followed by F.E. Grine, from the Stony Brook University (Stony Brook, US), with 58 papers. Interestingly, the highest h-index, i.e., the number of papers (h) published in a journal that has been cited at least h times, is held by B.A. Wood (h-index of 59), from the Center for the Advanced Study of Human Paleobiology (Washington, D.C., US), who published 39 works on australopiths. It is also interesting to note that authors from the University of the Witwatersrand occupy 3 of the top 4 positions in this table.
The collaboration network among authors who published more than 30 documents on this subject is shown in Fig. 7, so, clusters are centered around a main author and the connecting scholars are individuals who have collaborated with that individual. Table 4 summarizes the main authors detected in each cluster, which were built using the various keywords, and the number of authors that make up each of them.

Collaborative network of authors with over 30 publications on australopiths
There were 9 clusters identified, with the largest being the red cluster, which included 29 authors. The main authors in this cluster were R.J. Clarke and J.F Thackeray, both from the University of the Witwatersrand, with 49 and 48 papers each. Other relevant authors within this cluster were T.R. Pickering and K.J. Carlson, with 33 and 31 publications in the topic, both belonging to the previous institution. The second relevant cluster is depicted in light green, which is composed of 21 authors, in which stands out W.H. Kimbel (Institute of Human Origins) and C.O. Lovejoy (Kent State University), both from the USA, who performed 46 and 42 publications on this topic, respectively. Other notable authors within this cluster are H.M. McHenry (University of California, Davis, USA) with 41 published articles on the subject, White, T.D. (University of California, Berkley, USA) with 33 published articles, and G. Suwa (The University of Tokyo, Japan) with 31 publications on this topic. The third cluster, in blue, consists of 20 authors, in which the main authors are F.E. Grine (Stony Brook University) and M. Sponheimer (University of Colorado Boulder), both from the USA, who made 58 and 35 publications on this topic, respectively. Included in this cluster is also C.V. Ward (University of Missouri, Columbia, USA), with 33 papers related to the subject. The fourth cluster (yellow) consists of 19 authors, with B.D. Richmond (American Museum of Natural History, NY, USA) and B.A. Wood (Center for the Advanced Study of Human Paleobiology, WA, USA) as the central authors, having published 27 and 39 papers on this subject, respectively. The fifth cluster (purple) also includes 19 authors, with L.R. Berger (University of the Witwatersrand, Johannesburg, South Africa) and S.E. Churchill (Duke University, Durham, USA) as the leading authors, having published 74 and 29 papers on this subject, respectively. The sixth cluster is blue light-colored, and it is composed of 16 authors, being the most relevant author D.S. Strait (University of Johannesburg, South Africa), who developed 26 publications on this topic. The seventh cluster is colored in orange, and it contains 14 researchers, and its central author is B. Wood (Center for the Advanced Study of Human Paleobiology, WS, USA), who published 39 papers on australopiths. Note that this author is probably the same as the author of the aforementioned cluster four as he has the same surname, and his first initial and works are in the same institution. The eighth cluster is brown-colored and contains 13 authors and is led by P.V. Tobias (School of Clinical Medicine, Johannesburg, South Africa) and D. Falk (Florida State University, Tallahassee, USA), with 36 and 34 papers, respectively. Finally, the ninth group of authors is colored in pink and contains 8 authors whereas the central one is Y. Haile-Selassie (Institute of Human Origins, Tempe, USA), who published 26 papers on this topic.
Source (Journals)
Scientific journals play a pivotal role in the dissemination of research findings, serving as crucial platforms for sharing knowledge across the global scientific community. The selection of the right journal for publishing scientific articles holds paramount importance, as it determines the reach and impact of the research (Nievas-Soriano et al., 2023). The indexing of a journal in reputable databases, that is, Web of Science or Scopus, enhances its visibility and accessibility, ensuring that the research is discoverable by fellow researchers, practitioners, and policymakers (Marín-Buzón et al., 2021). One metric often used to gauge the influence of a journal is the Journal Citation Reports (JCR) impact factor, which measures the average number of citations received per article published in that journal over a specific time frame.
This factor provides insights into the journal’s significance within its field. Moreover, the categorization of journals based on subject areas assists researchers in locating articles relevant to their interests.
Table 5 details the main journals where more than 60 scientific articles related to australopiths research have been published. This table presents the following: N, the number of articles published on this topic, SJR (SCImago Journal Rank) developed by SCImago Lab and based on Scopus data, JCR (Journal Impact Factor) established by Clarivate Analytics and based on Web of Science, Category of JCR, and the rank in this category. Notice that the two journals with the highest number of publications fall within the Anthropology category, as expected. However, it is noteworthy that the following four belong to the Multidisciplinary category, and two of them are the top two within this lasts category. This highlights the significant general interest in this topic.
Keywords as Global Perspective
One of the goals of bibliometric studies is to analyze the keywords included in papers in order to establish relationships among various publications on a given topic and to identify scientific communities or clusters related to the specific subject. By extracting keywords from publications, we can get an overview of the most common keywords related to the australopith topic (Fig. 8). As expected, the search terms are among the main ones, and other head indexing terms were also identified, including Hominid, Hominidae, Human*, Animal*, Evolution, Female, Skull, Bone, Fossil*, Diet, Ape, Male, Australopithecus, and South Africa.

Word cloud of keywords in australopiths research
Table 6 presents the results of analyzing the keywords by country, and it details the five main keywords for the main countries that have conducted research on this topic. To create this table, search terms were not included. It is notable that the most prominent keywords include hominid*, human*, fossil, evolution, Paleolithic, and Pleistocene.
Scientific Communities or Clusters
The clusters were built through the various keywords, and each of them shares similar research trends, which allowed their classification. The documents published between 1925 and 2022 built four-colored clusters drawn in Fig. 9, and their main keywords collected are detailed in Table 7.

Network of keywords in australopiths publications
Cluster 1: Palaeobiology
The first cluster, “Palaeobiology,” is depicted in red and is built by 313 keywords, being the main ones are as follows: Hominid, South Africa, Hominin evolution, Pliocene, tooth, taphonomy, paleontology, paleoecology, carbon isotopes, Makapansgat, stratigraphy, and diet.
Hominin evolution is a prominent topic within this cluster. The establishment of hominin phylogeny is an open process, which is revived as new fossil discoveries take place. This happens especially when the existence of new species is confirmed, and therefore the paleontological discoveries made in the last decade have greatly expanded the known diversity of extinct taxa (Mongle et al., 2023). Currently, there are several uncertainties in hominin phylogeny, for instance, the phylogenetic relationships of Australopithecus spp. with their possible ancestor Ardipithecus (e.g., White et al., 2009), and their presumably descendant (Homo and Paranthropus) taxa (e.g., Villmoare et al., 2015) promote hot debates. Given that A. ramidus became extinct later than 4.3 mya, while Australopithecus appeared earlier than 4.2 mya or earlier, the two genera overlapped in time and, thus, a phyletic evolution seems infeasible between them (Bobe & Wood, 2022). On the other hand, a link between Australopithecus and other archaic hominins such as Orrorin tugenensis and Sahelanthropus tchadensis does not seem plausible, judging by their morphology (Strait, 2013). Lately, the study of new craniodental material led to the consideration of A. ramidus as being derived relative to Sahelanthropus and as the sister taxon of all later hominins (Mongle et al., 2019). At this point, new fossils are expected to help stabilize the origin of Australopithecus.
A remarkable attempt to build australopith’s phylogeny was effected through parsimony analyses on sixty craniodental and mandibular characters (Strait et al., 1997). Authors have found evidence for a Paranthropus clade, with Australopithecus being paraphyletic and specimens attributed to A. afarensis being reclassified as P. africanus, which is believed to be the sister of later hominids. In addition, Paranthropus was positioned as the sister of Homo and A. africanus as the sister of the Homo + Paranthropus clade. However, it was noted that certain aspects of hominid evolution remain enigmatic.
