
Abstract
Sponges and corals from the Bartonian marly formations of the Pamplona Basin (South Pyrenean area, Navarre) are described for the first time. The fossiliferous levels correspond respectively to flood-influenced delta-front (Ardanatz Sandstone) and restricted outer-platform (Ilundain Marls Fm.) environments. The fossil sponges exhibit diagenetic fragmentation, but they are often complete specimens. The skeleton appears partially or totally replaced by calcite and/or in some cases large crystals of celestite. Celestite forms relatively early during diagenesis in a dysoxic environment. Neomorphic fibrous quartzine-lutecine spherulites are also present. Hexactinellids and lithistids occur, but the former predominate. The associations include the species Laocoetis samueli, Guettardiscyphia thiolati and/or Pleuroguettardia iberica, cf. Rhizocheton robustus, and two lithistids indet. Corals are present only in the Ardanatz Sandstone. The fossil skeletons are composed of large neomorphic sparry calcite crystals. The assigned species are Stylocoenia taurinensis, Astrocoenia octopartita, Ceratotrochus bodellei, Placosmiliopsis bilobatus, and Desmophyllum castellolense. The sponge and coral taxa are similar to those previously described from other contemporaneous geological formations of the Pyrenean realm. The Pamplona Basin assemblages appear less diverse than those of the Bartonian of the eastern South Pyrenean area, more similar to that of the Eocene of Biarritz (Aquitanian Basin). This lower diversity is not due to a lower-resolution sampling but to taphonomic bias and/or paleoecological differences. The dominance of hexactinellids, erect morphologies, and sedimentological and micropaleontological data show that the sponge communities lived in deep shelfal waters. The corals, mainly associated with levels with high terrigenous content, seem reworked from shallower and more proximal environments.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Middle Eocene marly marine formations are widespread throughout Western Europe, and some of these formations have been well known since the 19th century as the home to important fossil sites: for example, the Bartonian marly sites of Barton-on-Sea in England and the Paris Basin (Deshayes 1864; Pacaud and Le Renard 1995; West 2002). Further southwards, a significant number of French geologists and paleontologists (d’Archiac 1846, 1850; Rouault 1850; de Bouillé 1876; Boussac 1911) carried out pioneering studies of nummulitic faunas in the marly formations of the North Pyrenean area (Aquitaine Basin) around the localities of Biarritz (Miarritze), Bayonne (Baiona), Dax, and Pau. Similar marly formations also occur in the South Pyrenean area, but comparable studies have been largely restricted to the eastern (Catalonian and Aragonian) part of the region (Álvarez-Pérez 1993; Álvarez et al. 1994; Abad 2001; Serra-Kiel et al. 2003; Costa et al. 2012).
Interestingly, the South Pyrenean marly formations have long been known to extend westwards as far as the city of Pamplona (Navarre), as shown by the early publication of the French geologist Carez (1881). This author regarded the Pamplona marls as identical to the “marls of Serpula spirulaea”, which were in turn a continuation of the La Canal de Berdún marls (Zaragoza). Carez mentioned the occurrence of abundant specimens of a small bivalve south of Pamplona, which he attributed to a new species (“une Plicatule inédite”), Plicatula pamplonensis, later re-classified as Dimya pamplonensis (Carez 1881) by Calzada and Astibia (1996). However, it was not until many years later that Ruiz de Gaona (1947) and Ruiz de Gaona and Colom (1950) completed a more comprehensive study of these marly formations describing their foraminiferal content and concluding that they were Bartonian in age; these authors also mentioned the occurrence of important macrofossil associations near the village of Ardanatz and other localities near Pamplona. Since then, except for some local determinations (Puigdefàbregas 1975; Alameda et al. 1993), a micropaleontological revision of these formations has been pending. Regarding the macrofauna, with the exception of a brief note on Dimya pamplonensis (Calzada and Astibia 1996) and several studies on fossil turtles and sirenian remains (Astibia et al. 1999, 2005, 2006; de Buffrénil et al. 2008), nothing has been published yet. Consequently, the Middle Eocene marly marine formations of the Pamplona Basin are not well known from a paleontological point of view.
The purpose of this paper is to fill this gap in regional knowledge and thereby provide new information for a better characterization and comprehension of the widespread Western European Middle Eocene marly formations and their marine invertebrate biodiversity. To this end, we present a first study on the invertebrate fossil associations of the Pamplona marly formations and, more specifically, their sponge and coral faunas. In order to obtain their paleobathymetric and paleoenvironmental preferences, the sedimentological features of the host rocks and the benthic foraminifera of key levels were also analyzed.
Geological setting
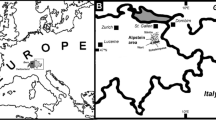
In Eocene times the South Pyrenean zone was a NW-opening marine gulf (Fig. 1). Until early Bartonian times the basin corresponded to a deep-water trough, but siliciclastic sediments derived from the uplifting and denuding Pyrenees progressively filled in the basin, so that large fluvio-deltaic systems became widespread by Bartonian and Priabonian times (Plaziat 1981; Pujalte et al. 2002; Barnolas et al. 2004). The Bartonian stage is well recorded in the Pamplona Basin by alternating marly and sandy lithostratigraphic units (Fig. 1). The fossil assemblages studied herein come from two successive units: the uppermost part of the Ardanatz Sandstone, exposed at the localities of Ardanatz and Aranguren, about 6 km southeast of Pamplona; and the Ilundain Marls from the locality of Arraitza, about 20 km southwest of Pamplona. One additional specimen comes from Badoztain, near Ardanatz, but geologically located in the Ilundain Marls.

Above simplified geological map of the Pamplona Basin, southwestern Pyrenees (see Eocene paleogeographic location in inset; Pa Pamplona Basin); boxed localities correspond to the studied fossil sites. Below synthetic log of the Eocene marly succession in the Pamplona Basin, with its constituent lithostratigraphic units and their depositional environments; the circle shows the interval containing the fossil sites. Adapted from Astibia et al. (2005)
The Ardanatz Sandstone outcrops at Ardanatz and Aranguren were thoroughly studied by Astibia et al. (1999, 2005, 2006). Silty marls are dominant. They contain interbedded tabular sandstones with abundant carbonaceous plant remains, vertical and horizontal burrows and some bioclasts (e.g., corals and Nummulites). On the basis of their sedimentological features, the sandstones were interpreted as deposits from non-confined, high-concentration turbulent flows. Given that the sandstone beds are arranged in several thickening- and coarsening-upward sequences, they were further interpreted as sandy hyperpycnites accumulating in flood-generated delta-front lobes (sensu Mutti et al. 2000, 2003).
The Arraitza section is located approximately 100 m above the base of the Ilundain Formation. The section is characterized by thick tabular levels of alternating dark and light grey marls with gradational boundaries. Their siliciclastic content mainly consists of clay, whereas the carbonate content mainly derives from skeletal remains.
On the basis of their stratigraphic position and the associations of Nummulites, Astibia et al. (1999, 2005) determined an early Bartonian age (SBZ17 of Serra-Kiel et al. 1998) of the Ardanatz–Aranguren deposits, but no comparable data are available for the Ilundain Marls in Arraitza. Therefore, determining the stratigraphic and paleoenvironmental relationships between the Ardanatz–Aranguren and Arraitza fossils is not straightforward. The Ardanatz Sandstone clearly grade upwards into the Ilundain Marls, but both lithostratigraphic units also interfinger laterally (Fig. 1). As the transition between the Ardanatz Sandstone and the Ilundain Marls is probably time-transgressive, the stratigraphic position of the Arraitza fossils in the latter unit does not necessarily imply that they are younger than the fossils in the former; in fact, the opposite could even be possible. In the absence of a sound stratigraphic correlation between the sites, only a sedimentological approach can be attempted here. In Arraitza the Ardanatz Sandstone is typified by thinner and finer-grained sandstone beds than in its type area. This suggests that the Arraitza area corresponded to a distal or marginal area of the Ardanatz delta front. It is hence reasonable to conclude that there was a continuous facies transition from the most proximal Ardanatz delta-front facies into the most distal/marginal outer-platform Ilundain Marl facies, through the intermediate fine-grained and thin-bedded sandstone facies of the Ardanatz Sandstone in Arraitza.
Materials and methods
The Ardanatz Sandstone fossils were collected from three outcrops located south and east of the village of Ardanatz (AD1, AD2, and AD3) and another outcrop (AG1) northwest of Aranguren, all of which display approximately the same stratigraphic interval at the top of the unit (Fig. 2). Unfortunately, a precise bed correlation was hampered due to the discontinuous character of the delta-front sandstone lobes and the occurrence of vegetation between outcrops. However, a tentative stratigraphic correlation suggests that a similarly patchy nature can also be assumed for the fossil-bearing marly levels, as their number and relative position vary between sections (Fig. 2). East of Arraitza two fossil-bearing levels (ARR1.1 and ARR1.2) of the same outcrop were sampled, which embrace approximately 40 m of the Ilundain Marls.

