
Abstract
Subfossil azoxanthellate deep-sea coral mounds occur at 355–410 m on the continental slope of the NE Tyrrhenian Sea between Gorgona and Capraia islands, Tuscan Archipelago. These low-relief patch reefs are at present buried by a thin muddy drape. Their age is latest Pleistocene. The colonial scleractinian Madrepora oculata is the major frame builder, in association with the solitary coral Desmophyllum dianthus and the colonial coral Lophelia pertusa. These NE Tyrrhenian Madrepora-dominated coral mounds represent one of the few known Mediterranean examples of deep-coral colonization of a muddy, low-gradient continental slope.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Submerged deep-sea coral bioconstructions and related sedimentary deposits are widespread in the Mediterranean basin (Taviani et al. 2004, 2005). The Tyrrhenian Sea with an area of ca 247,000 km2, depths exceeding 3,000 m, and irregular submarine topography, offers many chances to successful suitable habitats and environmental conditions for scleractinian growth in the deep-sea. In fact, deep-water corals, especially Lophelia pertusa, Madrepora oculata , Dendrophyllia cornigera and Desmophyllum dianthus (syn. D. cristagalli) have been recorded at various localities, even if many such records refer to non-living or clearly subfossil occurrences (e.g., Segre 1959; Segre and Stocchino 1969; Selli 1970; Sartori 1980; Zibrowius 1980; Rocchini 1983; Bonfitto et al. 1994; Terreni and Voliani 1995). Most Mediterranean occurrences of azooxanthellate deep-water corals relate to sediment-starved situations characterized by rocky bedrock such as seamount/bank flanks and edges, canyon heads and walls, steep cliffs etc. Records of deep-coral mounds developed on firm sedimentary substrata are considerably scarcer. Forest and Cals (1977) first recorded dead and living Madrepora oculata from 450 m off the islands of Gorgona and Capraia, Tuscan Archipelago. Scientific information by ecologists belonging to ARPAT environmental agency (Agenzia regionale per la protezione ambientale della Toscana) confirms the presence of deep-sea corals in this area and the disturbance caused by coral framework to commercial fishing operations (F. Serena 1999, personal communication). Furthermore, there is anecdotal information from commercial fishermen and amateur malacologists on the potential existence of unspecified living “white corals” (sensu Pérès and Picard 1964) in this area.
In the context of the systematic exploration of the Mediterranean Sea conducted by the CNR of Bologna since 1995 to identify, image and sample deep coral settings, a sector of the NE Tyrrhenian Sea was targeted to search for such mounds. A geo-biological survey of the NE Tyrrhenian Sea (cruise LM-99 Lophelia -Millenium) was then conducted in winter 1999–2000 aboard RV Urania (Taviani and Remia 2001; Remia et al. 2004). Another survey of this same area was conducted in winter 2003–2004 (cruise CORTI of RV Urania) but only some preliminary data are included in the present article.
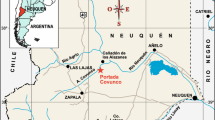
Shallow-buried low-relief deep-coral mounds were identified along the muddy continental slope between Gorgona and Capraia islands, Tuscan Archipelago and are discussed here (Fig. 1).

Location map of the coral mound area in the Tuscan Archipelago discussed here with sampling stations and selected seismic tracks. Bathimetry is from IBCM (International Bathymetric Chart of the Mediterranean)
Material and methods
Positioning during Cruises LM-99 and CORTI was provided by GPS navigational system installed on RV Urania. High-resolution seismic imaging was achieved using Chirp-Sonar (frequency 2–7 kHz), integrated by DESO-25 (frequency 12 kHz), echo-sounding. Sampling was undertaken by using a large-volume (65 l) modified Van Veen grab, epibenthic and heavy-chained dredges. Successful sampling stations, dredging tracks and related profiles are reported in Fig. 2 and Table 1.

Seismic profiles across the coral mound area reported in Fig. 1; A Line A-B DESO-25 (12 kHz); B Line C-D chirp-sonar. TWTT (ms) is two-way travel time. Depth (m) is calculated with sound speed (1,500 m/s)
Results
Coral mounds punctuate a virtually flat muddy bottom on the upper slope between 355 and 410 m offshore Gorgona and Capraia islands at c. 43°13 N and 09°36 E (Fig. 1). These mounds display a patchy distribution and are acoustically detectable. For instance, the mounds are visible on both chirp-sonar profiles and DESO-25 (frequency 12 kHz) acoustic images (Fig. 2). As seen on Chirp line C–D (Figs. 1, 2) the coral buildups occur as discrete mounds many tens of meters wide and up to 3–4 m in height. Direct sampling (grab and epibenthic dredge) provided evidence that these coral mounds are at present buried only by a thin veneer of soft mud and are at places still exposed. Large-volume grab samples have consistently documented that corals still embedded in mud are well preserved and white, while those exposed on the sea bottom are deeply bioeroded, Mn-Fe patinated and exploited by a number of dwellers (zoozoans sensu Taylor and Wilson 2002).