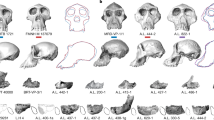
In addition to the previous attempt, there have been other ones to establish the phylogeny of hominids (e.g., Mongle et al., 2019, 2023; Strait & Grine, 2004; Villmoare, 2018). Figure 10 shows a cladogram based on a recently published phylogenetic analysis using currently available morphological craniodental data (Mongle et al., 2023; Fig. 5: Results of iteration 4 Bayesian inference analysis). In this analysis, A. sediba was reconstructed in a polytomy at the Homo + Paranthropus node, while H. habilis and H. rudolfensis were recovered in a polytomy at the base of the genus Homo. A. africanus and A. platyops (syn. Kenyanthropus platyops) were reconstructed together as sister taxa to a clade containing A. sediba, Paranthropus, and Homo. Such matrix was consistent with previous ones (e.g., Mongle et al., 2019; Strait & Grine, 2004). The main conclusion was that probably, A. sediba constitutes the end of a lineage sharing a common ancestor with the first Homo spp.

Cladogram representing a recently published phylogeny for hominins (adapted from Mongle et al., 2023)
While for Paranthropus, there is a wide consensus that it constitutes a distinct monophyletic genus (e.g., Strait et al., 1997; Wood & Constantino, 2007); the phylogenetic relationships between Australopithecus taxa are the subject of deep controversy. Careful interpretations of the fossil record of Australopithecus accurately recognize five species, which is supported by the presence of uniquely derived, or unique combinations of shared derived, morphological features: A. afarensis, A. africanus, A. garhi, A. platyops, and A. sediba (e.g., Alemseged, 2023; Berger et al., 2010).
One challenge in classifying fossil hominid material is sexual dimorphism, as the morphological variation in the hominid fossil record exhibits greater sexual dimorphism than exists among modern humans (Brace, 1973; Johanson & White, 1979).
Some time ago, it was considered that A. anamensis and A. afarensis were separate species in an anagenetic series (e.g., Kimbel et al., 2006), and it was believed that the former was the oldest species of Australopithecus, which was discovered in Kanapoi and Allia Bay (Kenya). Bipedalism for this species was established through the study of a tibia (Leakey et al., 1995). A. anamensis remains showed a mosaic of primitive and derived features that led to considering this species as an ancestor to A. afarensis, and given that more than one early hominid species could have coexisted at any one time, this discovery suggested that A. ramidus was a sister species to A. anamensis and all later hominids (Leakey, 1995). Later, the study of a lower first deciduous molar (dm1) allowed, confirming that A. anamensis was morphologically intermediate between A. ramidus and A. afarensis, and age estimation was ~ 4 mya (Leakey et al., 1998; Ward et al., 1999). However, today A. anamensis and A. afarensis are interpreted as one single taxon because the changes noted over time in the cranium, mandible, and dentition in A. anamensis to A. afarensis are interpreted as an intraspecies temporal change. Therefore, the former would represent a synonym of the latter (Alemseged, 2023; Kimbel, et al., 2006).
Traditionally, A. afarensis (from the Afar region of Ethiopia) has been considered one primitive australopith species and the common ancestor of A. africanus, A. platyops, A. garhi, Paranthropus, and H. habilis (Alemseged, 2023; Johanson & White, 1979). However, the stratigraphic analysis necessary for accurate dating of karst cave sites in South Africa is highly complex, so this phylogenetic assumption may be uncertain. In this regard, the study of Australopithecus fossils from Sterkfontein (South Africa) led other authors to question that A. africanus descended from A. afarensis, and that there was a coexistence of these two species, which suggests a highly complex family tree in human evolution (Granger et al., 2022).
From time to time, new species of such hominins have appeared, yielding new insights about the origins of early Homo. For instance, the discovery of ~ 2.5 mya hominins remains in Hata beds of Ethiopia’s Middle Awash allowed the recognition of a new Australopithecus species: A. garhi. It is believed that this species is descended from A. afarensis and was considered one of the possible ancestor candidates of Homo since it presents intermediate traits between Australopithecus and Homo: humanlike humeral/femoral ratio and an apelike upper arm-to-lower arm ratio (Asfaw et al., 1999).
A controversial case is related to the taxonomy of A. platyops. At first, it was found enough morphological distance between it and A. afarensis to reflect a different dietary adaptation, thus, justifying the naming of a new genus, i.e., Kenyanthropus (Leakey et al., 2001). However, subsequent discoveries remain, i.e., a maxilla, reduced morphological differences with Australopithecus, for instance, between K. platyops and A. deyiremeda. Thus, it was proposed to rename K. platyops as A. platyops (Alemseged, 2023).
Regarding the paleoecology of australopiths, environmental reconstructions suggest that Australopithecus had a high environmental adaptability. For example, in Kanapoi (Rift Valley, Kenya), the habitats of Australopithecus were characterized by a seasonal climate, abundant inland waters, and diverse fauna, and a competition for resources probably occurred (Dumouchel et al., 2021). Overall, Australopithecus was adapted to a diversity of habitats such as forests, savannahs, and grassland environments such as that of Kanapoi, than to more humid and forest-rich habitats such as those of the Omo Mursi Formation (southwestern Ethiopia) (e.g., Dumouchel et al., 2021; Fillion et al., 2022). Such adaptability may have favored to early Australopithecus populations to disperse and adapt to a variety of ecological niches, which allowed several speciation events. Then, the later Australopithecus, i.e., A. africanus, A. sediba, and A. garhi and early Paranthropus and Homo, successfully exploited resources from various environments (Alemseged, 2023). An environmental shift to more prevalent aridity took place after 3 mya, and this might have triggered the emergence of several new Australopithecus species, Homo, and Paranthropus, and perhaps the extinction of A. afarensis (Alemseged, 2023; Alemseged et al., 2020).
Other studies ascribed to this cluster are devoted to the relative size of the posterior teeth to diet. All australopiths had jaws with densely enameled teeth, as those of anatomically modern humans (AMH), but they showed also ape-like canines and molars, which has long been interpreted as an adaptation an omnivorous behavior (Kay, 1985). Relative tooth size was modeled for forest and savanna primates and Homo and was also calculated for Australopithecus using posterior maxillary area sums and size. It was found that these hominins were adapted to a diet that included heavily chewed grains, roots, and scavenged animal protein (Wolpoff, 1973). In the same year, it was suggested that robust and gracile australopiths had different diets, being Paranthropus vegetarians and Australopithecus omnivorous, given that their range was in different adaptive zones (Kay, 1985; Wallace, 1973). The dietary habits of A. robustus (P. robustus) were also assessed through the 13C/12C ratio analysis of structural carbonate in tooth enamel, as its robust masticatory apparatus and dental features suggest a specialized dietary niche. The results for P. robustus showed a more generalized or omnivorous diet compared to other australopiths (Lee-Thorp et al., 1994). Later on, stable carbon isotope analysis of A. africanus from Makapansgat Limeworks, South Africa, indicated that this hominin ate huge amounts 13C-rich foods such as grasses or animals that ate these plants, or both. Thus, it was suggested that probably such hominins consumed animal foods before the development of stone tools by Homo species (Sponheimer & Lee-Thorp, 1999). Furthermore, studies through carbon isotopes of A. africanus from Sterkfontein (~ 2.5–2.0 mya) showed that this hominin intensively exploited the savanna food web (Van der Merwe et al., 2003). Interestingly, the cranial and dental features of australopiths between 4.4 and 2.3 mya indicated that the dietary capabilities of such earliest hominids evolved in a wide variety of habitats, making them well adapted for using the various resources associated with climatic fluctuations (Teaford & Ungar, 2000).