Tentative correlation of the sections studied in the Ardanatz–Aranguren area, showing the stratigraphic position of fossil-bearing levels (sandstone beds are not to scale; from Astibia 1979, unpubl. data)
A semi-quantitative study of foraminifera was carried out from level ARR1.1 in Arraitza and from levels AD2.1 and AD3.1 in Ardanatz from the fraction > 100 μm. The percentages of planktic foraminiferal taxa (P/B ratio) and several proxies for benthic foraminiferal analysis, such as the percentages of agglutinated and infaunal taxa and the species richness (number of species), were calculated. Paleobathymetric inferences were based on the P/B ratio and the occurrence and abundance of depth-related benthic species, and on the morphogroup analysis (Jones and Charnock 1985; Murray et al. 2011).
Some macrofossil specimens were cleaned with potassium hydroxide (90 %, flakes QP); other samples were partially dissolved in weak hydrochloric acid in order to see their skeletons under a binocular microscope. A selection of 17 thin-sections of fossil sponges and five of coral samples were prepared for standard transmitted light petrography, CL, and carbonate staining with alizarin red S and potassium ferricyanide (following Dickson 1965). Fragments of sponges and corals were examined under SEM using a Jeol JSM-T6400 at the Universidad del País Vasco. Transmitted light microscope observations were made with a binocular Leica MZ 16 equipped with a DFC 320 digital camera, and with a Nikon Labophot T2-Pol petrographic microscope coupled to a Coolpix 4500 digital camera. All CL work employed a Technosyn Cold Cathode Luminescence system, model 8200 Mk II, mounted on an Olympus triocular research microscope with a maximum magnification capability of 400× using universal-stage objectives. Standard operating conditions included an accelerating potential of 12 kV and a 0.5–0.6 mA beam current with a focused beam diameter of approximately 5 mm. Three samples were analyzed by X-ray diffraction (XRD) using a Philips PW-1710 diffractometer (University of the Basque Country) with Cu-ka radiation, an automatic divergence slit, and a diffracted-beam graphite monochromator.
Fossil assemblages
Ardanatz and Aranguren
In these sections silty marls predominate. The micropaleontological content is dominated by foraminifera with scarce ostracods and charophyte gyrogonites (Ruiz de Gaona 1947; Ruiz de Gaona and Colom 1950). The foraminiferal content is high, but specimens are moderately preserved in most of the samples, usually filled with pyrite (Table 1). Benthic foraminifera strongly dominate the assemblages, the P/B ratio being 15 and 5 % in AD2.1 and AD3.1, respectively. As for the benthic foraminiferal test composition, AD2.1 and AD3.1 are dominated by hyaline taxa (77 and 63 %, respectively), agglutinated taxa are common (15 and 27 %), and porcelaneous taxa are less common (8 and 10 %). Benthic foraminiferal assemblages are moderately diverse (average 45–50 species per sample in AD2.1 and AD3.1, respectively), with similar proportions of epifaunal and infaunal taxa. The most common hyaline taxa are cibicidids, particularly in AD3.1, including Cibicides tenuimago in AD2.1, and Cibicides lobatulus and Cibicides refulgens in AD3.1. Eponides species are common in AD3.1, while Cibicidoides, Oridorsalis, and Gyroidinoides species are more abundant in AD2.1. Nodosariids, Anomalinoides, Siphonina, and Bulimina species are common and largely restricted to AD2.1. Nodosariids, Anomalinoides, Siphonina, Bulimina and Marginulina species are common and largely restricted to AD2.1. The most common agglutinated taxa in AD3.1 are calcareous-cemented taxa, including Gaudryina abuillonensis (15 %) and Spiroplectammina alansis (6 %), the latter also being common in AD2.1 (5 %). Trochamminids (6 %) and Ammobaculites species (4 %) are common in AD2.1, but G. abuillonensis is very scarce (2 %).
Macropaleontological associations include abundant sponges, corals, encrusting and erected bryozoans, Terebratulina-like brachiopods, tube-dwelling polychaetes (Rotularia spirulaea and others), mollusks (mainly gastropods: Cerithidae, Turritellidae, Volutidae, etc.; bivalves: Chama, Spondylus, Pectinidae, oysters, etc.; and scaphopods: Dentalium) and echinoderms (crinoid stalk fragments, plates—ossicles—of asteroids, and spines and test fragments of Cidaroidea). The sandstone beds contain abundant larger foraminifera (nummulitids), fossil plant debris, and Ophiomorpha and Thalassinoides-like burrows and other ichnofossils. The marine invertebrate assemblage is characterized by a great abundance, a small size of the specimens, and a low species diversity. Among fossil vertebrates there are shark teeth, turtle plates as well as sirenian vertebrae and ribs (Astibia et al. 1999, 2005, 2006; de Buffrénil et al. 2008).
Sponges are one of the most conspicuous faunal elements of the fossil assemblage. Both hexactinellids and lithistids occur, but the former predominate. The fossil sponges identified in Ardanatz and Aranguren include the species Laocoetis samueli (d’Archiac, 1850), Guettardiscyphia thiolati (d’Archiac, 1846) and/or Pleuroguettardia iberica Pisera and Busquets, 2002, cf. Rhizocheton robustus Pisera and Busquets, 2002, and Lithistida sp. indet. A. The list of corals includes the following species: Stylocoenia taurinensis (Michelin, 1842), Astrocoenia octopartita (Oppenheim, 1901), Ceratotrochus bodellei Barta-Calmus, 1973, Placosmiliopsis bilobatus (d’Achiardi, 1868), and Desmophyllum castellolense Álvarez-Pérez, 1992 (for details see below).
Arraitza
The foraminiferal content is characterized by the abundance and good preservation of benthic foraminifera, some of them filled with pyrite (Table 1). Planktic foraminifera are scarce (P/B ratio 28 %) and show a low diversity, strongly dominated by chiloguembelinid taxa. As for the benthic foraminiferal test composition, hyaline taxa dominate the assemblages (80 %), agglutinated taxa are common (17 %), and porcelaneous taxa are rare (3 %). The benthic foraminiferal assemblages are diverse (average 90 species per sample), with similar proportions of infaunal and epifaunal taxa. Cibicidoides, Eponides, cibicidids, elongate uniserial taxa (nodosariids and stilostomellids), and buliminids are the most common hyaline taxa, and include Cibicidoides ungerianus, Eponides ouachitaensis, and Cibicides lobatulus. The agglutinated taxa are dominated by trochamminids (12 %), characteristic being the presence (3 %) of tubular forms (Bathysiphon), exclusive to Arraitza.
The macrofossil assemblages from Arraitza are less diverse than in Ardanatz and Aranguren. They include sponges, Terebratulina-like brachiopods, oysters, and small bivalves (spondylids, Dimya pamplonensis, etc.), Lichenoporidae and other bryozoans, spines and test fragments of echinoids (Cidaridae and others), ossicles of asteroids, and shark teeth. The micropaleontological content is characterized by the abundance and dominance of foraminifera and common ostracods.
The sponges are: Laocoetis samueli (d’Archiac, 1850), cf. Rhizocheton robustus Pisera and Busquets, 2002, cf. Phlyctia expansa Pomel, 1872, Lithistida sp. indet. A., and Lithistida sp. indet. B. No corals have been found in Arraitza so far.
Taphonomic features of fossil sponges and corals
Ardanatz and Aranguren
Fossil sponges are especially abundant in the Ardanatz AD3 outcrop. In this outcrop sponges occur in groups within the silty marls and associated with very abundant remains of erect bryozoans. The sponges display intense fragmentation, but they often seem to correspond to complete or nearly complete specimens, without evidence of abrasion, as do the delicate bryozoan remains (Fig. 3a). They probably correspond to demic organisms and autochthonous or parautochthonous elements (sensu Fernández López 1990, 1991) deposited during periods of reduced sedimentation in a low-energy environment.

Laocoetis samueli (d’Archiac, 1850) from the Middle Eocene (Bartonian) levels of the Arraitza (ARR1) and Ardanatz (AD3) sections (Navarre). a Field photograph of a specimen from level ARR1.2 showing intense fossil-diagenetic fragmentation. b Longitudinal thin-section of the wall of specimen AD3.1.6 showing regular organization of the choanosomal opaline skeleton completely dissolved and later calcified. The spicules and axial canals (a) are filled with spatic calcite (Cc) with the same orientations. Between spicules, pelletoid microsparite (fine granular spar) (m). Crossed nicols. Width of photograph 2.4 mm. c, d Microphotographs of specimen ARR1.1.6 partially replaced by silica of a fibrous variety (quartzine-lutecite) (Qz) and sparry calcite (bottom part) (Cc), with parallel (c) and crossed nicols (d). Width of photographs 2.4 mm. e–h Lithistida sp. indet. B from the Arraitza (ARR1) section. e, f Specimen ARR1.2.10, completely replaced by a big celestine crystal (Ce) and small silica spherulites (Qz). A miliolid (mi) is preserved but also replaced by celestine. Microphotograph with parallel (e) and crossed nicols (f). Note that the quartzine-lutecite spherulites are later than the celestine crystal. Width of photographs 2.4 mm. g, h Specimen ARR1.2.8, scanning electronic microscope (SEM) images of the wall (tuberculated? tetraclones) treated with dilute HCl. Big celestine crystals (Ce), empty spaces corresponding to calcite (Cc) dissolved by the acid (g) and complex framboidal pyrites (p) associated with organic matter alteration during bacterial sulfate reduction are evident (h)
The original skeleton of Guettardiscyphia/Pleuroguettardia, and Laocoetis appears partially or totally replaced by calcite. Darker micrite/microsparite carbonate matrix fills meshes within the skeleton (Figs. 3b–d, 7l). Associations of spherical aggregates of unaltered framboidal pyrites (FeS2) can be observed (Fig. 3h). X-ray diffraction analysis of AD3.1.3 Laocoetis specimens confirms the presence of calcite and quartz.