As demonstrated by sampling transects, not much coral rubble is present around any single coral mound. The coral assemblages show the absolute dominance of fresh-looking Madrepora oculata branches (Fig. 3c), and Desmophyllum dianthus (Fig. 3a, b) with the corallites still retaining their original lustre. Subfossil branches belonging to Lophelia pertusa are less common (Fig. 3d). The largest single piece of dredged Madrepora colony attains a size of 26 cm (Fig. 3c). Early diagenetic limestones (wackestones, packstones and framestones) encasing biostromal coral rubble are occasionally recovered from this area.

Dominant species from the coral mound area: a,b Desmophyllum dianthus (Esper 1794), large well-preserved specimen under study for trace and minor element geochemistry; c well-preserved Madrepora oculata Linnaeus, 1758, largest piece of coral framework dredged; d Lophelia pertusa Linnaeus, 1758, fragment of coral colony; all from st. LM99–124
Diversity and taphonomy
Ecological work on living Lophelia mounds in the Eastern Atlantic Ocean has clearly demonstrated that such environments promote a significant increase of biodiversity with respect to the surrounding sea bottom (e.g., Jensen and Frederiksen 1992; Freiwald 1998, 2002; Freiwald et al. 2004). However, only a few faunal elements seem strictly related to such deep-coral buildups and, among them, only a handful of taxa actually settle on living parts of the coral reefs while the number of exploiters of coral substrata substantially increases on dead portions of these coral mounds suggesting complicated patterns of taphonomic turnovers (Freiwald 1998; Freiwald and Wilson 1998).
Taphonomically, these Tyrrhenian coral mounds (Figs. 4, 5) share many analogies with E Atlantic counterparts. Most corals appear encrusted by subfossil valves of Spondylus gussonii, serpulid polychaete tubes and bryozoan colonies. Various living organisms were also observed on degraded and patinated corals and coral framestones including hydroids, sponges, bivalves (e.g., Asperarca nodulosa), polychaetes (e.g., Metavermilia multicristata), brachiopods (e.g., Neocrania sp., Terebratulina retusa), etc. (Fig. 6). Extra-calcification as a response to parchment tubes of the commensal polychaete Eunice is common on both Madrepora and Lophelia (Fig. 5). Pitted scars possibly attributable to parasitic epibiont foraminifers have been observed on some Lophelia branches. Other scars found on Madrepora and Lophelia related to cemented brachiopods (Neocrania sp.).

Taphonomic aspects; A highly encrusted and endolithic-bioeroded Madrepora colony, st. LM99–124; B the bivalve Coralliophaga lithophagella (Lamarck 1819) nestling inside an Eunice tube-gall in Lophelia, st. LM99–124; C pitted scar of a benthic foraminifer ( arrow), st. LM99–124; D valve of Spondylus gussonii with an extraordinarily developed hinge cemented to an Eunice-related tube-gall, st. CORTI-29; E scar related to the brachiopod Neocrania sp. and fouling serpulids and bryozoans, st. LM99–124

Taphonomic aspects; A coral wackestone, (mi) early diagenetic micritic cement and (se) pockets of still unlithified muddy sediment (sample recovered off Gorgona Island by the trawler Franci); B loosely cemented coral packstone encasing articulated brachiopod shells belonging to Gryphus vitreus (Born 1778), st. CORTI-29; C Madrepora coral colony with an in situ cemented shell of the predatory gastropod Coralliophila squamosa (Bivona 1838), st. CORTI-57; D Madrepora coral packstone encasing a shell of the limpet Emarginula sp., st. CORTI-29; E loosely cemented coral framestone encasing an articulated brachiopod shell belonging to Megerlia truncata (Linnaeus 1767), st. CORTI-57

Associated calcareous invertebrates; A attached valves belonging to the bivalve Spondylus gussonii Costa, 1829, st. LM99–124; B tube galls and parchment tubes of the commensal polychaete Eunice (st. LM99–124); C fresh solitary coral Stenocyathus vermiformis (Pourtalès 1868) attached to a highly degraded and patinated coral substrate, st. CORTI-56; D a living specimen of the bivalve Asperarca nodulosa (Müller 1776) byssated on a subfossil valve of S. gussonii, st. CORTI-29; E living specimen of the serpulid polychaete Metavermilia multicristata (Philippi 1844), one of the commonest members of the attached epifauna, st. CORTI-56; f: Neocrania sp. (Müller 1776) attached to a highly degraded and patinated coral substrate, st. CORTI-57
A collection of skeletal invertebrates was obtained through sampling of Madrepora-mounds, including 10 Scleractinia (Table 2). Such an invertebrate assemblage does not necessarily represent the actual biodiversity of the original deep-coral mound.