Dental microwear texture was used a long time ago as an adequate tool to identify the diets of gracile and robust australopiths (Henry et al., 2012; Scott et al., 2005). It was hypothesized that differences in occlusal microwear indicate that the diets of both Paranthropus and Australopithecus were qualitatively different (Grine, 1986). However, this assumption was rebutted, arguing that A. africanus dental microwear texture is more anisotropic but also more variable in anisotropy than that of P. robustus. This implies that A. africanus ate more tough foods, while P. robustus consumed more hard and brittle items, but both had similar diets (Scott et al., 2005). It is likely that stone tools helped such food behaviors, and the oldest direct evidence of their manufacture (2.6 and 2.5 mya) comes from Gona (Ethiopia) where identification of several cut-marked bones indicates stone tool use and stone-tool-assisted consumption of ungulates by A. afarensis (McPherron et al., 2010). Some authors indicate that probably, both gracile and robust australopiths consumed the same foods, but based on dental differences, it is though that P. robustus would have required higher energy to break down most usually consumed foods, given by orthognathism and larger chewing muscles, as well as a thicker enamel for resisting such chewing energy (Berthaume & Kupczik, 2021). Although doubts for any carnivory behavior after the appearance of Homo erectus have been exposed (Barr et al., 2022), as above mentioned, hippopotamoid butchery proofs were found associated with Paranthropus sp., while a tooth from this species possessed carbon isotopic values indicative of a C4 foods-rich diet. Then, the processing of diverse foods, including megafauna, may have been quite usual for Paranthropus sp. (Plummer et al., 2023).
The diet of A. sediba was carefully assessed. The combined analysis of plant phytoliths from dental calculus, stable carbon isotope, and dental microwear texture revealed that this hominin consumed C3 foods in preference to widely available C4 resources diet. Thus, the diet of this species was similar to that of A. ramidus and modern savanna chimpanzees (Henry et al., 2012).
Cluster 2: Cranial Evolution
The second cluster, “Cranial evolution,” is depicted in green and is built by 228 keywords, being the main ones are as follows: human, animal, skull, physical anthropology, paleodontology, haplorhini, allometry, craniometry, and cranial capacity. Early research that can be included in this cluster is related to the redetermination of the endocranial volume of the Taung skull, which was set to 405 cm3, instead of the previously reported 525 cm3, and for an adult, the volume was estimated to have 440 cm3. This value, added to other australopith measures, lowers the average to 442 cm3 and increased the statistically significant differences from both robust australopithecines and the Olduvai Gorge hominid, which belongs to the Homo genus (H. erectus) (Holloway, 1970).
In all cases, the cranial capacity of australopiths was much less than those of Homo species. For instance, A. africanus had a mean cranial volume of 451 cm3, although some individuals reached 515 cm3. Paranthropus had a mean cranial volume of 479.4 cm3, and the range was from 410 (P. boisei) to 530 cm3 (P. robustus and P. boisei), while for Paranthropus aethiopicus, there were intermediate values. It needs to be considered that the cranial volume of modern gorillas is 484 cm3, while for an AMH, it is ~ 1450 cm3 (Falk et al., 2000). Although the cranial volume of all australopiths is closer to that of the great apes than to that of the AMH, the brain morphology of A. africanus appears more human-like than that of Paranthropus in terms of overall frontal and temporal lobe shape. Then, the former could have been ancestral to Homo (Falk et al., 2000).
It needs to be considered that the taxonomic separation of australopiths based on cranial and facial features is questionable due to the limited number of samples available for analysis. For example, A. boisei (syn. P. boisei), which was originally described based on a cranium found in Olduvai Gorge, Tanzania in 1959, is known for its robust masticatory apparatus. However, some fossil remains discovered in Konso, Ethiopia, including a cranium and associated mandible, showed significant variation within this species. Moreover, these fossils extend the known geographical range of P. boisei, suggesting that P. boisei and H. erectus may have coexisted (Suwa et al., 1997). This highlights the importance of considering multiple samples when examining the taxonomy of ancient hominids.
A. sediba was selected to study the brain enlargements and organization from australopiths to Homo. Reports on their craniodental and postcranial skeletons from ~ 1.95 to 1.78 mya led to believe that this hominin descended from A. africanus and shares more derived features with early Homo than any other australopith species (Berger et al., 2010). A. sediba brain shows a gradual evolution in the orbitofrontal region from Australopithecus to Homo, though the small endocast volume of A. sediba suggests that brain enlargement occurred after this reorganization (Carlson et al., 2011).
Cluster 3: Locomotion
The third cluster, in blue, was labeled “Locomotion,” given significant keywords performing such cluster, which contains 134 items: A. afarensis, anatomy and histology, biomechanics, locomotion, physiology, pelvis, posture, and pelvic bones. Within this grouping, the most cited publications are related to the locomotion of australopiths. Research on this subject started on Early Pleistocene hominids from East and South Africa and was based primarily on the foot and leg bones from Olduvai Gorge (Preuschoft, 1971). Early research on the biomechanics of Australopithecus locomotion was focused on the pelvis and femur, and it was concluded that the gait pattern of AMH and the two recognized forms of Australopithecus were indistinguishable. This suggests that the differences in morphology between Australopithecus and AMH having biomechanical significance appear to be related to the combination of a fully bipedal striding gait with different degrees of encephalization, rather than to differences in the gait pattern itself (Lovejoy et al., 1973).
Important research within this cluster is related to the bodily proportions of australopiths, which support phylogenetic interpretations of their evolution. In this regard, the skeleton of A. afarensis (AL 288–1; “Lucy”) recovered in 1974 from the Hadar area of Ethiopia (Johanson & Taieb, 1976) allowed direct assessment of body size, limb proportions, and skeletal allometry of this species, which has been widely researched to study the biomechanics of australopiths. The body proportions of A. afarensis, as represented by the skeleton of Lucy (AL 288–1), are suitable for bipedalism but differ from those of modern Homo. The reduced relative stride length of Lucy may suggest that bipedal locomotion required more energy for A. afarensis compared to AMH (Jungers, 1982). Overall, there is a wide consensus among researchers that the earliest recognized hominins exhibited adaptations indicating habitual bipedalism. Such behavior was attributed also to their supposed ancestor, A. ramidus, which is thought to combined arboreal climbing with primitive bipedalism (White et al., 2009).
Probably, australopiths inhabited the oases of the African savannah where trees remained and some other plant foods, and traditionally, this fact has been considered the cause of why they started descending from the trees to walking upright (Reynolds et al., 2011). The ability to stand upright would have favored their survival, as they can travel long distances to feed as scavengers, to collect fallen fruit, insects, and many other resources far from the trees (Cordain et al., 1998). This latter behavior would have provided the ability for migration to exploit other less explored food resources, as well as to anticipate predators in the savannah, given their ability to look up (Shillington, 2019).
However, the various features of bipedal locomotor behavior in fossil hominins constitute a hot topic, for instance, the degree of arboreal behavior among the various australopiths and the locomotor variability across different species. All studies of the locomotor skeleton of australopiths indicated both arboreal climbing/suspension and terrestrial bipedalism. Evidence supports the idea that A. afarensis, represented by the postcranial skeleton and footprints found in the Laetoli Beds of Tanzania, had adaptations for both tree climbing and terrestrial bipedalism. Such bipedalism differed from that of AMH in that it involved less extension at the hip and knee than occurs in modern humans and only limited transfer of weight onto the medial part of the ball of the foot (Stern & Susman, 1983). Sexual differences in locomotor behavior, likely due to marked size dimorphism, were also observed, with males possibly engaging in less arboreal activity and more terrestrial bipedalism (Stern & Susman, 1983; Susman et al., 1984). This suggests that A. afarensis had a unique combination of characteristics that allowed for flexible movement in both trees and on the ground, although it was argued that the evidence presented for arboreal behavior was not conclusive (Ward, 2002). For A. afarensis, there were clear proofs for bipedal locomotion, as the foot and other evidence from the lower limb provide, and most authors believed that this hominin had an arboreal behavior, as the gorilla-like scapula and long and curved manual phalanges suggest (e.g., Alemseged et al., 2006).
