
Abstract
Home-range sizes, movements, and daily activity of wolves (Canis lupus L. 1758) were studied in Dalmatia, Croatia in 1998–2001. The total home ranges (100% MCP) of two packs were 160 km2 and 141 km2, mean=150.5 km2. Core areas (50% kernel) were 26.2 km2 and 3.3 km2, respectively. Differences in core area sizes were influenced by human activity—hunting and sheep grazing. Compared with random locations, wolf locations were closer to the nearest water source (mean=937 m) and farther from houses (mean=653 m). Wolves were significantly more active during the night than during the day (activity indexes were 0.53 vs. 0.35), and night activity was higher during summer (0.58), and lower during winter (0.48). A correlation was found between distances traveled and activity index (r=0.58, p=0.003). Home range, seasonal variations in home-range size, habitat use, and activity of wolves in Dalmatia were oriented to make the compromise from danger of proximity to humans and also to benefit from human-related food sources.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Radiotracking remained the method of choice during last 35 years for studying species, as elusive as wolf (Canis lupus L. 1758) (Mech et al. 1998). Home-range sizes, movements, habitat use, activity, and other aspects of wolf ecology were intensively studied in North America (Mech 1970; Fritts and Mech 1981; Messier 1985; Potvin 1988; Fuller 1991; Mech 1992, 1994; Mech et al. 1998). In Europe, Okarma et al. (1998) described results of wolf telemetry study in an area with abundant and diverse natural prey, and synthesized results of all previous wolf home-range studies covering Europe and former Soviet Union. Other European studies focus on home ranges, movements, and activity of wolves living in areas intensively used by humans (Vilà et al. 1990, 1995; Ciucci et al. 1997), and in areas less used by people (Theuerkauf et al. 2003a, b). Recently, wolves have started re-colonizing parts of their former range in Europe, appearing in areas with rather intensive human use (Promberger and Schröder 1993). In such areas, information on wolf activity, space, and habitat use can be obtained only by means of radiotelemetry, and this information is often critical for successful conservation of the species.
We studied use of space, movements and activity of wolves in the mediterranean part of Dinarids, Dalmatia. The area represents the Southeastern end of wolf distribution in Croatia (Kusak 2002). After the legal protection of wolves in Croatia in 1995 (Anonymous 1995), the highest number of requests for wolf damage compensation came from Dalmatia (Kusak 2002). Dalmatia comprises only one-third of the total wolf distribution range in Croatia, but includes 97% attacks of wolves on livestock, and a significant number of illegal killing of wolves happen there (Huber et al. 1999). In the late 1980 s, it was believed that wolves do not permanently inhabit Dalmatia (Frković and Huber 1992), and there was no other information on wolves in this area. During 1990 s, wolves became increasingly present, followed by a growing animosity of the public that urged need for information to support sound management decisions. A radiotelemetry study of wolves in Dalmatia was part of a broader project on the entire wolf range in Croatia where research of distribution, numbers, population trends, food habits, influence on livestock, mortality, and habitat suitability was conducted. Objectives of our radiotelemetry investigation were to assess (i) home-range sizes, (ii) movements, and (iii) daily activity patterns of the wolves.
Materials and methods
Study area
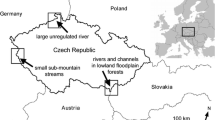
The study area (43°35′N–43°53′N and 16°07′E–16°35′E, Fig. 1) is situated in inner Dalmatia, which represents the southern part of the Dinaric range in Croatia. The part of Dalmatia with the highest density of wolf attack on livestock (Huber et al. 1999) was chosen as the study area. The relief features are typical for limestone karst: sharp bare rocks, cliffs, valleys, caves, sinkholes, and underground streams. Soils are mainly of poor quality and can be found in valleys and between rocks (Božičević 1995). The climate is characterized with hot and dry summers, with the average temperature above 22°C. Winters are mild and with rain, while snow is a rare and short-lasting event (Seletković and Katušin 1992). Forest communities of the area are submediterranean and epimediterranean, thermophile deciduous forests of mediterranean oak and hornbeam (Carpino orientalis and Quercetum virgiliane). Those forests were subject to intensive degradation caused by humans during several thousand years. They were used as the source of firewood and as forest pastures. Soil on southern slopes was eroded, leaving open areas with bare rocks and some soil with grass in between. Northern slopes and some valleys remained covered with degraded and regenerating stages of dense, shrubby remains of autochthonous vegetation—chaparral (Rauš et al. 1992).