Fossil corals occur in the Ardanatz and Aranguren sections and, with the exception of one species (Desmophyllum castellolense) described below, they appear mainly associated with marls with high terrigenous content (silty and sandy marls) or with sandstone beds with other bioclasts, such as pectinid fragments and larger foraminifera (nummulitids). The latter suggests that the corals have been reworked from shallower and more proximal environments (Fig. 4a).

Fossil corals from the Middle Eocene (Bartonian) locality of Ardanatz (Navarre). a Field photograph of a specimen of Placosmiliopsis bilobatus (d’Achiardi, 1868) from the AD3·section (level AD3.2). Scale bar 20 mm. b Transverse section of specimen AD2.1.1 of Placosmiliopsis bilobatus with borings (b). Scale bar 2 mm. c, d Placosmiliopsis bilobatus (d’Achiardi, 1868) (AD.16). Thin-section microphotograph with crossed nicols (c). Septa were initially aragonite, but are now completely neomorphized by big calcite crystals (Cc). The interseptal spaces (is) are filled with detritic sediments (quartz, mica, feldspar), micritic pellets, and opaque minerals. Under cathodoluminescence (d), the septa present a low-dull luminescence, unlike the interseptal spaces which display a characteristic intense red luminescence. Width of photograph 2.4 mm
In the fossil corals of the Ardanatz section, the septa are composed of large neomorphic sparry calcite crystals. The interseptal spaces are occupied by microsparite and, to a lesser extent, by iron oxides, quartz with irregular edges and tests of foraminifera (Rotalia, Operculina and miliolids), together with calcareous algae, smaller corals, bryozoans, echinoid spine fragments, etc. In the periphery, the crystals are arranged as equant sparry calcite. Pyrite is scarce. Borings now highlighted by sparry calcite can be observed (Fig. 4b).
Under cathodoluminescence, the septa of the corals are not luminescent or have a poor, “dull” luminescence, without evidence of zoning, while the interseptal matrix is red luminescent. It seems that the neomorphism of the septal sparry calcite occurred in a reducing environment, favoring the entry of iron (Fe2+) and manganese (Mn2+). The effect of both ions being present is to produce dull or moderately luminescing calcite depending on the exact proportions of activator (Mn2+) to quencher (Fe2+), respectively (Fig. 4d).
Arraitza
Similar to the Ardanatz AD3 outcrop, in the Arraitza section the sponges occur in groups within the silty marls. In Laocoetis the original opaline skeleton is replaced by calcite and/or shows neomorphism in the form of fibrous quartzine-lutecine spherulites. The meshes of the skeleton are infilled with microsparite (Fig. 3c–d). Unaltered framboidal pyrite aggregates are also included.
In the Lithistida sp. indet. B stem fragments, the skeleton is poorly preserved as a result of a widespread replacement with large crystals of celestite (SrSO4) and also mineralization by numerous buttons of neomorphic fibrous silica (quartzine-lutecine), together with aggregates of unaltered framboidal pyrites (Fig. 3e–h).
X-ray diffraction analysis of the ARR1.1.4 Laocoetis specimens shows the presence of calcite and quartz. Analysis of the ARR1.2.4 Lithistid sp. indet. B specimen clearly shows that, in addition to calcite and quartz, celestite occurs (Fig. 5). The petrographic evidence indicates that replacement of celestite occurs relatively early on during diagenesis in a dysoxic environment, as was detected by Boyce et al. (1990) and Ábalos and Elorza (2011) in other sections of the Basque Cantabrian basin. Large crystals of celestite include skeleton and small bioclastic sediment (miliolids) and debris trapped inside the meshes (Fig. 3e–f). Celestite may form as a result of a rapid decomposition of organic matter in a dysoxic microenvironment that produces hydrogen sulfide. The latter can be oxidized to sulfate by subsequent bacterial sulfoxidation, and this sulfate produces celestite with the mobile strontium, the latter coming either from the aragonite-to-calcite neomorphism (Taberner et al. 2002), or from other processes, such as decomposition of inherited sheet silicate mineral of continental origin with a decrease-acidification of the pH (Baker and Bloomer 1988).

Diffractogram of the siliceous sponge Lithistida sp. indet. B from the Bartonian of Arraitza (specimen ARR1.2.4) showing a calcite (Cc), quartz (Qz) and celestite (Ce) composition. The horizontal axis at the bottom represents the Bragg scattering angle in degrees 2Θ in the figure. The vertical axis represents intensity in counts per second
Taxonomic paleontology
-
Phylum Porifera Grant, 1836
-
Class Hexactinellida Schmidt, 1870
-
Subclass Hexasterophora Schulze, 1887
-
Order Hexactinosa Schrammen, 1903
-
Family Craticulariidae Rauff, 1893
-
Genus Laocoetis Pomel, 1872
-
Type species: Laocoetis crassipes Pomel, 1872
-
Laocoetis samueli (d’Archiac, 1850) (Figs. 3a–d, 6a–d)
Fig. 6 
Laocoetis samueli (d’Archiac, 1850) from the Middle Eocene (Bartonian) levels of the Arraitza (ARR1) and Ardanatz(AD3) sections (Navarre). a, b, Specimen ARR1.2.7, outer (a) and inner (encrusted with a colony of bryozoans) (b) side views. c, Specimen AD3.1.5, outer surface view. d, Specimen ARR1.2.6, treated with diluted HCl, showing a regular craticulariid pattern of canal openings and the dictyonal skeleton. Dictyonal skeleton on the surface with hemispherical and tuberculated nodes (SEM). cf. Rhizocheton robustus from the Middle Eocene (Bartonian) levels of the Arraitza (ARR1) and Ardanatz (AD3) sections (Navarre). e, Specimen AD3.1.12, stem fragment. f, g Specimen ARR1.1.1, stem fragment (f). Strongly modified dictyonal skeleton, reflected light (g) (photograph is 7 mm wide). Scale bars of figures a, b, c, e and f represent 20 mm
-
1850 Scyphia samueli—d’Archiac: 413, pl. 9, figs. 11–12

Material.
Numerous fragments from the Ardanatz (especially the lower part of AD3; also present, but scarce, in AD2.1) and Arraitza (ARR1) sections; a small sample comes from the Aranguren section (level AG1.1).
Description.
The fragments belong to cup- or vase-shaped (cyathiform) sponges, with a widely folded wall that is 2.2–4.7 mm thick displaying oval canal openings on the outer and inner surfaces, arranged in a regular rectangular pattern. In some fragments the maximum diameter of the outer surface canal openings is 0.5–0.8 mm. Skeletal bridges between openings are 0.9–1.1 mm wide horizontally, and 0.9–1.0 mm wide vertically. On the inner surface the canal openings are 0.5–1.1 mm in diameter, and are separated by skeletal bands 0.9–1 mm wide horizontally and 0.8–1 mm wide vertically. In other fragments, the maximum diameter of the outer and inner canal openings is wider (1.21–1.44 mm and 0.9–1.33 mm). The openings on the outer surface are more oval than on the inner surface. The choanosomal dictyonal skeleton has more or less quadrangular meshes, which are 0.20 by 0.25 mm wide.
Remarks.
Very characteristic sponges that are cyathiform, with a widely folded wall and displaying oval canal openings on the outer and inner surfaces, arranged in regular rectangular pattern, belong to the genus Laocoetis Pomel, 1872, most probably Laocoetis samueli (d’Archiac, 1850). The genus Laocoetis (= Craticularia Zittel, 1877) was defined by Pomel (1872) for fossil sponges from the Miocene of Algeria. L. samueli (under the name “Scyphia Samueli”) was first described for fossil sponges from the Eocene of the Biarritz area (southwestern Aquitaine Basin, Basque Country) by d’Archiac (1850). This species is probably also present in the Middle Eocene (Early Lutetian) of Val di Chiampo (Italy), where Matteucci and Russo (2005) described and figured a specimen (Laocoetis sp.) with oval canal openings on both surfaces. A similar species, L. patula Pomel, 1872, which is abundant in the Eocene (Bartonian-Priabonian, SBZ 18; Costa et al. 2012) of the Vic Marls Formation (eastern margin of the Ebro Basin, Catalonia), was described by Pisera and Busquets (2002). L. patula has rectangular canal openings on the outer surface, which differ from the rounded canal openings of L. samueli.
-
Order Lychniscosa Schrammen, 1903
-
Family unknown
-
Genus Rhizocheton Lachasse, 1943
-
Type species: Xylospongia tarraconensis Hérenger, 1942
-
cf. Rhizocheton robustus Pisera and Busquets, 2002 (Fig. 6e–g)
Material.
Numerous stem fragments from the lower part of the Arraitza (level ARR1.1) and Ardanatz AD3 (level AD3.1) sections.