A nearly complete wrist and hand of an adult female A. sediba from South Africa (dating to 1.977 mya) exhibit both Australopithecus-like features, such as a strong flexor apparatus for arboreal locomotion, and Homo-like features, such as a long thumb and short fingers associated with precision gripping and potentially stone tool production. This suggests that A. sediba had a unique combination of adaptations that may have allowed for flexible movement in both trees and on the ground, as well as the ability to produce and use tools. Thus, A. sediba was considered a candidate for early stone tool use and production (Kivell et al., 2011). Moreover, through the study of an articulated partial foot and ankle and other bones from this hominin recovered in the Malapa site, South Africa, it was deduced that it may have practiced a unique form of bipedalism and some degree of arboreality (Zipfel et al., 2011).
Recently, a study was developed on hip joint loading conditions of AMH, baboons, and fossil hominins attributed to A. africanus, P. robustus, and Homo. The conclusions were that australopiths had human-like bipedal locomotion, although other features of their musculoskeletal systems exhibit ape-like characteristics (Ryan et al., 2018). It is believed that bipedalism evolved in an arboreal context, associated to a foraging strategy, which was concluded through the study of the positional behavior and terrestrially in a savanna-mosaic community of chimpanzees (Pan troglodytes schweinfurthii) (Drummond-Clarke et al., 2022). Such bipedalism has been linked to an adaptation for locomotion on flexible branches. In this regard, orangutans react to branch flexibility like humans running on springy tracks by increasing knee and hip extension, while all other apes do the reverse. Then, more than an innovation, human bipedalism seems to be an exploitation of a locomotor behavior retained from the common great ape ancestor (Thorpe et al., 2007).
However, strong controversy about the origin of bipedalism in australopiths exists. There are investigations whose results diverge from this last hypothesis, arguing that bipedalism in australopiths evolved as a dry-habitat foraging strategy. To test this hypothesis, the measure of femoral shaft obliquity in humans, forest chimpanzees, and dry-habitat (Semliki) chimpanzees was recently investigated (Hunt et al., 2021). It was found strong evidence that such human obliquity develops in response to forces generated during bipedal positional behavior. Given their close phylogenetic proximity, obliquity among the Semliki chimpanzees could develop via the same mechanism, thus, suggesting that Semliki chimpanzees engage in bipedalism more often than forest chimpanzees. It was found that dry-habitat chimpanzee femora were intermediate between forest chimpanzees and humans. Furthermore, human femoral obliquity was compared with the values of six australopiths, and angles of australopiths were not significantly greater than those of humans. Thus, femoral obliquity among dry-habitat chimpanzees is consistent with the hypothesis that bipedalism evolved as a dry-habitat foraging strategy (Hunt et al., 2021).
Cluster 4: Mandible Evolution and Morphometry
The fourth cluster, in yellow, includes 134 keywords, and among these standout H. habilis, mandible, morphometry, body size, dentition, odontometry, and teeth; thus, it was named “Mandible evolution and morphometry.” Research on this topic includes the study of the body of such hominins and the use of mandibles for determining phylogeny. Clearly, this cluster partially overlaps with the first one, given some keywords belonging to the latter, as tooth and diet. Therefore, some information given here complements that of the first cluster.
A. ramidus was selected as a model with Paleobiological implications for understanding the earliest stages of human evolution through the study of its teeth, including associated maxillary and mandibular sets. Interestingly, the postcanine megadontia of Australopithecus is absent in A. ramidus. For the latter, an omnivorous/frugivorous niche was suggested through the dental morphology and wear pattern, while the canine/lower third premolar complex indicates a reduction of canine size and honing capacity early in hominid evolution, probably directed by a selection toward the male upper canine (Suwa et al., 2009).
The compact bone distribution and biomechanics of early hominid mandibles help to understand the differentiation in hominin’s evolution. The distribution of compact bone in the postcanine mandibular corpus of A. africanus and P. robustus was analyzed using computed tomography to examine its effects on the biomechanical properties of these bones (Daegling & Grine, 1991). The compact bone contours were used to calculate cross-sectional biomechanical properties, and it was found that the mechanical properties of the jaws of A. africanus and P. robustus are distinct: A. africanus had less cortical bone than P. robustus, suggesting that the phenomenon of mandibular “robusticity” in australopiths may be due to postcanine megadontia and/or reduced canine size (Daegling & Grine, 1991). Traditionally, the craniofacial morphology of australophits has been considered a dietary adaptation for feeding on either small, hard objects, or on large volumes of food (Grine, 1986). Therefore, the evolution of australopith craniofacial form has been attributed to the loads applied to the premolars during feeding. However, some researchers have proposed that certain aspects of australopith craniofacial morphology may have evolved in response to the ingestion and initial processing of large, mechanically protected food items like nuts and seeds. These foods may have been important sources of sustenance for australopiths during times of food scarcity (Strait et al., 2009). In this regard, using a combination of physical testing and finite element analysis, it has been formulated the hypothesis that australopith occlusal morphology may not represent adaptations for inducing fractures in large, hard food items, but rather for resisting fractures in the tooth crown (Berthaume et al., 2010).
Recently, geometric morphometrics and finite element analysis have been used to analyze mandible morphology and chewing biomechanics for establishing the variation within eight fossil hominin species. By relating stress and shape variables and grouping fossil individuals into broad categories based on the hardness of their diet, researchers were able to conclude that some hominins previously thought to consume hard foods, such as those belonging to the Paranthropus clade, actually relied on softer foods. This finding is consistent with other studies using microwear and stable isotope analyses (Marcé-Nogué et al., 2020).
Morphometry and body size are also research belonging to this cluster. Traditionally, evolutionary models argued that body size increased from Australopithecus to early Homo, while sexual dimorphism in body size decreased (e.g., McHenry & Coffing, 2000; Pilbeam & Gould, 1974). Through the relationship between various measures of skeletal size and body mass in modern ape and human specimens of known body weight, the male and female average weights of some australopiths were determined by McHenry (1992), who calculated much higher weights for australopith males. Later, Grabowski et al. (2015) stated that many early hominids had smaller bodies than previously thought and provided evidence that large modern human-like size first appeared at least 3 to 3.5 mya in some individuals of A. afarensis. Through equations based on a large (n = 220) sample of modern humans of known body masses, Grabowski et al. (2015) estimated male and female average weights: A. afarensis 49.5 and 31.2 kg, A. africanus 38.9 and 25.8 kg, P. robustus (syn. A. robustus) 32.3 and 24 kg, and A. boisei 45.1 and 30.9 kg, thus questioning the widely accepted evolutionary model of the increase in body size from Australopithecus to the first Homo. Besides body-size dimorphism, canine-size dimorphism is a feature of all australopiths species (e.g., McHenry, 1996). Both morphologic characteristics have been interpreted as a mating system involving a low degree of male-male competition and social structure, especially in A. afarensis, as the body weight of males is estimated almost twice than that of females. This level of dimorphism is similar to that of chimpanzees and gorillas, indicating intense male-male competition and polygyny (Plavcan et al., 2005), although there is some discrepancy in this hypothesis, arguing failures in methods that are used to estimate body size (e.g., Reno & Lovejoy, 2015). Interesting findings derived from the mandible study defining the range for A. afarensis, whose occurrence was first ascribed to the eastern Rift Valley from Tanzania to Ethiopia. The distribution area of such australopith was enlarged through the discovery of a mandible associated with a fauna biochronologically estimated to be 3.0–3.5 mya old in Chad, in the region of Bahr el Ghazal (2,500 km west of the Rift Valley) (Brunet et al., 1995).