Study area located in southeastern part of Croatia, Dalmatia and representing the southeastern part of wolf distribution range in Croatia
Although scientific data are missing, it can be assumed that there were no resident wolves in Dalmatia in the late 1980 s when the number of wolves in other parts of Croatia was low (Frković et al. 1992; Huber et al. 2002). The common way of sheep husbandry in Dalmatia at that time was to leave flocks alone on open pastures even over night. The human population of the Dalmatian inland remarkably decreased from 1960 onward (Korenčić 1979), and this trend has continued until recent times (Borovac 2002). Livestock number was decreasing too, with consequent recovery of autochthonous mediterranean oak and hornbeam forests. Many slopes, former pastures with some grass and bare rocks, are now overgrown by dense stands of pioneering prickly juniper bushes (Juniperus oxycedrus). First, and still the only, ungulates which re-colonized the area were wild boars (Sus scrofa) in the early 1980 s. Wolves settled in the area at the beginning of 1990 s, during the war in former Yugoslavia (Kusak 2002). Based on analyses of stomach content of dead wolves and of wolf feces, their diet consisted predominantly (67.7–73.4%) of domestic animals. Wolves in Dalmatia fed on livestock, which they killed and on illegally disposed slaughtery refuses and livestock carcasses (Kusak 2002; Pavlović 2003). No other large carnivores are permanently present in the area, while golden jackal population persists along the coast and on some islands (Kryštufek and Tvrtković 1990).
Field procedures
The current presence of wolves in the area was determined by inspecting recent cases of attack on livestock, scent-marking posts, howling surveys, and by occasional observation while doing fieldwork. Howling surveys were used to determine if radiotracked packs were alone or by members of corresponding packs (Harrington and Mech 1982). Wolf home ranges, movements, and activity were studied by telemetry. Two females and one male wolves were captured, radiocollared, and tracked from October 1998 to July 2001. All three wolf were captured with foothold traps (Rancher’s Supply INC, Alpine, TX, USA, SAD-#7 wolf trap, modified by the authors). Wolves were immobilized by the use of “Zoletil 100” (100 mg/ml of tiletamine and 100 mg/ml of zolazepam, Vibrac Laboratories, 06516 Carros, France) with doses recommended by Ballard et al. (1991a, b). Sex and reproductive status were determined; while age was estimated on the basis of body size, tooth wear (Gipson et al. 2000), and date of capture. Wolves were marked by radiocollars (MOD-500, Telonics Inc., Messa, AZ, USA) and plastic ear tags.
Due to high density of local roads, almost all radiotracking was done by use of a small field vehicle. The routine protocol included 3-day-long telemetry sessions at 10-day intervals throughout the year. During three trapping seasons, which lasted from one to two summer months, the previously marked wolves were located daily. Locations were collected during all 24 h of the day (from 06:00 hrs to 18:00 hrs, we made 57.9% of locations and from 18:00 hrs to 06:00 hrs, 42.1% of locations), with minimum of one location at daytime and one location at night. We successfully found wolves in 89% of all attempts. We used DOS based palmtop PC and “Locate II” program (Nams 1993) set to maximum likelihood estimator (MLE) method (Lenth 1981) to calculate locations and the size of 95% confidence areas. Locations were calculated from at least three bearings in the field immediately. The time interval between the first and last successful bearing was between 20 min and 40 min. Bearings were measured until the desired accuracy (95% confidence area of size ≤0.3 km2) of calculated location was achieved. Calculated locations were mapped, and their distances to the nearest paved or unpaved road leading to inhabited house (excluding forestry roads), water source at traditional ponds and wells for livestock (there are no springs, creeks or lakes in the area), vegetation type (karst pasture, meadow, prickly juniper brush, deciduous forests, and bare rocks), elevation, and exposure were recorded. We used minimum-convex-polygon (MCP, White and Garrot 1990) and kernel methods (Worton 1989) to calculate the size of home ranges. For calculating kernel home ranges, we used fixed kernel method with smoothing factor (bandwidth) calculated by least square cross validation (LSCV) of wolf locations (Seaman et al. 1999). Those calculations were done by the use of “Animal movement” (Hooge et al. 1999) extension for ArcView GIS 3.1 (1996).