Description and remarks.
Lychniscosan sponges are also numerous, represented mostly by more or less cylindrical? fragments bearing irregular longitudinal furrows on the surface of the choanosomal dictyonal skeleton, which is strongly modified and irregular. No lychnisc nodes have been observed in our material, but both the morphology and character of the skeleton, which is identical to that in stem fragments of Rhizocheton robustus as described by Pisera and Busquets (2002), suggest that we are most probably dealing with the same species.
Other Hexactinellida
-
Guettardiscyphia thiolati (d’Archiac, 1846) and/or Pleuroguettardia iberica Pisera and Busquets, 2002 (Fig. 7a–l)
Fig. 7 
Guettardiscyphia thiolati (d’Archiac, 1846) and/or Pleuroguettardia iberica Pisera and Busquets, 2002, from the Middle Eocene (Bartonian) levels of the Ardanatz (AD3) section (Navarre). a, b Specimen AD3.1.2, lower part of the sponge with folded wall, in lateral view (a1) and cross section (b). c-e Specimen AD3.1.7, lower part, encrusted with a tube-dwelling polychaete, details of the canal openings and parietal oscula, in lateral view (c), enlarged view of the surface (d) and regular dictyonal choanosomal skeleton in cross section (e). Width of photographs d and e 4 mm and 8 mm, respectively. f, g Specimen AD3.1.4, folded stellate part, in lateral view (f) and cross section (g). h, i Specimen AD3.1.10, outer surface (h) and inner surface views of a wing (i). j, k Specimen AD3.1.11, lower part of the sponge with folded wall, in lateral view (j) and cross section (k). l AD3.1.8, thin transverse section of the wall showing calcitic replacement of spicules. Width of photograph 2.4 mm. Scale bars represent 20 mm

Material.
Numerous fragments from the lower part of the Ardanatz AD3 section (level AD3.1).
Description.
Sponges that are tubular (in the lower part) to folded in a star-like pattern in transverse section in the upper part. The stellate pattern originates from the plication of the wall of the tubular part. The number of folds or wings in the studied fossils is 3, 4 or 6, and they are 6.9–8.8 mm thick. The thickness of the wall ranges between 1.5 and 5.2 mm, at the base or at the ends of the wings, respectively. Along the edge of the folds or wings parietal oscula, 2.1–4.6 mm in diameter, may occur. These are separated by skeletal bands exceeding 18 mm in length and being most commonly only 2.5–5.5 mm wide. Oscula were not found in the lateral parts of the wings. In the more tubular parts, the canal openings are generally irregularly arranged on the outer surface, but other parts exhibit a more regular organization of the canal openings. Their maximum diameter is between 0.12 and 0.56 mm, and they are 0.15–0.77 mm apart. The openings on the inner surface are more regularly distributed. Their diameter, which is less variable, ranges between 0.30 and 0.68 mm, and they are 0.14–0.56 mm apart. Dictyonal skeleton can be observed only in some of the best-preserved parts, and is regular.
Remarks.
These very common hexactinellids are representatives of the homeomorphic genera/species Guettardiscyphia (most probably Guettardiscyphia thiolati (d’Archiac, 1846)) and Pleuroguettardia (most probably Pleuroguettardia iberica Pisera and Busquets, 2002). Both species are very similar in general morphology but belong to different families that differ in their canalization pattern, i.e. Cribrospongiidae (cribrospongiid or irregular pattern) and Craticulariidae (quadrangular, craticulariid pattern). This pattern is often disturbed in both genera, especially near parietal gaps, so assignment to either of the genera may be difficult in the case of poorly preserved and/or fragmentary material (Pisera and Busquets 2002), and assignment is rather difficult without detailed studies. It seems that in the investigated material, both genera/species are present.
We could not locate the collection of sponges described by d’Archiac (1846) from the Eocene of Rocher de Goulet, near Biarritz. In the original description of “Guettardia thiolati “, d’Archiac (1846) mentioned, however, that “les cellules [= canal openings] ne paraissent pas d’ailleurs affecter une disposition symétrique régulière”, thus suggesting that this species belongs to Guettardiscyphia, despite the fact that the specimens illustrated by d’Archiac (1846, pl. 5, fig. 15, pl. 8, figs. 5–7) display only their stellate form, but not the arrangement and shape of the canal openings. In a later work d’Archiac described and figured the new species Scyphia quinquelobata (d’Archiac 1850, pl. 9, fig. 13) from the Eocene of Biarritz. His illustration does not show the distribution of the canal openings. In our opinion, this may be a fragment of the basal portion of “Guettardia thiolati”, but with folds less developed than in the specimens figured by d’Archiac (1846).
Pleuroguettardia iberica is known from the Eocene (Bartonian-Priabonian) of the Vic Marls Formation (Pisera and Busquets 2002) and from the Middle Eocene (Early Lutetian) of Val di Chiampo, where Matteucci and Russo (2005) also assigned some specimens to Pleuroguettardia aff. iberica and Pleuroguettardia sp. In both these localities Guettardiscyphia thiolati is also present.
-
Class Demospongia Sollas, 1875
-
“Lithistida”
Preliminary remarks.
Lithistida were originally described as, and long considered, a formal taxonomic unit of the rank of order. Pisera and Levi (2002) proposed formally that this is a polyphyletic group, with desmae that developed independently in various groups of demosponges of different ages. Consequently, the order Lithistida is now abandoned, and the term lithistid sponges should only be used in an informal way.
-
Suborder Rhizomorina Zittel, 1878
-
Genus Phlyctia Pomel, 1872
-
Type species: Phlyctia expansa Pomel, 1872
-
cf. Phlyctia expansa Pomel, 1872 (Fig. 8a–e)
Fig. 8 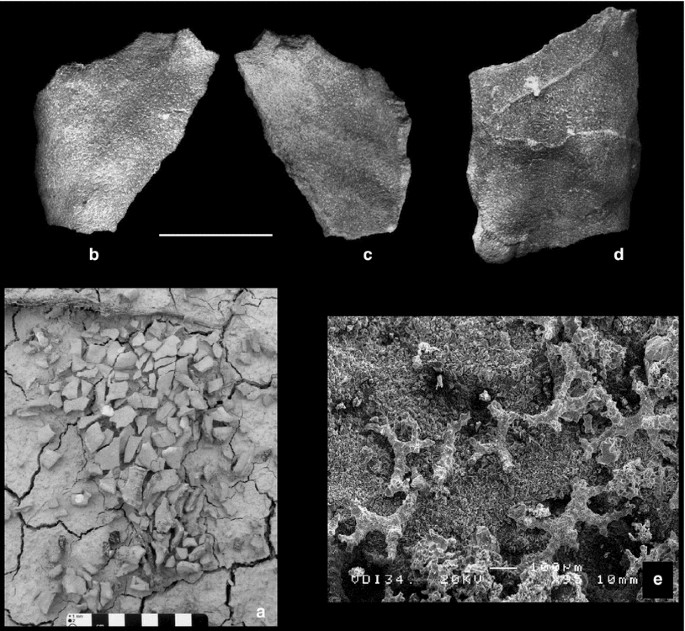
cf. Phlyctia expansa Pomel, 1872, from the Middle Eocene (Bartonian) level of the Arraitza (ARR1) section. a Field photograph of a specimen from level ARR1.2 showing diagenetic fragmentation. b, c Specimen ARR1.2.11, in outer (b) and inner (c) side views. d Specimen ARR1.2.12, in outer view. e Specimen ARR1.2.13, treated with diluted HCl showing the choanosomal skeleton composed of spinose rhizoclones (SEM picture). Scale bars 20 mm

-
1872 Phlyctia expansa—Pomel: 236, pl. 12, fig. 4, pl. 15, fig. 4
-
1872 Phlyctia cupulata—Pomel: 327, pl. 15, fig. 5
-
1924 Phlyctia expansa Pomel—Moret: 17, fig. 4, pl. 1, figs. 8–9, pl. 2, fig. 1
Material.
Numerous fragments from the upper part of the Arraitza section (level ARR1.2).
Remarks.
Most common among the lithistid sponges are undulating and leafy specimens that have rhizoclone desmae. These most probably belong to the genus Phlyctia Pomel, 1872. The studied specimens are similar to those figured by Pisera and Busquets (2002) from the Eocene of Catalonia, as well as to Pomel’s species (Pomel 1872; Moret 1925). It is possible that this species is also present in the Eocene of Italy in Val di Chiampo (Italy) (Matteucci and Russo 2005).
Other lithistids
-
Lithistida sp. indet. A (Fig. 9a–g)
Fig. 9 
Lithistid sp. indet. A from the Middle Eocene (Bartonian) levels of the Arraitza (ARR1) and Badoztain (BA1) sections. a, b Specimen ARR1.2.14, in upper (a) and lower (b) face views. c, d Specimen ARR1.2.15, in upper (c) and lateral (d) views. e, f Specimen BA1.1.1, in upper (e) and lower (f) face views. g Specimen BA1.2.2, choanosomal skeleton showing tuberculated ?tetraclones (SEM picture). Lithistida sp. indet. B from the Middle Eocene (Bartonian) levels of the Arraitza (ARR1) and Ardanatz (AD3) sections (Navarre). h, i Specimen ARR1.2.3, fragment of the cup-like part in outer (h) and inner (i) side views. The external part of the wall is covered with a characteristic network of channels in the choanosomal skeleton. j Specimen ARR1.2.2, outer surface view showing oscula and an encrusting lichenoporid bryozoan. Lithistida sp. indet. B (?), stem fragments. k Specimen ARR1.2.4, in lateral view. l Specimen ARR1.2.5, in lateral view. m Specimen ARR1.2.8, tuberculated ?tetraclones (SEM picture). Scale bars of a–f 10 mm; scale bars of h–l 20 mm

Material.