Concerning mandibular evolution, important research is related to masticatory muscles. Australopithecus and Paranthropus had these muscles like most primates. Conversely, Homo species showed smaller masticatory muscles. The evolutionary process for the reduction of the masticatory apparatus was in parallel to an encephalization in early Homo. The gene encoding the main myosin heavy chain (MYH) expressed in certain muscles was inactivated after the human and chimpanzee lineages diverged, approximately 2.4 million years ago. This mutation predates the emergence of AMH morphology (Stedman et al., 2004). This suggests that the inactivation of this gene may have been a factor in the evolution of human anatomy.
Finally, the study of mandibular postcanine dentition for australopiths from the Shungura formation, Ethiopia, yielded evidence about an incipient robust taxon, A. aethiopicus, that emerged at ~ 2.9 mya, which was largely plesiomorphic: its dentitions had a larger size than that of A. robustus (Suwa et al., 1996).
Evolution of Recent Australopith Research
The trend in the evolution of australopith research from the year 2000 until present is depicted in Fig. 11. The legend shows that there is a color gradation from purple (the oldest keywords) to yellow (the more recent ones). Notice that the oldest terms for australopith* research were human, physical anthropology, comparative study, and paleodontology. It is likely that such terms reflect that research on this subject was focused mainly on the knowledge of differences in morphology among australopiths and with respect to that of Homo spp. In the intermediate period, the more outstanding keywords were hominid, biomechanics, locomotion, female, and A. afarensis, which reflects an interest in the biomechanics of the locomotion. Recently, the more frequent search term was anatomy and physiology, which indicates a growing interest in such features of australopith beings (Cofran, 2019).

Evolution trend of keywords in australopiths publications
Collaboration Network of Countries Involved in Australopith Research
Figure 12 shows the collaborative network among the various countries that perform research on australopiths. Table 8 details the countries belonging to each grouping and the leader country in each cluster. The countries most central to this network of collaboration are the USA, Kenya, Australia, France, and the UK. Notice the importance relative to the USA, although practically, it performs research in an independent way. The largest cluster is led by Kenya, with 8 collaborative countries, but without significant weight in research on this topic. This cluster was closely followed by those led by Australia, France, and the UK, with 7, 6, and 5 attached countries. In this figure, it can be appreciated that the proximity of the investigations are performed between South African and UK researchers.

Countries’ network collaboration in australopiths publications
Conclusions
Australopiths as a research topic proved to be of high scientific interest as demonstrated by the fact that it has been investigated by researchers from 63 different countries. The analysis of the scientific literature included in the Scopus database about australopiths showed a continuous increase from 1925 to 2022, with 2246 studies published on this topic. Then, a continuous increase from 1970 to 2016 takes place with just over 1945 papers. After this period, published works were stabilized at approximately 65 each year. The research on australopiths shows nine collaborative clusters among western countries and other ones from Africa and Australia. The main terms indexed when developing research on australopiths are hominid, animal, human, South Africa, and Australopithecus afarensis. From the analysis of the keywords of all these documents, four clusters were found (i) “Palaeobiology,” having as main keywords tooth, taphonomy, paleontology, paleoecology, diet, and carbon isotopes; (ii) “Cranial evolution,” having keywords as the skull, physical anthropology, paleodontology, craniometry, and cranial capacity; (iii) “Locomotion,” in which significant keywords were anatomy and physiology, biomechanics, locomotion, pelvis, posture, and pelvic bones; (iv) “Mandible evolution and morphometry,” in which the outstanding keywords were mandible, morphometry, body size, dentition, odontometry, and teeth. These clusters demonstrate the diversity and depth of research being conducted on australopiths and highlight the importance of these early human ancestors in our understanding of human evolution. Future research in this field has the potential to continue shedding light on the lives and adaptations of these interesting hominins and to further our understanding of our own evolutionary history. Overall, the research on australopiths is essential for our understanding of human evolution and for providing context and insights into the unique characteristics and adaptations that make us human. By continuing to study and learn about these early human ancestors, knowledge of our own evolutionary history will be expanded and placed in the natural world.
Data Availability
The data have been extracted from the Scopus scientific database and are available to all those who have access to it. However, they will be made available upon reasoned request.
References
Alemseged, Z., Wynn, J. G., Geraads, D., Reed, D., Andrew Barr, W., Bobe, R., ... & Mohan, J. (2020). Fossils from Mille-Logya, Afar, Ethiopia, elucidate the link between Pliocene environmental changes and Homo origins. Nature communications, 11, 2480. https://doi.org/10.1038/s41467-020-16060-8
Alemseged, Z. (2023). Reappraising the palaeobiology of Australopithecus. Nature, 617, 45–54. https://doi.org/10.1038/s41586-023-05957-1
Alemseged, Z., Spoor, F., Kimbel, W. H., Bobe, R., Geraads, D., Reed, D., & Wynn, J. G. (2006). A juvenile early hominin skeleton from Dikika, Ethiopia. Nature, 443, 296–301. https://doi.org/10.1038/nature05047
Almécija, S., Hammond, A. S., Thompson, N. E., Pugh, K. D., Moyà-Solà, S., & Alba, D. M. (2021). Fossil apes and human evolution. Science, 372, eabb4363. https://doi.org/10.1126/science.abb4363
Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S., & Suwa, G. (1999). Australopithecus garhi: A new species of early hominid from Ethiopia. Science, 284, 629–635. https://doi.org/10.1126/science.284.5414.629
Barr, W. A., Pobiner, B., Rowan, J., Du, A., & Faith, J. T. (2022). No sustained increase in zooarchaeological evidence for carnivory after the appearance of Homo erectus. Proceedings of the National Academy of Sciences, 119, e2115540119. https://doi.org/10.1073/pnas.211554011
Berger, L. R., De Ruiter, D. J., Churchill, S. E., Schmid, P., Carlson, K. J., Dirks, P. H., & Kibii, J. M. (2010). Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science, 328, 195–204. https://doi.org/10.1126/science.1184944
Berthaume, M., Grosse, I. R., Patel, N. D., Strait, D. S., Wood, S., & Richmond, B. G. (2010). The effect of early hominin occlusal morphology on the fracturing of hard food items. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 293, 594–606. https://doi.org/10.1002/ar.21130
Berthaume, M. A., & Kupczik, K. (2021). Molar biomechanical function in South African hominins Australopithecus africanus and Paranthropus robustus. Interface Focus, 11, 20200085. https://doi.org/10.1098/rsfs.2020.0085
Bobe, R., & Wood, B. (2022). Estimating origination times from the early hominin fossil record. Evolutionary Anthropology: Issues, News, and Reviews, 31, 92–102. https://doi.org/10.1002/evan.21928
Bonnefille, R., Potts, R., Chalié, F., Jolly, D., & Peyron, O. (2004). High-resolution vegetation and climate change associated with Pliocene Australopithecus afarensis. Proceedings of the National Academy of Sciences, 101, 12125–12129. https://doi.org/10.1073/pnas.040170910
Brace, C. L. (1973). Sexual dimorphism in human evolution. Yearbook of Physical Anthropology, 16, 31–49.