To estimate the activity of wolves by radiosignal, we employed a method described by Roth and Huber (1986), which uses the fluctuation in signal strength to determine the activity of the tracked animal. Activity reading was recorded immediately after measuring every bearing. In addition, during each 3-day-session, one 24 h activity monitoring was done, when the activity was determined in time intervals of every 10 min–15 min (maximum four readings per hour were used). For one wolf, one continuous 7-day tracking and activity monitoring was done. Data sets were categorized by animals, and only those with at least two complete 24 h monitoring sessions were used, with a daily average of minimum five activity readings per hour, resulting in a minimum of 240 readings in one set. All activity readings were categorized in four seasons, where spring and summer were considered as warm and autumn and winter as cold seasons. Data on day length, dusk, and dawn, were obtained from U.S. Naval Observatory Astronomical Applications Department, Washington, DC 20392-5420, USA (http://aa.usno.navy.mil/). Data were calculated for the center of the study area (E016°14′, N43°39′), with dawn defined as the period from the moment when the edge of the sun disk was 6° below the horizon to the moment when it touched the horizon at sunrise. Dusk was defined as period starting when the sun disk disappeared from the horizon and until it reached 6° below it during sunset. Night was the period from dusk to dawn; day was from sunrise to sunset (see http://aa.usno.navy.mil/faq/docs/RST_defs.html#top). Because of small sample sizes for dusk and dawn, those data were sometimes merged with night data sets. The following variables were used in wolf activity analyses: date, time, season, animal activity, time of the day (day, night, dusk, and dawn), and relative sun time in radians. Time was converted to radians where sunrise was 0, sunset was π, and the whole day was 2π long. Day and night were assigned the length of π, enabling the comparison of the activity regardless of the real length in hours. The activity index was calculated as the average of all activity (0=inactive, 1=active) during a certain interval. Non-parametrical statistical tests (Mann–Whitney U-test, Spearman’s RHO test, and chi-square test) were used for analyses of activity distribution, correlations, and significance of differences in activity pattern.
Results
The three radiotracked wolves were members of two neighboring packs. This was confirmed by four visual observations, 49 checkings of howlings and by concurrent radiotracking. We saw a group of two wolves three times, and three wolves at one occasion. Wolves replied on howlings 18 times (36.7%). Each time there was more than one wolf howling, and in five (27.7%) cases we distinguished pup voices. In the period from August 23, 1999 to August 3, 2000, two wolves were tracked concurrently (202 locations), and during this time (346 days), they were never found together. The overall tracking effort and data collected are shown in Table 1. Two females from two packs were tracked 643 days and 699 days respectively, exceeding the minimum of 9–12 months of radiotracking needed for reliable home-range estimation (Okarma et al. 1988). Wolf W1 stayed with the pack for 417 days, thereafter it dispersed and those locations (n=23) were not used in home-range calculations. Wolf W2 was a breeding female of the pack during all radiotracking time. Data of W3 were not sufficient for the analyses. Home ranges of two female wolves were 160 km2 and 141 km2, respectively (MCP, 100% of locations, Fig. 2). It was apparent that home ranges of two wolves overlapped on surface of 16 km2 (10.0% of “Opor” pack and 11.3% of “Vučevica” pack territory) but there was a time shift of almost one year. Core areas, comprising 50% and of locations, were calculated by the fixed kernel method. The calculated bandwidth (LSCV) for Opor and Vučevica wolf packs was hLSCV=1595.1 and hLSCV=857.6, respectively. The size of wolf W1 core area was 26.2 km2 (Fig. 2). This comprised 16.4% of calculated MCP home range. Core area of W1 was divided into two major subunits, which W1 used intermittently. The southeastern part was used evenly during warmer and colder part of the year, while the northwestern part was used almost exclusively (99.5% of all locations) during the warmer part of the year. During spring/summer, the size of W1 home range was 86.2 km2, while during autumn/winter it was 91.6 km2 (MCP, 100% of locations, Fig. 2). The size of core areas for W2 was only 3.3 km2, comprising 2.3% of calculated MCP home-range size. This core area was significantly smaller (χ2=15.149, d.f.