Six specimens from the upper part of the Arraitza section and one specimen from the lower part (levels ARR1.2 and ARR1.1), three specimens from the lower part of the Ardanatz AD3 section (level AD3.1) and two specimens from the upper part of the Badoztain section (BA1.2).
Description and remarks.
Small, hemispherical or flattened globular sponges. Their surface is finely striated with a series of deep grooves or channels with small and more or less radially arranged pores. Their desmae are regularly tuberculated ?tetraclones (with terminal articulation). This sponge is common in the two study areas. From its morphology and size it is comparable to the sponge “Tragos mamillatus” described by d’Archiac (1850, pp. 413, pl. 9) in the Eocene of Biarritz. More precise determinations need further detailed studies of spiculation.
Material.
Four cup-shaped fragments and numerous fragmentary stems from the upper part of the Arraitza section (level ARR1.2).
Description and remarks.
A cup-shaped sponge with strong protuberances on the outer surface and endowed with a stem, its desmas are tuberculated ?tetraclones. Morphologically these sponges look like the specimens figured by Pisera and Busquets (2002, figs. 8–11) from the Eocene of Catalonia, which were provisionally classified as tetracladine lithistid sp. A. The sponges of Arraitza are unfortunately too poorly preserved to be accurately determined; tuberculated tetraclones occur in many lithistid genera, but if the morphology is not well known, a precise determination is not possible. More detailed studies are required.
-
Phylum Coelenterata Frey and Leuckart, 1847
-
Class Anthozoa Ehrenberg, 1834
-
Subclass Zoantharia de Blainville, 1830
-
Order Scleractinia Bourne, 1900
-
Suborder Astrocoeniina Vaughan and Wells, 1943
-
Family Astrocoeniidae Koby, 1890
-
Subfamily Astrocoeniinae Koby, 1890
-
Genus Stylocoenia Milne-Edwards and Haime, 1849
-
Type species: Astrea emarciata Lamarck, 1816
-
Stylocoenia taurinensis (Michelin, 1842) (Fig. 10a–c)
Fig. 10 
Fossil corals from the Middle Eocene (Bartonian) locality of Ardanatz (Navarre). Stylocoenia taurinensis (Michelin, 1842). Specimen AD.10, views of a fragment of a cerioid colony (a, b) and detail of the colony (c). Astrocoenia octopartita (Oppenheim, 1901). Specimen AD.11, view of a fragment of a cerioid colony (d) and detail of the colony (e). Scale bars of a, b, d 20 mm; scale bars of c–e 5 mm

-
1842 Astraea taurinensis—Michelin: 62, pl. 13, fig. 3
-
1847 Madrepora taurinensis (Michelin)—Reuss: 27, pl. 5, fig. 2
-
1856 Astrea bistellata—Catullo: 66, pl. 7, fig. 4
-
1866 Astrocoenia laminosa—d’Achiardi: 45, pl. 4, fig. 3
-
1902 Stylocoenia minuscula—Osasco: 114, pl. 9, fig. 3
-
1988 Stylophoropsis renzii—Reig: 7, pl.1, fig. 2
-
1990 Hydnoseriatopora viai—Reig: 33, pl.7, figs. 2–4
-
1990 Colliastrea guillermoi—Reig: 46–47, pl. 10, figs. 1–3
Stratigraphic distribution.
Eocene to Middle Oligocene.
Material.
One well-preserved specimen from Ardanatz (Eguesibar) (section and level unknown).
Description.
Cerioid colony, massive or ramose. The calices are circular, moderately elliptical or polygonal. Adjacent calices are directly bonded along the wall. The septa are arranged in two cycles, with hexameral radial symmetry. S1 are joined with the columella. S2 are shorter and slender. The margins of the septa are dentate. The columella is styliform and prominent. The columns of the septothecal wall are conical with an oval base and vertical grooves. They appear at the junction of three or more calices, rarely in a single wall. The endotheca is formed by numerous thin tabulae. Coenosteum granulose. Budding extratentacular. Microstructure not preserved. Calyx diameter: 0.9–3 mm; wall columns: base 0.2–1.1 mm; height 0.1–0.6 mm.
Remarks.
Stylocoenia taurinensis has very variable growth forms. The three new species described by Reig (1988, 1990) are simply variations of this species, and their parameters are included within the limits indicated by Zlatarski (1963). The wall columns are only visible in well-preserved areas of some polyps. They are scarce and difficult to see. This has led to serious doubts being cast on the validity of the genus (Solé Sabarís 1942). These wall columns have been described and figured by Zlatarski (1963) and Álvarez-Pérez (1993).
Stylocoenia taurinensis is present in the Paleogene coral associations of mixed platforms in the Balearic Islands (Ramos-Guerrero et al. 1989-1990) and in the coral associations of the South Pyrenean Eocene Foreland Basin (Álvarez et al. 1994; Álvarez-Pérez 2009).
This species can be distinguished (a) from Stylocoenia sanctaorosiae Altuna and Álvarez-Pérez, 2003 from the Bartonian of the Jaca Basin (South Central Pyrenean zone, Huesca) by its growth pattern, the latter being found in spherical or slightly flattened, non-branching colonies with septa arranged in octameral as opposed to hexameral symmetry (Altuna et al. 2003); (b) from Stylocoenia aurelii Álvarez-Pérez, 1997 from the La Tossa Formation (Priabonian of Igualada, eastern margin of the Ebro Basin, Catalonia), by its bigger calices, the smaller distance between calicular centers and by its less robust wall pillars (Álvarez-Pérez 1997); (c) from Stylocoenia macrostyla Reuss, 1870 by its spherical colonies, its robust wall pillars and septa arranged in octameral symmetry; (d) from Stylocoenia sanmigueli Solé-Sabarís, 1942 by its cupulate colonies and its robust wall pillars; (e) from Stylocoenia emarciata Lamarck, 1816 by its spherical colonies and its robust wall pillars; and (e) from Stylocoenia neutra Barta-Calmus, 1973 by its much lighter corallum and much shorter septa.
-
Subfamily Pinacophyllinae Vaughan and Wells, 1943
-
Genus Astrocoenia Milne-Edwards and Haime, 1848
-
Type species: Astrea numisma Defrance, 1826
-
1901 Goniaraea octopartita—Oppenheim: 201, pl. 16, fig. 2
-
1912 Dictyaraea octopartita—Oppenheim: 97, 134
-
1988 Madracis calzadai—Reig: 10, pl. 1, fig. 4, pl. 2, fig. 2
-
1990 Madracis octopartita—Reig: 40–42, pl. 7, figs. 7–9
-
1990 Pseudodictyaraea rosi—Reig: 21–22, pl. 5, figs. 5–6
-
1990 Septastrea catalaunica—Reig: 44–45, pl. 9, figs. 3–4
Stratigraphic distribution.
Bartonian and Priabonian.
Material.
One well-preserved specimen from Ardanatz (Eguesibar) (section and level unknown).
Description.
Cerioid colony, massive or ramose. Circular calyces are regularly arranged and equal in size. In thin branches the calices are polygonal. The septa are arranged in two cycles with octameral radial symmetry. S1 are thick, and the distal margin is dentate with 4–5 stout teeth. They are joined to the columella. S2 are shorter and at depth join the columella. The septa are regularly perforate, and their lateral faces are slightly granulose. The columella is styliform, occasionally laminar, and forms, with two opposite S1, a plane of bilateral symmetry. Wall septothecal. The endotheca consists of numerous tabulae that enclose the bottom of the calyx. Budding extratentacular. Coenosteum scarce and granulose. Microstructure not preserved. Calyx diameter: 1.4–3.4 mm.
Remarks.
Reig (1990) created the new genus Pseudodictyaraea versus Dictyaraea, but the description of this new genus coincides with that of the genus Astrocoenia, and the description of P. rosi coincides with that of Astrocoenia octopartita. With regard to the genus Madracis, the S1 do not reach the columella, which is highly developed, and S2 are reduced to spines. Analyzing our specimen in detail, it does not agree with these descriptions, but matches the description of Astrocoenia octopartita. With regard to Septastrea catalaunica, it is simply a variety of Astrocoenia octopartita, which was previously known from the coral associations of the South Pyrenean Eocene Foreland Basin (Álvarez et al. 1994; Álvarez-Pérez 2009).