Brunet, M., Beauvilain, A., Coppens, Y., Heintz, E., Moutaye, A. H., & Pilbeam, D. (1995). The first australopithecine 2,500 kilometres west of the Rift Valley (Chad). Nature, 378, 273–275. https://doi.org/10.1038/378273a0
Carlson, K. J., Stout, D., Jashashvili, T., De Ruiter, D. J., Tafforeau, P., Carlson, K., & Berger, L. R. (2011). The endocast of MH1, Australopithecus sediba. Science, 333, 1402–1407. https://doi.org/10.1126/science.12039
Cascajares, M., Alcayde, A., Salmerón-Manzano, E., & Manzano-Agugliaro, F. (2021). The bibliometric literature on Scopus and WoS: The medicine and environmental sciences categories as case of study. International Journal of Environmental Research and Public Health, 18, 5851. https://doi.org/10.3390/ijerph18115851
Cofran, Z. (2019). Brain size growth in Australopithecus. Journal of Human Evolution, 130, 72–82. https://doi.org/10.1016/j.jhevol.2019.02.006
Cordain, L., Gotshall, R. W., & Eaton, S. B. (1998). Physical activity, energy expenditure and fitness: An evolutionary perspective. International Journal of Sports Medicine, 19, 328–335. https://doi.org/10.1055/s-2007-971926
Daegling, D. J., & Grine, F. E. (1991). Compact bone distribution and biomechanics of early hominid mandibles. American Journal of Physical Anthropology, 86, 321–339. https://doi.org/10.1002/ajpa.1330860302
Dart, R. A. (1925). Australopithecus africanus: The man-ape of South Africa. Nature, 115, 195–199. https://doi.org/10.1038/115195a0
Drummond-Clarke, R. C., Kivell, T. L., Sarringhaus, L., Stewart, F. A., Humle, T., & Piel, A. K. (2022). Wild chimpanzee behavior suggests that a savanna-mosaic habitat did not support the emergence of hominin terrestrial bipedalism. Science Advances, 8, eadd9752. https://doi.org/10.1126/sciadv.add975
Dumouchel, L., Bobe, R., Wynn, J. G., & Barr, W. A. (2021). The environments of Australopithecus anamensis at Allia Bay, Kenya: A multiproxy analysis of early Pliocene Bovidae. Journal of Human Evolution, 151, 102928. https://doi.org/10.1016/j.jhevol.2020.102928
Falk, D., Redmond, J. C., Jr., Guyer, J., Conroy, C., Recheis, W., Weber, G. W., & Seidler, H. (2000). Early hominid brain evolution: A new look at old endocasts. Journal of Human Evolution, 38, 695–717. https://doi.org/10.1006/jhev.1999.0378
Fillion, E. N., Harrison, T., & Kwekason, A. (2022). A nonanalog Pliocene ungulate community at Laetoli with implications for the paleoecology of Australopithecus afarensis. Journal of Human Evolution, 167, 103182. https://doi.org/10.1016/j.jhevol.2022.103182
Garrido-Cardenas, J. A., Cebrián-Carmona, J., González-Cerón, L., Manzano-Agugliaro, F., & Mesa-Valle, C. (2019). Analysis of global research on malaria and Plasmodium vivax. International Journal of Environmental Research and Public Health, 16, 1928. https://doi.org/10.3390/ijerph16111928
Garrido-Cardenas, J. A., Mesa-Valle, C., & Manzano-Agugliaro, F. (2018). Human parasitology worldwide research. Parasitology, 145, 699–712. https://doi.org/10.1017/S0031182017001718
Grabowski, M., Hatala, K. G., Jungers, W. L., & Richmond, B. G. (2015). Body mass estimates of hominin fossils and the evolution of human body size. Journal of Human Evolution, 85, 75–93. https://doi.org/10.1016/j.jhevol.2015.05.005
Granger, D. E., Stratford, D., Bruxelles, L., Gibbon, R. J., Clarke, R. J., & Kuman, K. (2022). Cosmogenic nuclide dating of Australopithecus at Sterkfontein, South Africa. Proceedings of the National Academy of Sciences, 119, e2123516119. https://doi.org/10.1073/pnas.2123516119
Grine, F. E. (1986). Dental evidence for dietary differences in Australopithecus and Paranthropus: A quantitative analysis of permanent molar microwear. Journal of Human Evolution, 15, 783–822. https://doi.org/10.1016/S0047-2484(86)80010-0
Henry, A. G., Ungar, P. S., Passey, B. H., Sponheimer, M., Rossouw, L., Bamford, M., ... & Berger, L. (2012). The diet of Australopithecus sediba. Nature, 487, 90–93. https://doi.org/10.1038/nature11185
Holloway, R. L. (1970). Australopithecine endocast (Taung specimen, 1924): A new volume determination. Science, 168, 966–968. https://doi.org/10.1126/science.168.3934.966
Hunt, K. D., Dunevant, S. E., Yohler, R. M., & Carlson, K. J. (2021). Femoral bicondylar angles among dry-habitat chimpanzees (Pan troglodytes schweinfurthii) resemble those of humans: Implications for knee function, australopith sexual dimorphism, and the evolution of bipedalism. Journal of Anthropological Research, 77, 303–337. https://doi.org/10.1086/715398
Isaac, B. (1987). Throwing and human evolution. African Archaeological Review, 5, 3–17. https://doi.org/10.1007/BF01117078
Johanson, D. C., & Taieb, M. (1976). Plio-pleistocene hominid discoveries in Hadar, Ethiopia. Nature, 260, 293–297. https://doi.org/10.1038/260293a0
Johanson, D. C., & White, T. D. (1979). A systematic assessment of early African hominids. Science, 203, 321–330. https://doi.org/10.1126/science.104384
Jungers, W. L. (1982). Lucy’s limbs: Skeletal allometry and locomotion in Australopithecus afarensis. Nature, 297, 676–678. https://doi.org/10.1038/297676a0
Kay, R. F. (1985). Dental evidence for the diet of Australopithecus. Annual Review of Anthropology, 14(1), 315–341.
Kimbel, W. H., Lockwood, C. A., Ward, C. V., Leakey, M. G., Rak, Y., & Johanson, D. C. (2006). Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. Journal of Human Evolution, 51, 134–152. https://doi.org/10.1016/j.jhevol.2006.02.003
Kivell, T. L., Kibii, J. M., Churchill, S. E., Schmid, P., & Berger, L. R. (2011). Australopithecus sediba hand demonstrates mosaic evolution of locomotor and manipulative abilities. Science, 333, 1411–1417. https://doi.org/10.1126/science.1202625
Leakey, L. S. (1959). A new fossil skull from Olduvai. Nature, 184, 491–493.
Leakey, M. G., Feibel, C. S., McDougall, I., & Walker, A. (1995). New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature, 376, 565–571. https://doi.org/10.1038/376565a0
Leakey, M. G., Feibel, C. S., McDougall, I., Ward, C., & Walker, A. (1998). New specimens and confirmation of an early age for Australopithecus anamensis. Nature, 393, 62–66. https://doi.org/10.1038/29972
Leakey, M., Spoor, F., Brown, F., Gathogo, P. N., Kiarie, C., Leakey, L. N., & McDougall, I. (2001). New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature, 410, 433–440. https://doi.org/10.1038/35068500
Lee-Thorp, J. A., van der Merwe, N. J., & Brain, C. K. (1994). Diet of Australopithecus robustus at Swartkrans from stable carbon isotopic analysis. Journal of Human Evolution, 27, 361–372. https://doi.org/10.1006/jhev.1994.1050
Lovejoy, C. O., Heiple, K. G., & Burstein, A. H. (1973). The gait of Australopithecus. American Journal of Physical Anthropology, 38, 757–779. https://doi.org/10.1002/ajpa.1330380315
Marcé-Nogué, J., Püschel, T. A., Daasch, A., & Kaiser, T. M. (2020). Broad-scale morpho-functional traits of the mandible suggest no hard food adaptation in the hominin lineage. Scientific Reports, 10(1), 1–11. https://doi.org/10.1038/s41598-020-63739-5
Marín-Buzón, C., Pérez-Romero, A., Lopez-Castro, J. L., Ben Jerbania, I., & Manzano-Agugliaro, F. (2021). Photogrammetry as a new scientific tool in archaeology: Worldwide research trends. Sustainability, 13, 5319. https://doi.org/10.3390/su13095319
McHenry, H. M. in The archaeology of human ancestry: Power, sex and tradition (eds Shennan, S. & Steele, J.) 82–99 (Routledge, 1996).