=1, p<0.01) than the core area of wolf W1. Female wolf W2 used the same core area year-round, but the size of seasonal home ranges varied over time. During spring/summer, the size of her home range was 59.3 km2, while during autumn/winter she used 120.8 km2, the difference being significant (χ2=56.243, d.f.=1, p<0.01). Seasonal differences in home-range use of W1 were influenced by human activities—hunting and sheep grazing. The northwestern part of the core area had open pastures on southern exposures and dense chaparral on northern slopes. Sheep are kept on pastures during the warm season, while they stay in fences close to houses during winter (Kusak 2002). The hunting season was from October to March, and the most common way of hunting was chase hunting with dogs. The northwestern part of W1 core area was the hunting area of a local hunting club, while on southeastern part had no hunting and no sheep grazing because the area was densely forested. Wolf W1 stayed close to sheep when they were on pastures during summer, and moved to another area to avoid hunters during winter. The core area of W2 was situated in valley Vučevica, which was, like the adjacent mountain, densely forested. The valley and the mountain were the largest (65 km2) patch of forest in the whole study area, without any roads or villages. The area was undisturbed by hunting from 1994 until 2000, as it was then leased as hunting ground.

Home ranges (100% MCP and 50% kernel) and seasonal ranges (100% MCP) of two radiotracked wolves in Dalmatia, Croatia. Wolf W1 was tracked from October 30, 1998 to December 21, 1999 and wolf W2 from August 23, 1999 to July 22, 2001
A total of 90 24-h movements for W1 and 93 for W2 were documented during the whole tracking period. The straight distances traveled in 24 h ranged from 0 m to 13.2 km, mean=2.5 km/day, SD=2.9. During one continuous 7-day tracking, day-to-day travels of wolf W2 were 9.4 km/day on average, measured on the 3D surface model as sums of Euclidean distances for consecutive locations, while on 2D map, same trip averaged at 8.9 km/day.
Distances of wolf locations to features in the surrounding environment (roads, houses, water sources, and elevation) were measured on maps, and compared with randomly selected locations. Compared (t-test) with random locations (n=100), wolves preferred to stay closer to the nearest water source (p=0.02), and farther from houses (p=0.03), while other distances were not significantly different from random (Table 2).
In the period from October31, 1998 to July22, 2001 a total of 3234 activity readings were collected for the three wolves. Data from wolf W3 were not sufficient and were excluded, giving the final set of 3182 activity readings for two female wolves. A correlation between sun cycle and wolf activity was found (r=0.389, p<0.01, Spearman’s Rho test, Fig. 3). There was no significant difference in daily activity between W1 and W2 (p=0.365, Mann–Whitney U-test), so the data were pooled. Activity indexes for morning and evening twilights were not significantly different from indexes for the night, but were different from indexes for the day. Twilight indexes were pooled and analyzed together with data for the night. Activity indexes of both wolves were significantly higher during the night than during the day (0.53 vs. 0.35, p<0.001, chi-square test). Night activity was significantly higher during summer (0.58, p=0.007) and significantly lower during winter (0.48, p<0.001), when compared to year-round night activity (0.53, Table 3). Daytime activity in spring (0.42, p=0.050) and autumn (0.37, p=0.027) was significantly higher than year-round daytime activity (0.35), while overall winter activity (0.42, p=0.043) was significantly lower than year-round overall activity (0.44, Table 3). Nightly activity during warm season (0.57) was significantly higher (p<0.001, Mann–Whitney U-test) than night activity during cold season (0.51, Fig. 3).

Overall daily activity, daily activity in warm and cold part of the year of two wolves in Dalmatia, Croatia related to sun cycles
Movement and activity of W2 was continuously monitored in one session during 163 h. A total of 41 location and 639 activity readings were recorded. The sum of Euclidean distances between successive locations of wolf W2 was 64.1 km covering 59.9 km2 (100% MCP), while the overall activity in the same time was 0.42. A correlation between distances traveled and activity index was found with r=0.581, p=0.003 (Spearman’s Rho test, Fig. 4).