Sampling on and around the mounds provided many skeletal taxa, most of which are indicative of soft clay coral-free bottom. Among macrobenthic invertebrates, Mollusca are by far the most diverse taxonomic group with 140 species (44 bivalves, 88 Gastropoda, 3 Polyplacophora and 5 Scaphopoda that will be treated in a separate paper), followed by Cnidaria (Scleractinia, Octocorallia), Annelida (Serpulidae: Protula sp., Serpula spp., Metavermilia multicristata, Semivermilia spp., Filogranula spp., Placostegus tridentatus, Vermiliopsis cf. infundibulum, Ditrupa arietina; Spirorbidae: Janua sp., cf. Pileolaria sp.), Brachiopoda (Neocrania sp., Megerlia truncata, Gryphus vitreus, Terebratulina retusa, Platidia anomioides, P. davidsoni), Crustacea Decapoda (e.g. Ebala), Echinoidea (e.g. Cidaris sp.). Microbenthos is dominated by Foraminifera (e.g. Ammonia aff. beccarii, Elphidium spp., Bigenerina nodosaria, Textularia sp., Brizalina sp., Sigmoilina spp., Cornuspira involvens, Lenticulina sp., Dentalina spp., Pyrgo spp., Quinqueloculina sp., Bulimina inflata, B. marginata, Cassidulina spp., Planulina ariminensis, Paromalina coronata, Stilostomella spp, Cibicides lobulatus, Cibicidoides sp., Lagenodosaria scalaris, Praeglobobulimina pupoides, Hyalinea balthica, Saracenaria italica, Uvigerina mediterranea, U. peregrina, U. proboscidea and agglutinated species) and Ostracoda (e.g., Bythocypris sp., Polycope sp., Paracytherois sp., Henryhowella sp. and Argilloecia acuminata).
Age
One specimen of Desmophyllum dianthus from station LM99–124 provided an U/Th age of 11.1±0.1 ka (Research School of Earth Sciences, Australian National University, Canberra, Australia). Associated fauna includes, amongst others, abundant glacial-Pleistocene subfossil valves of the scallop Pseudamussium septemradiatum (e.g., Malatesta and Zarlenga 1986). Further U/Th and AMS–14C dating of corals is in progress, but there is little doubt that such mounds thrived during the last glacial epoch.
Impact by commercial trawling
Deep-sea trawling operations are routinely conducted in this sector of the Tyrrhenian Sea. Unlike the Atlantic Ocean (e.g., Hall-Spencer et al. 2002; Freiwald et al. 2004), Mediterranean deep-coral banks are not a target for and therefore are not deliberately impacted by any commercial fishing. On the contrary, they represent a type of bottom that trawlers try carefully to avoid in order not to damage their nets. Fishing-boat echo-sounders are capable of indicating the likely presence of coral mounds. The experience gained by the accidental entangling of nets with coral colonies has greatly reduced such accidents among commercial fishermen (A. Voliani 2003, personal communication). Notwithstanding such observations, deep-coral rubble is almost ubiquitous in any trawling catch from this area because of the patchy presence of shallow-buried corals and the perpetual re-collection of corals discarded by previous trawling activity.
Discussion
While most known submarine deep coral buildups in the Mediterranean basin colonized sediment-starved rugged topographic highs (seamounts, banks, canyons, steep overhangs etc.), these Tyrrhenian mounds represent an example of corals developed on firm sedimentary units contributing to a gently dipping continental slope. The coral mounds under scrutiny may be categorized as patch reefs in the sense proposed by Wilson (1979) and Freiwald (2002). Similar bio-constructions are not uncommon in the modern oceans (e.g., Moore and Bullis 1960; Allen and Wells 1962; Stetson et al. 1962; Squires 1965; Mullins et al. 1981; Newton et al. 1987) and in the fossil record (e.g., Squires 1964). In the Mediterranean basin, the healthy Lophelia -reef reported from the Ionian Sea are another example of successful colonization of a prevalently non-rocky substratum on a gently sloping margin (Taviani et al. 2004).