-
Suborder Caryophylliina Vaughan and Wells, 1943
-
Superfamily Caryophyllicae Gray, 1847
-
Family Caryophyllidae Gray, 1847
-
Subfamily Nicaeotrochinae Barta-Calmus, 1973
-
Genus Ceratotrochus Milne-Edwards and Haime, 1848
-
Type species: Ceratotrochus multiserialis Milne-Edwards and Haime, 1848
-
Ceratotrochus bodellei Barta-Calmus, 1973 (Fig. 11a–c)
Fig. 11 
Fossil corals from the Middle Eocene (Bartonian) locality of Ardanatz (Navarre). Ceratotrochus bodellei Barta-Calmus, 1973. a, b Specimen AD.02, corallum in lateral (a) and upper (b) views. c Specimen AD2.1.4, corallum. Placosmiliopsis bilobatus (d’Achiardi, 1868). d, e Specimen AD.04, in lateral (d) and upper (e) views. f, g Specimen AD.08, in lateral (f) and upper (g) views. h, i Desmophyllum castellolense Alvarez-Pérez, 1992. Specimen AD1.1.1, corallum in lateral (h) and upper (i) views. j Bryozoan borings on coral specimen AD1.2.3. k detail of the bryozoans borings of specimen j. Scale bars of a–c, h–k 5 mm; scale bars of d–g 20 mm
-
1973 Ceratotrochus bodellei—Barta-Calmus: 432, pl. 41, figs. 14–18

Stratigraphic distribution.
Bartonian and Priabonian.
Material.
Two specimens from the Ardanatz AD1 section (level AD1.3?), 10 specimens from the Ardanatz AD2 section (level AD2.1), 1 specimen from Ardanatz (section and level unknown), 1 specimen from the Aranguren AG1 section (level AG1.3) and 3 specimens from the lower part of Urbikain (Izagaondoa) section, about 12 km southeast of the latter locality.
Description.
Solitary, trochoid, fixed or free. Curved at the base. Calyx elliptical. The axis of curvature is parallel to the short axis of the calyx. Flattened in the direction of the long axis of the calyx. The septa are arranged in four cycles with hexameral radial symmetry. S1 and S2 with paliform teeth. Columella papillose. The outer part of the curve has five to seven triangular and flat teeth. Wall septothecal. Costae slightly marked, coated with a thin epitheca.
Remarks.
Ceratotrochus includes deep-water species and sublittoral species. Ceratotrochus bodellei is present in the coral associations of the South Pyrenean Eocene Foreland Basin of Igualada, Manresa and Jaca (Álvarez-Pérez 2009).
-
Subfamily Parasmiliinae Vaughan and Wells, 1943
-
Genus Placosmiliopsis Russo, 1979
-
Type species: Placosmilia bilobata d’Achiardi, 1868
-
1868 Placosmilia bilobata—d’Achiardi: 25
-
1875 Placosmilia eocenica—d’Achiardi: 9
-
1979 Placosmiliopsis bilobatus (d’Achiardi)—Russo: 61, pl. 8, figs. 1–2
Stratigraphic distribution.
Ilerdian, Bartonian and Priabonian.
Material.
Three specimens from the upper part of the AD3 section of Ardanatz and 10 specimens from the same locality but from unknown sections and levels.
Description.
Solitary coral, with a flabellate and compressed form. Attachment by a basal peduncle. Elliptical calyx. Deep fossula elongated in the direction of the long axis of the calyx. The septa are arranged in six cycles with hexameral radial symmetry. S1 and S2 are thick and rhopaloid. S3 are thinner and slightly shorter. The other septa are still thinner and shorter. Distal margins of the septa are smooth. The lateral faces of the septa are spinose. The columella is lamellar, sinuous and deep. Wall paraseptothecal at the surface and septothecal at depth. Alternating thick and thin costae run the length of the surface of the corallum.
Remarks.
P. bilobatus is present in the coral associations of the South Pyrenean Eocene Foreland Basin (Álvarez-Pérez 2009).
-
Subfamily Desmophyllinae Vaughan & Wells, 1943
-
Genus Desmophyllum Ehrenberg, 1834
-
Type species: Desmophyllum cristagalli Milne-Edwards and Haime, 1848
-
1997 Desmophyllum castellolense—Álvarez-Pérez: 214, pl. 17, figs. 1–3
Stratigraphic distribution.
Bartonian and Priabonian.
Material.
Two specimens from the Ardanatz AD1 section (level AD1.1)
Description.
Solitary coral, with a trochoid form. Usually taller than wide. Slightly curved at the base. Calyx circular or elliptical. The septa are arranged in five cycles with hexameral radial symmetry. S1 and S2 are prominent, and the lateral faces are smooth. The axial edges exhibit laminar extensions anastomosing towards the columella. They join the columella and have a rhopaloid inner margin. The lateral faces of the remainder of the septa are spinose. The columella is lamellar and deep. Endothecal dissepiments are irregularly developed. The costae of the first two cycles are more prominent than the remainder of the septa. Costae are well developed near the calyx. Wall septothecal, covered with fine granules. Growth lines paralleling the calicular margin (Álvarez-Pérez 1993, 1997). Coral height: 65–113 mm; calyx diameter: 7–42 mm.
Remarks.
Desmophyllum castellolense was first described by Álvarez-Pérez (1997) on the basis of material from the La Tossa Formation (Bartonian of Igualada, eastern margin of the Ebro Basin, Catalonia). Some authors claim that Desmophyllum has no columella, but the existence of a small columella at the bottom of the calyx is currently considered possible. D. castellolense was a species that lived in deep waters. In its juvenile stage it was attached to the substrate but in its adult stage it broke its basal attachment and lived freely.
Paleoecological remarks
The sponges and corals are commonly encrusted by cyclostome and cheilostome bryozoans, small bivalves, serpulids, and cibicidid foraminifers (Figs. 7c, j, k, 9j). Borings produced by bryozoans on corals are very abundant (Fig. 11j).
As mentioned above, in the Ardanatz AD3 and Arraitza sections sponges occur within marl and seem to correspond to complete or nearly complete specimens. In the AD3 section fossil sponges appear with very abundant remains of well-preserved erect bryozoans, probably in life position. These features suggest a low-energy environment (Figs. 3a, 8d).
Preliminary data suggest the existence of some differences among the faunal compositions of the studied sections. Laocoetis is clearly the most abundant sponge and is present in the Ardanatz (AD3, AD2), Aranguren (AG1), and Arraitza (ARR1) sections. cf. Rhizocheton robustus and Lithistida sp. indet. A occur in AD3 and ARR1. Lithistida sp. indet. A is present in Badoztain, but it has not been found in the Aranguren outcrop. Guettardiscyphia/Pleuroguettardia have been found only in the Ardanatz AD3 section, and cf. Phlyctia expansa and Lithistida sp. indet. B only in Arraitza.
Although additional taxa not described in this paper may occur in the studied sections, the fossil association of sponges of the Ardanatz Sandstone and the Ilundain Marls of the Pamplona Basin appears less diverse than those of the Bartonian-Priabonian of the Vic Marls Formation in the northeastern part of the Ebro Basin, Catalonia (6 vs. 16 genera) (Pisera and Busquets 2002) and of the Early Lutetian of Val di Chiampo (Italy) (Matteucci and Russo 2005). The Pamplona Basin sponge association is more similar to that of the Eocene of Biarritz (Aquitanian Basin, Basque Country) described by d’Archiac (1846, 1850), where Laocoetis samueli and stellate forms such as Guettardiscyphia thiolati and Pleuroguettardia iberica are the most representative sponges of the fossil assemblage.
Despite the apparent lower diversity of the Ardanatz AD3 level, the stellate and conic siliceous sponges and erect bryozoan colonies “in growth position” can be compared to the shelf deposits from the Vic Marls Formation (Busquets et al. 1994). Pisera and Busquets (2002) estimated a minimum of 100 m, and more probably 200 m or more, for the depth of the Vic Marls with sponge faunas (La Guixa and Gurp members). As pointed out by these authors, Guettardiscyphia and Pleuroguettardia are extinct and cannot be used as bathymetric indicators. The Recent species Laocoetis perion Lévi, 1986 from the Indian Ocean has been found at depths of 350–750 m. A similar depth (300–400 m) is assumed by Brimaud and Vachard (1985) for a Miocene Laocoetis species. Most hexactinellids live in deep waters on soft substrates (Tabachnik 1991). The dominance of hexactinellids and erect morphologies shows that this sponge community lived in deep waters (Charbonnier et al. 2007). Today, lithistid sponges are common in rather deep water (from 100 m down to several hundred meters), but theonillids may also occur in shallow waters (Vacelet 1988; Pisera et al. 2006). The low proportion of planktic foraminifera and the high abundance of benthic foraminifera, their composition (including common porcelaneous foraminifera) and moderate diversity suggest a shelfal depth of deposition (Murray 2006).
The paleontological and stratigraphic data indicate that of the two studied areas the Arraitza outcrop may correspond to somewhat deeper environments. The fine-grained nature of the marl, the absence of sediment gravity-flow deposits and the absence of light-dependant organisms suggest that the Arraitza deposits accumulated in a flat, aphotic seabed located well below storm-wave base. This interpretation is supported by the foraminiferal content. The scarcity of planktic foraminifera in Arraitza, which is slightly higher than in Ardanatz, and the diversity and composition of benthic foraminifera, including the presence of agglutinated tubular forms (Jones and Charnock 1985; Murray et al. 2011), are indicative of outer-platform to upper-slope environments, of approximately 200 m water depth. The peak in chiloguembelinid abundance recorded in Arraitza, usually associated with nutrient-enriched waters (Li et al. 1995; Malumián et al. 2009), and the high abundance of benthic foraminifera may be indicative of increased local productivity. Furthermore, the lower abundance of erect bryozoans, the apparent absence (?) of larger foraminifers, and the presence of pelagic shark teeth (Odontaspididae, Mitsukurinidae and Hexanchidae; C. Corral pers. comm.) support this interpretation.