The relation of the average distances traveled and activity index of wolf W2 in Dalmatia during a continuous 7-day radiotracking in February 2000. Shown are calculated means for each hour of those 7 days
Discussion
The fact that only two wolf packs were followed advises for caution when trying to apply this data for other wolf populations. Ongoing studies by our group show that even the wolves in another region of Croatia (Gorski kotar) behave differently. Each of two wolves was radiotracked for more than a year, fulfilling the requirements discussed by Okarma et al. (1998) for reliable home-range assessment. Wolves were located, and activity was determined, during all 24 h of the day, ensuring that eventual long excursions over night were not missed. The total amount of collected data is limited, but considering that we were rather efficient in finding animals (89% of all attempts), and that animals were tracked during the day and night (from 06:00 hrs to 18:00 hrs we made 57.9% of locations and from 18:00 hrs to 06:00 hrs 42.1% of locations), and that the tracking periods lasted 643 and 699 days, we are confident that our data represent home ranges of this animals and their corresponding packs. The estimation of wolf numbers in the pack was not reliably possible due to thick vegetation, night activity of wolves and absence of snow cover. However, because of few night sightings and from repeated howling observations, we knew they were not solitary animals, but were pack members. The average home range of two wolf packs in Dalmatia (150.5 km2) falls within the range of home-range sizes for established wolf populations in southern and central Europe, viz. 82–243 km2 (synthesized by Okarma et al. 1998), 197 km2 (Ciucci et al. 1997), and is comparable to home ranges of wolves in Northcentral Minnesota (116 km2, Fuller 1989). The smaller range of wolf W2 during spring/summer confirms earlier findings that reproductive females are bonded to den or rendezvous sites when pups are growing (Okarma et al. 1998; Jędrzejewski et al. 2002).
Daily distances traveled by wolves in Alaska can be up to 80 km/day, depending on prey density and vulnerability (Mech et al. 1995). In different ecological conditions in North America, with higher density of ungulates, wolves travel on average between 1.6 km and 9.0 km/day (Kolenosky and Johnston 1967). In the Białowieża temperate lowland forest (Poland), with a rich community of ungulate species, straight-line wolf movements are 4.4 km, and this is related to prey density (Jędrzejewski et al. 2002). In areas like Dalmatia or central Italy, where wolves rely mainly on human food sources, and do not need to search large areas, those distances are even shorter (3.3 km/day, Abruzzo NP, Ciucci et al. 1997). Distances calculated in this way represent only about 21% of the real daily distances traveled, based on continuous radiotracking (locations at 15–30 min intervals, Jędrzejewski et al. 2002). In our study, this difference was 28.1% (2.5 km/day straight lines versus 8.9 km/day during 7 days of continuous tracking).
Wolves in Dalmatia preferred to stay away from humans, as found of wolves in Białowieża (Theuerkauf et al. 2003a, b). In contrast, Thiel et al. (1998) found that the expanding wolf population in Minnesota, which reached areas used by humans, can be tolerant to human activities close to den or rendezvous sites. Similar distances between wolves and humans can be interpreted as avoidance in one case, or as tolerance in another, depending on the prevailing conditions in a particular area. Wolves on the edge of the expanding Minnesota wolf population recently developed tolerance to human activity. Such a situation is more often found in Europe, where wolves not only tolerate human proximity, but also take advantage of it. Wolves are using garbage dumps in Italy (Ciucci et al. 1997) and Romania (Promberger 1996) or can live in open oat fields, as in Spain (Blanco 1993). Wolves in Dalmatia feed mainly from human sources (livestock, garbage and slaughter-house dumps (Kusak 2002), and because of killing livestock are hardly accepted by locals (Bath and Majić 2000) with consequent killing of wolves whenever possible (Huber et al. 2002). Wolves in Dalmatia were staying closer to water sources, which seem to be one of critical factors in Dalmatia, especially during dry summer months. Those water sources are mostly man-made (ponds and wells) and used to provide livestock with water for many centuries. Wolves in Deccan grassland of Solapur, India selected sites for den/rendezvous sites close to waterholes (Kumar 1998). Wolves in Dalmatia did not avoid roads; sometimes they even used them for night travels. The density of roads is often used as a measure of habitat quality (Dupré et al. 1995; Mladenoff et al. 1995), but this should be done with caution.