It appears that deep coral growth along this sector of the Tyrrhenian slope was severely reduced at the end of the Pleistocene, c. 11 ka ago. The postglacial basinal decline of deep-sea corals in the Mediterranean Sea has been noticed long ago (Taviani and Colantoni 1984; Delibrias and Taviani 1985). We observe that these Tyrrhenian mounds were rapidly buried by a thin veneer of mud and we tentatively suggest that this silting event is possibly a response to sediment availability due to an Early Holocene climatically-forced strengthening of continental runoff. Planktonic foraminifer assemblages recovered from the mud draping the coral mounds contain Globigerinoides ruber (pink), G. sacculifer , G. trilobus, Globigerina precalida, G. predigitata and are indicative of warm water (possibly the Holocene Climatic Optimum), together with fresh-temperate forms such as Globorotalia truncatulinoides, G. inflata, Neogloboquadrina pachyderma, Globigerina quinqueloba, G. bulloides (A. Borsetti 2003, personal communication). Coexistence of such forms may be taken as an indication that mud-burial of the coral mounds started at the very end of the Pleistocene but that its peak took place between 6,000–9,000 years ago.
References
Allen JRL, Wells JW (1962) Holocene coral banks and subsidence in the Niger Delta. J Geol 70:381–397
Bonfitto A, Bigazzi M, Fellegara I, Impiccini R, Gofas, S, Oliverio M, Taviani M, Taviani N (1994) Rapporto scientifico sulla crociera DP 91 (Margine orientale della Sardegna, Mar Mediterraneo). Boll Malacolog 30:129–140
Delibrias G, Taviani M (1985) Dating the death of Mediterranean deep-sea scleractinian corals. Mar Geol 62:175–180
Forest J, Cals P (1977) Une deuxième espèce du genre Bresilia Calman, B. corsicana sp. nov. Comparaison avec B.atlantica Calman (Crustacea Decapoda Bresiliidae). Bull Mus Natl Hist Nat 3 ser., 453:549–565
Freiwald A (1998) Geobiology of Lophelia pertusa (Scleractinia) reefs in the North Atlantic. Habilitationsschrift, Fachbereich Geowissenschaften der Universität Bremen, pp 116
Freiwald A (2002) Reef-forming cold-water corals. In: Wefer G, Billett D, Hebbeln D, Jørgensen BB, Schlüter M, Van Weering T (eds) Ocean margin systems. Springer, Berlin Heidelberg New York, pp 365–385
Freiwald A, Fosså JH, Grehan A, Koslow T, Roberts JM (2004) Cold-water coral reefs. UNEP-WCMC Report, Biodiversity Series 22:1–84
Freiwald A, Wilson JB (1998) Taphonomy of modern deep, cold-temperate water coral reefs. Hist Biol 13:37–52
Hall-Spencer J, Allain V, Fosså JH (2002) Trawling damage to northeast Atlantic ancient coral reefs. Proc R Soc Lond B 269:507–511
Jensen A, Frederiksen R (1992) The fauna associated with the bank-forming deepwater coral Lophelia pertusa (Scleractinia) on the Faroe shelf. Sarsia 77:53–69
Malatesta A, Zarlenga F (1986) Northern guests in the Pleistocene Mediterranean Sea. Geol Romana 25:91–154
Moore DR, Bullis HT Jr (1960) A deep-water coral reef in the Gulf of Mexico. Bull Mar Sci 10:125–128
Mullins HT, Newton CR, Heath K, Vanburen Hm (1981) Modern deep-water coral mounds north of Little Bahama Bank: Criteria for recognition of deep-water coral bioherms in the rock record. J Sediment Petrol 51:999–1013
Newton CR, Mullins HT, Gardulski AF (1987) Coral mounds on the Western Florida Slope: unanswered questions regarding the development of deep-water banks. Palaios 2:359–367
Pérès JM, Picard J (1964) Nouveau manuel de bionomie benthique de la Mer Méditerranée. Rec Trav Stat Mar End. 31:1–137
Remia A, Montagna P, Taviani M (2004) Submarine diagenetic products on the sediment-starved Gorgona slope, Tuscan Archipelago (Tyrrhenian Sea). Chem Ecol 20(1):S131–S153
Rocchini R (1983) Acesta excavata (Fabricius 1779), nuovo ritrovamento in Mediterraneo. Boll Malacolog 19:83–86