With regard to corals, their abundance at the Ardanatz locality is out of phase with that of sponges: where many sponges occur, there are few, if any, corals. With the exception of Desmophyllum castellolense, which is a deep-water inhabitant, corals are more common in “dirty” marls (with silt and sand) or in sandstones with abundant bioclasts, thus indicating that corals lived in shallower areas than sponges and that they were transported from shallower environments (by removal, reworking processes). Ceratotrochus can be a deep-water inhabitant, but the species C. bodellei seems to be sublittoral. It does not co-occur with Desmophyllum castellolense (level AD1.1, Fig. 2), but, mainly, in the level AD2.1, with few sponges. In Arraitza many sponges occur, but no corals have been found to date. Consequently, corals and sponges are regarded as living in spatially and/or temporally close but different environments.
It is hence reasonable to conclude that there was a continuous facies transition from the most proximal Ardanatz delta-front facies into the most distal/marginal outer-platform to upper-slope Ilundain Marl facies. Taking everything into account, the Ardanatz–Aranguren and Arraitza fossil sites can be regarded as the proximal and distal end-members, respectively, of a continuous spectrum of approximately coeval fossil assemblages. Given this depositional setting, the small size of some macrofossils and the inferred reducing environment suggest that fully oceanic conditions were not achieved, most probably due to paleogeographic constraints in the Bartonian South Pyrenean Basin.
Concluding remarks
Fossil sponges and corals from the extensive Bartonian marly formations of the Pamplona Basin (south Pyrenean area, Navarre) have been described for the first time. Although the fossil sponges exhibit intense diagenetic fragmentation, they often correspond to complete specimens, probably to demic organisms and autochthonous or parautochthonous elements. Other significant taphonomic processes that have affected the skeletons include mineralogical replacement and neomorphism. The sponge and coral taxa are similar to those previously described from other contemporaneous geological formations of the Pyrenean realm. The Pamplona Basin assemblages are, however, less diverse—five or six species of sponges and five of corals—than those from the Eocene of Catalonia—16 species of siliceous sponges are described by Pisera and Busquets (2002), and 65 species of corals according to Álvarez-Pérez (2009)—and other European areas. At least with respect to the sponges this is not due to a lower sampling resolution, but to taphonomic biases and/or paleoecological differences. The dominance of hexactinellids, erect morphologies, as well as sedimentological and micropaleontological data show that the sponge communities lived in deep shelfal waters. The corals, mainly associated with levels with higher terrigenous content, were probably reworked from shallower and more proximal environments.
References
Abad (2001) Paleotaxodonta y Pteriomorphia del Eoceno del Margen Sur de la Depresión Catalana. Unpublished Ph. D. thesis, Universitat Autònoma de Barcelona, Barcelona, 803 p
Ábalos B, Elorza J (2011) Latest Cretaceous cone-in-cone structures and soft-sediment deformation (Basque-Cantabrian Basin, north Spain): a record of deep-marine paleoseismicity? GSA Bull 123:427–438
Alameda J, Blas I, Cabra P, Del Valle de Lersundi J, García A, Martínez A, Ramírez del Pozo J, Solé J (1993) Cartografía Geológica de Navarra, escala 1:25.000, Hoja 141-II, Pamplona. Gobierno de Navarra, Departamento de Obras Públicas, Transportes y Comunicaciones
Altuna A, Álvarez-Pérez G, Busquets P, Etayo V (2003) Five new species of Bartonian (Eocene) corals: Jaca Basin, Pyrenees, Spain. In: Hubmann B, Piller WE (eds) Fossil corals and sponges. Proceedings of 9th international symposium on fossil Cnidaria and Porifera, Österr Akad Wiss Schriftenreihe Erdwiss Wien, pp 435–453
Álvarez G, Busquets P, Taberner C, Urquiola MM (1994) Facies architecture and coral distribution in a mid Eocene reef tract, South Pyrenean Foreland Basin (NE Spain). Cour Forsch-Inst Senckenberg 172:249–260
Álvarez-Pérez G (1993) Cnidaria fóssils de la Conca d’Igualada. Unpublished Ph. D. thesis, Universitat de Barcelona, Barcelona, 270 p
Álvarez-Pérez G (1997) New Eocene coral species from Igualada (Barcelona, NE of Spain). Bol Real Soc Esp Hist Nat 91:297–304
Álvarez-Pérez G (2009) Actualització de les espècies coral.lines de l’Eocè de les conques sudpirinenques. Batalleria 14:5–10
Astibia H, Murelaga X, Payros A, Pereda X, Tosquella J (1999) Tortugas y sirenios fósiles en el Eoceno marino de Navarra y Cuenca de Jaca. Geogaceta 25:15–18
Astibia H, Payros A, Pereda Suberbiola X, Elorza J, Berreteaga A, Etxebarria N, Badiola A, Tosquella J (2005) Sedimentology and taphonomy of sirenian remains from the Middle Eocene of the Pamplona Basin (Navarre, western Pyrenees). Facies 50:463–475
Astibia H, Pereda Suberbiola X, Bardet N, Payros A, Berreteaga A, Badiola A (2006) Nuevos fósiles de sirenios en el Eoceno medio de la Cuenca de Pamplona (Navarra). Rev Espan Paleont 21:79–91
Baker PA, Bloomer SH (1988) The origin of celestite in deep-sea carbonate sediments. Geochim Cosmochim Acta 52:335–339
Barnolas A, Payros A, Samso JM, Serra-Kiel J, Tosquella J (2004) La Cuenca surpirenaica desde el Ilerdiense medio al Priaboniense. In: Vera JA (ed) Geología de España. Instituto Geológico y Minero de España, Sociedad Geológica de España, Madrid, pp 313–320
Barta-Calmus S (1973) Révisions de collections de madréporaires provenant du nummulitique du Sud-Est de la France, de l’Italie et de la Yougoslavie septentrionales. Thèse de doctorat d’Etat, Université de Paris, vol 6, pp 1–695
Boussac J (1911) Études stratigraphiques et paléontologiques sur le nummulitique de Biarritz. Annales Hébert, Annls Stratigr Paléont Lab Géol Fac Sci Univ Paris vol, 5, pp 1–95
Boyce AJ, Fallick AE, Hamilton PJ, Elorza J (1990) Diagenesis of celestite in quartz geodes from the Basque-Cantabric basin, Northern Spain: evidence from sulfur and strontium isotopes. Chem Geol 84:354–356
Brimaud C, Vachard D (1985) Indications paléoécologiques fournies par les spongiaires du Miocène supérieur d’Espagne. Bull Mus Natl Hist Nat C7:3–11
Busquets P, Álvarez G, Solé de Porta N, Urquiola MM (1994) Low sedimentation rate in aphotic shelves with Dendrophyllia and sponges-Bartonian of the easternmost sector of the Ebro Basin. Cour Forsch-Inst Senckenberg 172:265–273
Calzada S, Astibia H (1996) Sobre Plicatula pamplonensis Carez: revisión. Bull Soc Hist Nat Toulouse 132:65–69
Carez L (1881) Étude des terrains crétacés et tertiaires du Nord de l’Espagne. Savy Edit, Paris
Catullo TA (1856) Dei terreni di sediment superiore delle Venezie e dei fossili bryozoari, antozoari e spongiari. Padova
Charbonnier S, Vannier J, Gaillard C, Bourseau JP, Hantzpergue P (2007) The La Voulte Lagerstätte (Callovian): evidence for a deep water setting from sponge and crinoid communities. Palaeogeogr Palaeoclimatol Palaeoecol 250:216–236
Costa E, Garcés M, López-Blanco M, Serra-Kiel J, Bernaola G, Cabrera L, Beamud E (2012) The Bartonian-Priabonian marine record of the eastern South Pyrenean foreland basin (NE Spain): A new calibration of the larger foraminifers and calcareous nannofossil biozonation. Geol Acta. doi:10.1344/105.000001779
d’Achiardi A (1866) Coralli fossili del terreno nummulitico dell’Alpi Venete. Mem Soc Italiana Sci Nat Milano Part 1, 2(4):1–53
d’Achiardi A (1868) Coralli fossili del terreno nummulitico dell’Alpi Venete. Mem Soc Italiana Sci Nat Milano Part 2, 4(1):1–31
d’Achiardi A (1875) Coralli Eocenici del Friuli. Atti Soc Tosc Sci Nat 1:1–102
d’Archiac AM (1846) Description des fossiles recueillis par M. Thorent dans les couches à nummulites des environs de Bayonne. Mém Soc géol France 2e sér 2:189–217