McHenry, H. M. (1992). Body size and proportions in early hominids. American Journal of Physical Anthropology, 87, 407–431. https://doi.org/10.1002/ajpa.1330870404
McHenry, H. M., & Coffing, K. (2000). Australopithecus to Homo: Transformations in body and mind. Annual Review of Anthropology, 29, 125–146. https://doi.org/10.1016/j.jhevol.2015.05.005
McPherron, S. P., Alemseged, Z., Marean, C. W., Wynn, J. G., Reed, D., Geraads, D., ... & Béarat, H. A. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466, 857–860. https://doi.org/10.1038/nature09248
Mongeon, P., & Paul-Hus, A. (2016). The journal coverage of Web of Science and Scopus: A comparative analysis. Scientometrics, 106, 213–228. https://doi.org/10.1007/s11192-015-1765-5
Mongle, C. S., Strait, D. S., & Grine, F. E. (2019). Expanded character sampling underscores phylogenetic stability of Ardipithecus ramidus as a basal hominin. Journal of Human Evolution, 131, 28–39. https://doi.org/10.1016/j.jhevol.2019.03.006
Mongle, C. S., Strait, D. S., & Grine, F. E. (2023). An updated analysis of hominin phylogeny with an emphasis on re-evaluating the phylogenetic relationships of Australopithecus sediba. Journal of Human Evolution, 175, 103311. https://doi.org/10.1016/j.jhevol.2022.103311
Montoya, F. G., Alcayde, A., Baños, R., & Manzano-Agugliaro, F. (2018). A fast method for identifying worldwide scientific collaborations using the Scopus database. Telematics and Informatics, 35, 168–185. https://doi.org/10.1016/j.tele.2017.10.010
Muyor-Rodriguez, J., Manzano-Agugliaro, F., & Garrido-Cardenas, J. A. (2019). The state of global research on social work and disability. Social Work in Health Care, 58, 839–853. https://doi.org/10.1080/00981389.2019.1659904
Nievas-Soriano, B. J., Martín-Latorre, M. D. M., Martín-González, M., Manzano-Agugliaro, F., & Castro-Luna, G. (2023). Worldwide research trends on bronchiolitis in pediatrics. Pediatric Pulmonology, 58, 2189–2203. https://doi.org/10.1002/ppul.26453
Paine, O. C., & Daegling, D. J. (2023). The game of models: Dietary reconstruction in human evolution. Journal of Human Evolution, 174, 103295. https://doi.org/10.1016/j.jhevol.2022.103295
Pilbeam, D., & Gould, S. J. (1974). Size and scaling in human evolution: Homo sapiens is a peculiar large primate; however, all australopithecines are versions of the “same” animal. Science, 186, 892–901. https://doi.org/10.1126/science.186.4167.89
Plavcan, J. M., Lockwood, C. A., Kimbel, W. H., Lague, M. R., & Harmon, E. H. (2005). Sexual dimorphism in Australopithecus afarensis revisited: How strong is the case for a human-like pattern of dimorphism? Journal of Human Evolution, 48, 313–320. https://doi.org/10.1016/j.jhevol.2004.09.006
Plummer, T. W., Oliver, J. S., Finestone, E. M., Ditchfield, P. W., Bishop, L. C., Blumenthal, S. A., ... & Potts, R. (2023). Expanded geographic distribution and dietary strategies of the earliest Oldowan hominins and Paranthropus. Science, 379(6632), 561–566. https://doi.org/10.1126/science.abo7452
Preuschoft, H. (1971). Body posture and mode of locomotion in early Pleistocene hominids. Folia Primatologica, 14, 209–240.
Quinn, R. L., & Lepre, C. J. (2021). Contracting eastern African C4 grasslands during the extinction of Paranthropus boisei. Scientific Reports, 11, 1–10. https://doi.org/10.1038/s41598-021-86642-z
Reno, P. L., & Lovejoy, C. O. (2015). From Lucy to Kadanuumuu: Balanced analyses of Australopithecus afarensis assemblages confirm only moderate skeletal dimorphism. PeerJ, 3, e925. https://doi.org/10.7717/peerj.925
Reynolds, S. C., Bailey, G. N., & King, G. C. (2011). Landscapes and their relation to hominin habitats: Case studies from Australopithecus sites in eastern and southern Africa. Journal of Human Evolution, 60, 281–298. https://doi.org/10.1016/j.jhevol.2010.10.001
Richmond, B. G., Green, D. J., Lague, M. R., Chirchir, H., Behrensmeyer, A. K., Bobe, R., ... & Braun, D. R. (2020). The upper limb of Paranthropus boisei from Ileret, Kenya. Journal of Human Evolution, 141, 102727. https://doi.org/10.1016/j.jhevol.2019.102727
Ryan, T. M., Carlson, K. J., Gordon, A. D., Jablonski, N., Shaw, C. N., & Stock, J. T. (2018). Human-like hip joint loading in Australopithecus africanus and Paranthropus robustus. Journal of Human Evolution, 121, 12–24. https://doi.org/10.1016/j.jhevol.2018.03.008
Salmerón-Manzano, E., Garrido-Cardenas, J. A., & Manzano-Agugliaro, F. (2020). Worldwide research trends on medicinal plants. International Journal of Environmental Research and Public Health, 17, 3376. https://doi.org/10.3390/ijerph17103376
Salmerón-Manzano, E., & Manzano-Agugliaro, F. (2019). The role of smart contracts in sustainability: Worldwide research trends. Sustainability, 11, 3049. https://doi.org/10.3390/su11113049
Scott, R. S., Ungar, P. S., Bergstrom, T. S., Brown, C. A., Grine, F. E., Teaford, M. F., & Walker, A. (2005). Dental microwear texture analysis shows within-species diet variability in fossil hominins. Nature, 436, 693–695. https://doi.org/10.1038/nature03822
Semaw, S., Rogers, M. J., Simpson, S. W., Levin, N. E., Quade, J., Dunbar, N., ... & Everett, M. (2020). Co-occurrence of Acheulian and Oldowan artifacts with Homo erectus cranial fossils from Gona, Afar, Ethiopia. Science Advances, 6(10), eaaw4694. https://doi.org/10.1126/sciadv.aaw4694
Shillington, K. (2019). Early prehistory of Africa. In History of Africa (pp. 9–22). Palgrave, London.