Sartori R (1980) Factors affecting the distribution of ahermatypic corals on the Mediterranean seafloor: a probabilistic study. Deep-Sea Res 27:655–663
Segre AG (1959) Observations générales sur l’orographie sous-marine de la Mer Tyrrhénienne. In: CNRS, La Topographie et la Géologie des profondeurs océaniques, Colloq.Intern. du CNRS 83:53–59
Segre AG, Stocchino C (1969) Nuove osservazioni sulla geologia e morfologia delle montagne submarine del Mar Tirreno. Ist. Idrogr. Marina Genova 1037:11–15
Selli R (1970) Campioni raccolti. Giorn Geol 37:55–72
Squires DF (1964) Fossil coral thickets in Wairarapa, New Zealand. J Paleontol 38:904–915
Squires DF (1965) Deep-water coral structure on the Campbell Plateau, New Zealand. Deep-Sea Res 12:785–788
Stetson TR, Squires DF, Pratt RM (1962) Coral banks occurring in deep water on the Blake Plateau. Am Mus Novitates 2114:20–39
Taviani M, Colantoni P (1984) Paléobiocoenoses profondes à Scléractiniaires sur l’escarpement de Malte-Syracuse (Mer Méditerranée): leur structure; leur âge et leur signification. Rev Inst Franc Pétr 39:547–559
Taviani M, Remia A, Corselli C, Freiwald A, Malinverno E, Mastrototaro F, Savini A, Tursi A (2004) First geo-marine survey of living cold-water Lophelia reefs in the Ionian Sea (Eastern Mediterranean). Facies (2004)
Taviani M, Freiwald A, Zibrowius H (2005) Deep-coral growth in the Mediterranean Sea: an overview. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, Berlin Heidelberg New York
Taviani M, Remia A (2001) I coralli del buio: archivi climatici degli oceani passati. Ricerca e Futuro 18:28–30
Taylor PD, Wilson MA (2002) A new terminology for marine organisms inhabiting hard substrates. Palaios 17:522–525
Terreni G, Voliani A (1995) New finding of Acesta excavata (Fabricius 1779) in the northern Tyrrhenian Sea. La Conchiglia 276:13–14
Wilson JB (1979) ‘Patch’ development of the deep-water coral Lophelia pertusa (L.) on Rockall Bank. J Mar Biol Assoc UK 59:165–177
Zibrowius H (1980) Les Scléractiniaires de la Méditerranée et de l’Atlantique nord-oriental. Mém Inst Océanogr Monaco 11, 227 pp
Acknowledgements
We thank captains Vincenzo Lubrano and Emanuele Gentile, officers, crew and colleagues onboard RV Urania during cruises LM99 and CORTI for their cooperation. Thanks are also due to the brothers Merlin, owners of the trawler Franci (Livorno), as well as to Fabrizio Serena and Alessandro Voliani (both ARPAT, Tuscany) for their information regarding deep coral mounds offshore the Tuscan Archipelago. We are indebted to Barbara Gualandi and Stefano Parisini (both CNR, Bologna) and Bruno Sabelli (University of Bologna) for producing digital documentation of LM99 and CORTI samples. Luca Gasperini (CNR, Bologna) helped with seismic processing. We are indebted to Malcolm McCulloch (Research School of Earth Sciences, Australian National University, Canberra) for dating of the specimen of D. dianthus. Systematic expertise was provided by Helmut Zibrowius (University of Marseille: scleractinians and polychaetes); Anna Maria Borsetti and Lucilla Capotondi (CNR, Bologna: foraminifers) and Nevio Pugliese (University of Trieste: ostracods). Cruises LM99 ( Lophelia-Millennium) and CORTI were funded by CNR. We thank John B. Wilson for the constructive review of the manuscript. This is a contribution to MIUR SINAPSI and ESF Euromargins MOUNDFORCE programmes. IGM scientific contribution n. 1390. This work is dedicated to the memory of the late Luciano Casoni, a friend, a skilful marine technician and a companion during cruise LM99 and other coral missions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Remia, A., Taviani, M. Shallow-buried Pleistocene Madrepora-dominated coral mounds on a muddy continental slope, Tuscan Archipelago, NE Tyrrhenian Sea. Facies 50, 419–425 (2005). https://doi.org/10.1007/s10347-004-0029-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-004-0029-2