d’Archiac AM (1850) Description des fossiles du groupe nummulitique recueillis par M.S.-P. Pratt et M.J. Delbos aux environs de Bayonne et de Dax. Mém Soc géol France 2e sér 3:397–456
de Bouillé R (1876) Paléontologie de Biarritz et de quelques autres localités des Basses-Pyrenées. Imprimerie et Lithographie Veronese, Pau
de Buffrénil V, Astibia H, Pereda Suberbiola X, Berreteaga A, Bardet N (2008) Variation in bone histology of Middle Eocene sirenians from western Europe. Geodiversitas 30:425–432
Deshayes GP (1864) Description des animaux sans vertèbres découverts dans le Basin of Paris pour servir de supplément à la description des coquilles fossiles des environs of Paris comprenant une revue générale de toutes les espèces actuellement connues, 2. Baillière et fils, Paris
Dickson JAD (1965) A modified technique for carbonates in thin section. Nature 205:587
Fernández López S (1990) El significado de la autoctonía/aloctonía tafonómica. In: Fernández López S (ed) Comunicaciones Reunión Tafonomía y Fosilización. Universidad Complutense de Madrid, Madrid, pp 115–124
Fernández López S (1991) Taphonomic concepts for a theoretical biochronology. Rev Espan Paleont 6:37–49
Jones RW, Charnock MA (1985) “Morphogroups” of agglutinating foraminifera. Their life positions and feeding habits and potential applicability in (paleo)ecological studies. Rev Paléobiol 4:311–320
Li Q, McGwran B, Boersma A (1995) Early Palaeocene Parvularugoglobigerina and late Eocene Praetenuitella: does evolutionary convergence imply similar habitat? J Micropalaeont 14:119–134
Malumián N, Jannou G, Náñez C (2009) Serial planktonic foraminifera from the Paleogene of the Tierra del Fuego Island, South America. J Foram Res 39:316–321
Matteucci R, Russo A (2005) The Middle Eocene siliceous sponges from Val di Chiampo (Lessini Mountains, northern Italy). Ann Univ Studi Ferrara, Museol Scient Natur vol spec, pp 51–62
Michelin H (1840–1847) Iconographie zoophytologique. Description par localités et terrains des polipiers fossiles de France et pays environnants. P Bertand, Libraire de la Soc géol France, Paris, 1:1–348
Moret L (1924) Contribution à l’étude des spongiaires siliceux du Miocènede l’Algérie. Mém Soc géol France 1:1–27
Moret L (1925) Sur quelques spongiaires de Catalogne (Argovien, Sénonien, Eocène). Bull Soc cie nat Barcelona “Club Muntanyenc” 9:1–11
Murray JW (2006) Ecology and applications of benthic foraminifera. Cambridge University Press, Cambridge
Murray JW, Alve E, Jones BW (2011) A new look at modern agglutinated benthic foraminiferal morphogroups: their value in palaeoecological interpretation. Palaeogeogr Palaeoclimatol Palaeoecol 309:229–241
Mutti E, Tinterri R, di Biase D, Fava L, Mavilla N, Angella S, Calabrese L (2000) Delta-front facies associations of ancient flood-dominated fluvio-deltaic systems. Rev Soc Geol España 13:165–190
Mutti E, Tinterri R, Benevelli G, di Biase D, Cavanna G (2003) Deltaic, mixed and turbidite sedimentation of ancient foreland basins. Mar Petrol Geol 20:733–755
Oppenheim P (1901) Über einige Alttertiäre Faunen der österreichsich-ungarischen Monarchie. Beitr Paläont Geol Österr-Ungarn Orient 13:1–227
Oppenheim P (1912) Neue Beiträge zur Eozänfauna Bosniens. Beitr Paläont Geol Österr-Ungarn Orient 25:87–149
Osasco E (1902) Contribuzione allo studio dei coralli cenozoici del Veneto. Palaeontograph Italiana Mem paleont 8:99–120
Pacaud JM, Le Renard J (1995) Révision des Mollusques paléogènes du Bassin de Paris. IV: liste systématique actualisée. Cossmanniana 3:151–187
Pisera A, Busquets P (2002) Eocene siliceous sponges from the Ebro Basin (Catalonia, Spain). Geobios 35:321–346
Pisera A, Levi C (2002) “Lithistid” Demospongiae. In: Hooper JNA, Van Soest RWM (eds) Systema Porifera. Kluwer Academics/Plenum Press, New York, pp 299–301
Pisera A, Cachão M, Marques de Silva C, Marques de Silva C (2006) Siliceous sponge spicules from the Miocene Mem Moniz marls (Portugal) and their environmental significance. Riv Italiana Paleont Stratigr 112:287–299
Plaziat JC (1981) Late Cretaceous to Late Eocene paleogeographic evolution of southwest Europe. Palaeogeogr Palaeoclimatol Palaeoecol 36:263–320
Pomel A (1872) Paléontologie de la province d’Oran, 5. Spongiaires. 256 p
Puigdefàbregas C (1975) La sedimentación molásica de la cuenca de Jaca. Pirineos 104:1–118
Pujalte V, Baceta JI, Payros A (2002) Tertiary: western Pyrenees and Basque-Cantabrian region In: Gibbons W, Moreno T (eds) The geology of Spain. Geol Soc. London, pp 293–301
Ramos-Guerrero E, Busquets P, Álvarez G, Vilaplana M (1989–1990) Fauna coralina de las plataformas mixtas del Paleógeno de las Baleares. Boll Soc Hist Natur Balears 33:9–24
Reig JM (1988) Tres nuevos géneros y varias especies de Madreporarios fósiles procedentes del Eoceno del Nordeste de España. Published by the author, Barcelona
Reig JM (1990) Madreporarios Eocénicos de Castellolí y de la Sierra de Malvals. Published by the author, Barcelona
Reuss AE (1847) Die fossilen Polyparien des Wiener Tertiärbeckens. Naturwiss Abhandl 2:1–109
Rouault A (1850) Description des fossiles du terrain éocène des environs de Pau. Mém Soc géol France 2e sér 3:457–507
Ruiz de Gaona M (1947) El Bartoniense de la Cuenca de Pamplona. Not Com Inst Geol Min España 17:155–165
Ruiz de Gaona M, Colom G (1950) Estudios sobre las sinecias de los foraminíferos eocénicos de la vertiente meridional del Pirineo (Cataluña-Vizcaya). Estud Geol 12:293–434
Russo A (1979) Studio monografico sui Coralli dell’Eocene di Possagno (Treviso, Italia). Atti Mem Accad Naz Sci Lett Arti Modena 21:1–87
Serra-Kiel J, Hottinger L, Caus E, Drobne K, Ferrández C, Jauhri AK, Less G, Pavlovec R, Pignatti J, Samsó JM, Schaub H, Sirel E, Strougo A, Tambareau Y, Tosquella J, Zakrevskaya E (1998) Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bull Soc géol France 169:281–299
Serra-Kiel J, Mató E, Saula E, Travé A, Ferràndez-Cañadell C, Busquets P, Samsó MJ, Tosquella J, Barnolas A, Àlvarez-Pérez G, Franquès J, Romero J (2003) An inventory of the marine and transitional Middle/Upper Eocene deposits of the Southeastern Pyrenean Foreland Basin (NE Spain). Geol Acta 1:201–229
Solé Sabarís LL (1942) Fauna coralina del Eoceno catalán. Mem Real Acad Cie Art Barcelona 26:259–440
Tabachnik KR (1991) Adaptation of the hexactinellid sponges to deep-sea life. In: Reitner J, Keupp H (eds) Fossil and recent sponges. Springer, Berlin, pp 378–386
Taberner C, Marshall JD, Hendry JP, Pierre C, Thirlwall MF (2002) Celestite formation, bacterial sulphate reduction and carbonate cementation of Eocene reefs and basinal sediments (Igualada, NE Spain). Sedimentology 49:171–190
Vacelet J (1988) Indication de profondeur données par les Spongiaires dans les milieux benthiques actuels. Géol Méditerr 15:13–26
West I (2002) Barton and Highcliffe, Eocene strata: geology of the south coast of England. http://www.soton.ac.uk/~imw/barton.htm (School of Ocean and Earth Sciences, Southampton University, U.K. Version U.04.02.02)
Zlatarski V (1963) Sur Stylocoenia taurinensis (Michelin), madréporaire du tertiaire méditerranéen. Ann Univ Sofia 56:61–71
Acknowledgments
Financial support was provided by the Gobierno Vasco/Eusko Jaurlaritza (Research Group IT-320-10 for HA and JE) and the Ministerio de Ciencia e Innovación, Spain (CGL2010 18851/BTE and CGL2011-23770 for AP and SO). Comments by Dr. P. Busquets (Universitat de Barcelona) and Dr. R. Matteucci (Università “La Sapienza”) considerably improved the manuscript. Technical and human support provided by SGIker (UPV/EHU, MICINN, GV/EJ, ERDF and ESF) is gratefully acknowledged. Mr. Rupert Glasgow has revised the English translation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Astibia, H., Elorza, J., Pisera, A. et al. Sponges and corals from the Middle Eocene (Bartonian) marly formations of the Pamplona Basin (Navarre, western Pyrenees): taphonomy, taxonomy, and paleoenvironments. Facies 60, 91–110 (2014). https://doi.org/10.1007/s10347-013-0364-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-013-0364-2