Animal activity measured by fluctuations in signal strength can be overestimated when compared to activity achieved from activity sensor or movement data (Theuerkauf and Jędrzejewski 2002). We used fluctuations in signal strength to determine wolf activity. Even if this method overestimated overall wolf activity, it still allowed us to compare the relative activity levels for different periods. Mech (1992) found that wolves in Minnesota are active at daytime during winter, while Ballard et al. (1991a, b) describe wolves being active at night in Alaska during summer. Theuerkauf et al. (2003a) found that wolves in Białowieża were active throughout the day, with peaks at dusk and dawn, what was related to wolf hunting activity. In those areas, wolves feed on wild ungulates and do not need to come close to people. Wolves in Dalmatia traveled and searched for food mostly at night. Nightly wolf activity was complementary to human activity, allowing the wolves to travel and visit places intensively used by people. We observed wolves to travel unpaved and paved roads, to walk among houses in villages at night, and to visit dumps and water holes close to villages. Vilà et al. (1995) found that wolves in Spain developed nocturnal activity with a tendency to bimodality and Ciucci et al. (1997) found the same pattern for wolves in central Italy. We believe that wolves in Dalmatia follow the same nocturnal pattern, which might have evolved to minimize contact with people. Higher wolf activity at night during summer (versus night activity during winter) can be explained with high-day temperatures during summer. On the other hand, wolf attack on livestock is most frequent during summer (Kusak 2002). This might be the result of two factors:higher need for food because of growing pups, and higher availability of livestock on open pasture during summer. Wolves had to travel from retreat areas, where they were spending days to places closer to people, where they were looking for food. Most of the activity was traveling, which explains the relatively high correlation between activity index and distance traveled. Activity other than traveling was documented too, but in most cases, we could not interpret it in the absence of visual observation. This activity could include feeding on a kill or social activity around the den/rendezvous site, as observed by Mech (1988).
In conclusion, we can state that seasonal variations in home range, habitat use and activity of wolves in Dalmatia reflected a trade-off between avoidance of and use of human-related food sources requiring closer proximity to humans. Seasonal differences in home-range use and daily activity were influenced by human activities—hunting and sheep grazing. Nightly wolf activity was complementary to human activity, allowing the wolves to travel and visit places intensively used by people.
References
Anonymous (1995) Regulation about the protection of certain mammal species (Mammalia). Narodne Novine 31:968
ArcView GIS 3.1 (1996) ArcView GIS 3.1. ESRI. Environmental Systems Research Institute Inc., CA
Ballard WB, Ayes LA, Gardner CL, Foster JW (1991a) Den site activity patterns of gray wolves, Canis lupus, in Southcentral Alaska. Can Field Nat 105:497–504
Ballard WB, Ayres LA, Roney KE, Spraker TH (1991b) Immobilization of gray wolves with a mixture of tiletamine hydrochloride and zolazepam hydrochloride. J Wildl Manage 55:71–74
Bath A, Majić A (2000) Human dimension in wolf management in Croatia. Project Report, 173pp
Blanco JC (1993) The wolf in Spain: problems of conservation in a densely populated country. In: Villa C, Castroviejo J (eds) Simposio Internacionale Sobre el Lobo. Junta de Castilla y León, pp 34–36
Borovac I (2002) The great atlas of Croatia. Mozaik knjiga, Zagreb, 480pp
Božičević S (1995) Position, climate, water and specific land forms. In: Zdunić D (ed) The natural heritage of Croatia. Buvina Ltd, Zagreb, pp 17–28