Sponheimer, M., & Lee-Thorp, J. A. (1999). Isotopic evidence for the diet of an early hominid, Australopithecus africanus. Science, 283, 368–370. https://doi.org/10.1126/science.283.5400.368
Stedman, H. H., Kozyak, B. W., Nelson, A., Thesier, D. M., Su, L. T., Low, D. W., ... & Mitchell, M. A. (2004). Myosin gene mutation correlates with anatomical changes in the human lineage. Nature, 428, 415–418. https://doi.org/10.1038/nature02358
Stern, J. T., Jr., & Susman, R. L. (1983). The locomotor anatomy of Australopithecus afarensis. American Journal of Physical Anthropology, 60, 279–317. https://doi.org/10.1002/ajpa.1330600302
Strait, D. S., Weber, G. W., Neubauer, S., Chalk, J., Richmond, B. G., Lucas, P. W., … & Smith, A. L. (2009). The feeding biomechanics and dietary ecology of Australopithecus africanus. Proceedings of the National Academy of Sciences, 106(7), 2124–2129. https://doi.org/10.1073/pnas.08087301
Strait, D. S. (2010). The evolutionary history of the australopiths. Evolution: Education and Outreach, 3, 341–352. https://doi.org/10.1007/s12052-010-0249-6
Strait, D. S. (2013). The biogeographic implications of early hominin phylogeny. In K. E. Reed, J. G. Fleagle, & R. E. Leakey (Eds.), The Paleobiology of Australopithecus (pp. 183–191). Springer.
Strait, D. S., & Grine, F. E. (2004). Inferring hominoid and early hominid phylogeny using craniodental characters: The role of fossil taxa. Journal of Human Evolution, 47, 399–452. https://doi.org/10.1016/j.jhevol.2004.08.008
Strait, D. S., Grine, F. E., & Moniz, M. A. (1997). A reappraisal of early hominid phylogeny. Journal of Human Evolution, 32(1), 17–82. https://doi.org/10.1006/jhev.1996.0097
Susman, R. L., Stern, J. T., & Jungers, W. L. (1984). Arboreality and bipedality in the Hadar hominids. Folia Primatologica, 43, 113–156. https://doi.org/10.1159/000156176
Suwa, G., Asfaw, B., Beyene, Y., White, T. D., Katoh, S., Nagaoka, S., ... & WoldeGabriel, G. (1997). The first skull of Australopithecus boisei. Nature, 389, 489–492. https://doi.org/10.1038/39037
Suwa, G., Kono, R. T., Simpson, S. W., Asfaw, B., Lovejoy, C. O., & White, T. D. (2009). Paleobiological implications of the Ardipithecus ramidus dentition. Science, 326, 69–99. https://doi.org/10.1126/science.11758
Suwa, G., White, T. D., & Howell, F. C. (1996). Mandibular postcanine dentition from the Shungura Formation, Ethiopia: Crown morphology, taxonomic allocations, and Plio-Pleistocene hominid evolution. American Journal of Physical Anthropology: The Official Publication of the American Association of Physical Anthropologists, 101, 247–282. https://doi.org/10.1002/(SICI)1096-8644(199610)101:2%3c247::AID-AJPA9%3e3.0.CO;2-Z
Teaford, M. F., & Ungar, P. S. (2000). Diet and the evolution of the earliest human ancestors. Proceedings of the National Academy of Sciences, 97, 13506–13511. https://doi.org/10.1073/pnas.260368897
Thorpe, S. K., Holder, R. L., & Crompton, R. H. (2007). Origin of human bipedalism as an adaptation for locomotion on flexible branches. Science, 316, 1328–1331. https://doi.org/10.1126/science.114079
Tobias, P. V. (1965). Australopithecus, Homo habilis, tool-using and tool-making. The South African Archaeological Bulletin, 20, 167–192. https://doi.org/10.2307/3887824
Van Der Merwe, N. J., Thackeray, J. F., Lee-Thorp, J. A., & Luyt, J. (2003). The carbon isotope ecology and diet of Australopithecus africanus at Sterkfontein, South Africa. Journal of Human Evolution, 44, 581–597. https://doi.org/10.1016/S0047-2484(03)00050-2
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, C. J., DiMaggio, E. N., Rowan, J., ... & Reed, K. E. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science, 347, 1352–1355. https://doi.org/10.1126/science.aaa13
Villmoare, B. (2018). Early Homo and the role of the genus in paleoanthropology. American Journal of Physical Anthropology, 165, 72–89. https://doi.org/10.1002/ajpa.23387
Wallace, J. A. (1973). Tooth chipping in the australopithecines. Nature, 244, 117–118.
Waltman, L., & Van Eck, N. J. (2013). A smart local moving algorithm for large-scale modularity-based community detection. The European Physical Journal B, 86, 471. https://doi.org/10.1140/epjb/e2013-40829-0
Ward, C. V. (2002). Interpreting the posture and locomotion of Australopithecus afarensis: Where do we stand? American Journal of Physical Anthropology: The Official Publication of the American Association of Physical Anthropologists, 119, 185–215. https://doi.org/10.1002/ajpa.10185
Ward, C. V. (2015). Australopithecines. In Basics in human evolution (pp. 129–142). Academic Press. https://doi.org/10.1016/B978-0-12-802652-6.00010-4
Ward, C. V. (2018). Australopithecine/australopith. The International Encyclopedia of Biological Anthropology, 1–10. https://doi.org/10.1002/9781118584538.ieba0043
Ward, C., Leakey, M., & Walker, A. (1999). The new hominid species Australopithecus anamensis. Evolutionary Anthropology: Issues, News, and Reviews: Issues, News, and Reviews, 7(6), 197–205. https://doi.org/10.1002/(SICI)1520-6505(1999)7:6%3c197::AID-EVAN4%3e3.0.CO;2-T
White, T. D., Asfaw, B., Beyene, Y., Haile-Selassie, Y., Lovejoy, C. O., Suwa, G., & WoldeGabriel, G. (2009). Ardipithecus ramidus and the paleobiology of early hominids. Science, 326, 64–86. https://doi.org/10.1126/science.1175802
Wolpoff, M. H. (1973). Posterior tooth size, body size, and diet in South African gracile australopithecines. American Journal of Physical Anthropology, 39, 375–393. https://doi.org/10.1002/ajpa.1330390306
Wood, B. A. (2010). Systematics, taxonomy, and phylogenetics: Ordering life, past and present. In A companion to biological anthropology (pp. 56–73). https://doi.org/10.1002/9781444320039
Wood, B., & Richmond, B. G. (2000). Human evolution: Taxonomy and paleobiology. Journal of Anatomy, 197, 19–60. https://doi.org/10.1046/j.1469-7580.2000.19710019
Wood, B., & Constantino, P. (2007). Paranthropus boisei: Fifty years of evidence and analysis. American Journal of Physical Anthropology: The Official Publication of the American Association of Physical Anthropologists, 134, 106–132. https://doi.org/10.1002/ajpa.20732
Wynn, J. G., Alemseged, Z., Bobe, R., Grine, F. E., Negash, E. W., & Sponheimer, M. (2020). Isotopic evidence for the timing of the dietary shift toward C4 foods in eastern African Paranthropus. Proceedings of the National Academy of Sciences, 117, 21978–21984. https://doi.org/10.1073/pnas.2006221117
Zapata-Sierra, A., Cascajares, M., Alcayde, A., & Manzano-Agugliaro, F. (2021). Worldwide research trends on desalination. Desalination, 519, 115305. https://doi.org/10.1016/j.desal.2021.115305
Zipfel, B., DeSilva, J. M., Kidd, R. S., Carlson, K. J., Churchill, S. E., & Berger, L. R. (2011). The foot and ankle of Australopithecus sediba. Science, 333, 1417–1420. https://doi.org/10.1126/science.12027
Funding
Funding for open access publishing: Universidad de Almería/CBUA.
Author information
Authors and Affiliations
Contributions
José Luis Guil-Guerrero: conceptualization, investigation, writing—review and editing, project administration, formal analysis, supervision, and validation. Francisco Manzano-Agugliaro: writing—review and editing, software, formal analysis, supervision, investigation, and visualization.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Guil-Guerrero, J.L., Manzano-Agugliaro, F. Worldwide Research on Australopiths. Afr Archaeol Rev (2024). https://doi.org/10.1007/s10437-024-09580-x
Accepted:
Published:
DOI: https://doi.org/10.1007/s10437-024-09580-x