Ciucci P, Boitani L, Francisci F, Andreoli G (1997) Home range, activity and movements of a wolf pack in central Italy. J Zool Lond 243:803–819
Dupré E, Corsi F, Boitani L, (1995) Potential distribution of the wolf in Italy: a multivariate based GIS model. In: Conference on European wolf migration. Neuchatel, Schwitzerland, pp 1–12
Fritts SH, Mech LD (1981) Dynamics, movements, and feeding ecology of a newly protected wolf population in Northwestern Minnesota. Wildl Monogr 80:1–79
Frković A, Huber D (1992) Wolves in Croatia: baseline data. In: Promberger C, Schröder W (eds) Wolves in Europe—status and perspectives. Munich Wildl Soc Ettal, pp 67–69
Frković A, Ruff R, Cicnjak L, Huber D (1992) Wolf mortality during 1946–86 in Gorski kotar of Croatia, Yugoslavia. In: Bobek B, Perzanowski K, Regelin WL (eds) Global trends in wildlife management. Proceedings of the 18th Congress International Union Game Biology, pp 353–358
Fuller TK (1991) Effect of snow depth on wolf activity and prey selection in North central Minnesota. Can J Zool 69:283–287
Fuller TK (1989) Population dynamics of wolves in North-central Minnesota. Wildl Monogr 105:1–41
Gipson PS, Warren BB, Ronald MN, Mech LD (2000) Accuracy and precision of estimating age of gray wolves by tooth wear. J Wildl Manage 64:752–758
Harrington FH, Mech LD (1982) An analysis of howling response parameters useful for wolf pack censusing. J Wildl Manage 46:686–693
Hooge PN, Eichenlaub W, Solomon E (1999) The animal movement program. USGS, Alaska Biological Science Center
Huber Đ, Kusak J, Frković A, Gužvica G, Gomerčić T (2002) Causes of wolf mortality in Croatia in the period 1986–2001. Veterinarski Arh 72:131–139
Huber D, Kusak J, Kovačić D, Frković A, Radović J, Štahan Ž (1999) Temporary wolf management plan for Croatia. Državna uprava za zaštitu prirode i okoliša 1–76
Jędrzejewski W, Schmidt K, Theuerkauf J, Jędrzejewska B, Okarma H (2002) Daily movements and territory use by radio-collared wolves (Canis lupus) in Białowieża primeval forest in Poland. Can J Zool 79:1993–2004
Kolenosky B, Johnston DH (1967) Radio-tracking timber wolves in Ontario. Am Zool 7:289–303
Korenčić M (1979) Settlements and population of SR Croatia 1857–1971. Type and altitude of settlements. Djela Jugoslvenske akademije znanosti i umjetnosti, Knjiga 54. Republički zavod za statistiku SR Hrvatske, Zagreb, pp 1–900
Kryštufek B, Tvrtković N (1990) Range expansion by Dalmatian jackal population in the 20th century (Canis aureus Linnaeus, 1758). Folia Zool 39:291–296
Kumar S (1998) Ecology and behavior of Indian grey wolf (Canis lupus pallipes Sykes, 1831) in the Deccan grassland of Solapur, Maharashtra, India. PhD Thesis, Center of Wildlife and Ornithology Alighar Muslim University, Alighar, India, pp 1–215
Kusak J (2002) Conditions for life of wolves (Canis lupus L.) in Croatia. PhD Thesis, Biology, Faculty of Science, University of Zagreb, pp 1–247
Lenth RV (1981) On finding the source of a signal. Tehnometrics 23:149–154
Mech LD (1970) The wolf. The ecology and behavior of an endangered species. University of Minnesota press, Minneapolis, MN, pp 1–384
Mech LD (1992) Daytime activity of wolves during winter in Northeastern Minnesota. J Mamm 73:570–571
Mech LD (1994) Regular and homeward travel speeds of arctic wolves. J Mamm 75:741–742
Mech LD, Meier TJ, Burch JW, Adams LG (1995) Patterns of prey selection by wolves in Denaly national park, Alaska. In: Carbyn LN, Frits SH, Seip DR (eds) Ecology and conservation of wolves in a changing world. Canadian Circumpolar Institute, Occasional Publication No. 35, pp 231–243
Mech LD, Adams LG, Meier TJ, Burch JW, Dale BW (1998) The wolves of Denali. University of Minnesota Press, Minneapolis, MN, pp 1–227
Mech LD (1988) The Arctic wolf: ten years with the pack. Voyager Press Inc., Minnesota, MN, pp 1–144
Messier F (1985) Solitary living and extra-territorial movements of wolves in relation to social status and prey abundance. Can J Zool 63:239–245
Mladenoff DJ, Sickley TA, Haight RA, Wydeven AP (1995) A regional landscape analysis and prediction of favorable gray wolf habitat in the Northern great lakes region. Conserv Biol 9:279–294
Nams VO (1993) Locate II. Truro, Nova Scotia
Okarma H, Jędrzejewski W, Schmidt K, Śnieżko S, Bunevich AN, Jędrzejewska B (1998) Home ranges of wolves in Białowieża primeval forest, Poland, compared with other Eurasian populations. J Mamm 79:842–852
Pavlović D (2003) Food habits analysis of wolf (Canis lupus L.) in Croatia. Student Scientific Paper, Veterinary Faculty, University of Zagreb, pp 1–24
Potvin F (1988) Wolf movements and population dynamics in Papineau-Labelle reserve, Quebec. Can J Zool 66:1266–1273
Promberger C, Schröder W (1993) Wolves in Europe—status and perspectives. Munich Wildl Soc Ettal1–136
Promberger C (1996) Carpathian wolf project. Annual report. Munich Wildl Soc Ettal V1: 1–34
Rauš Đ, Trinajstić I, Vukelić J, Medvedović J (1992) Flora of Croatian forests. In: Rauš Đ (ed) Forests in Croatia Šumarski fakultet Sveučilišta u Zagrebu and JP “Hrvatske šume”, Zagreb, pp 33–77
Roth HU, Huber D (1986) Diel activity of brown bears in Plitvice Lakes National Park, Yugouslavia. In: Zager P (ed) International conference on bear research and management, vol. 6, International Bear Association, pp 177–181
Seaman DE, Millspaugh JJ, Kernohan BJ, Brundige GC, Raedeke KJ, Gitzen RA (1999) Effects of sample size on kernel home range estimates. J Wildl Manage 63:739–747
Seletković Z, Katušin S (1992) Climate of Croatia. In: Rauš Đ (ed) Forests in Croatia. Šumarski fakultet Sveučilišta u Zagrebu and “Hrvatske šume”, Zagreb, pp 13–18
Theuerkauf J, Jędrzejewski W (2002) Accuracy of radiotelemetry to estimate wolf activity and locations. J Wildl Manage 66:859–864
Theuerkauf J, Jędrzejewski W, Schmidt K, Okarma H, Ruczynski I, Śnieżko S, Gula R (2003a) Daily patterns and duration of wolf activity in the Białowieża forest, Poland. J Mamm 84:243–253
Theuerkauf J, Jędrzejewski W, Schmidt K, Gula R (2003b) Spatiotemporal segregation of wolves from humans in the Białowieża forest (Poland). J Wildl Manage 67:706–716
Thiel RP, Merrill S, Mech LD (1998) Tolerance by denning wolves, Canis lupus, to human disturbance. Can Field Nat 112:340–342
Vilà C, Urios V, Castrovijeo J (1990) Ecologia del Lobo en la Cabrera (Leon) y la Carballeda (Zamora). In: Blanco HC, Cuesta L, Reig S (eds) El Lobo (Canis lupus) en Espana: Situacion, problematica y apuntes sobre su ecologia. Instituto para la Conervacion de la Naturaleza, Madrid, Spain, pp 95–108
Vilà C, Urios V, Castrovijeo J (1995) Observations on the daily activity patterns in the iberian wolf. In: Carbyn LN, Frits SH, Seip DR (eds) Ecology and conservation of wolves in a changing world. Canadian Circumpolar Institute, Occasional Publication No. 35, pp 335–340
White GC, Garrot RA (1990) Analysis of wildlife radio-tracking data. Academic Press, New York, pp 1–383
Worton BJ (1989) Kernel methods for estimating the utilization in home-ranges studies. Ecology 70:164–168
Acknowledgements
This project was financed by the Croatian Ministry of Science and Bernd Thies Foundation (Switzerland). Permission to study the protected species was given by the Ministry of Environmental Protection and Zoning. Inspectors for damages on livestock provided us valuable information and logistic help at the beginning and during field work. Special thanks go to 26 differing students, whose participation and help in field work was irreplaceable during long days and nights of radiotracking.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kusak, J., Skrbinšek, A.M. & Huber, D. Home ranges, movements, and activity of wolves (Canis lupus) in the Dalmatian part of Dinarids, Croatia. Eur J Wildl Res 51, 254–262 (2005). https://doi.org/10.1007/s10344-005-0111-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-005-0111-2