
Abstract
Neurocognitive impairment in response to methamphetamine (MA) has been proven in a variety of experimental and clinical studies. Elucidation of the underlying mechanisms of MA-induced cognitive deficits and finding preventive/therapeutic approaches need best-suited animal models. In modeling repeated MA exposure, while some believes that escalating doses simulate drug abuse conditions, others believe this regimen confers a preconditioning protection. The present study aimed to compare the effects of three different regimens of repeated MA administration on memory and cognitive function of adult rats. Rats in two different experimental groups were treated with escalating paradigms consisted of twice-daily i.p. injections; 1–4 mg/kg over 7 days or 1–10 mg/kg over 10 days. The third group received twice-daily doses of 15 mg/kg every other day over 14 days. Spatial working memory, novel object recognition task and anxiety-like behavior were measured sequentially in all MA-treated rats and vehicle-treated controls started from day 8 after last injection. All MA regimens decreased rates of spontaneous alternation in Y-maze and increased anxiety-like response. Short-term recognition memory was unchanged across all MA-treated animals, while long-term memory was impaired in the second and third MA regimen. Though MA deleterious effect especially in recognition memory is somehow dose dependent, preconditioning effect of increasing doses may be ruled out at least in the case of parameters measured here.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Background
Methamphetamine (MA) as a psycho-stimulant drug causes neurocognitive and emotional deficits. Cognitive performance such as episodic memory, executive function and information processing undergos substantial impairment upon MA abuse (Cadet and Bisagno 2016; Bernheim et al. 2016; Zhong et al. 2016). Persistent memory loss in chronic MA users (Rusyniak 2013) manifested in a variety of cognitive function tests, such as recall for both words and pictures (Simon et al. 2000; Woods et al. 2005), decision making (Paulus et al. 2002), task performance (London et al. 2005), verbal encoding, and retrieval and episodic memory (Woods et al. 2005; Gonzalez et al. 2007; Hoffman et al. 2006; Simon et al. 2004), highlights the importance of studies focused on the underlying mechanisms and approaches to encounter the long-term effects of MA. Undoubtedly, animal models are important contributors to achieve theses purposes.
According to clinical and experimental data, MA behavioral and psychological consequences depend vastly on various patterns of abuse (Davidson et al. 2005). A variety of experimental animal models have been developed in attempts to mimic the different patterns of MA consumption and to evaluate its neurobehavioral effects on the brain. Many studies have been carried out on MA-induced neurotoxicity. These studies have used patterns of exposure that include the single-day-single-dose administration or the single-day-multiple-dose administration (Marshall et al. 2007; Grace et al. 2010; Ghazvini et al. 2016). These acute toxic dosing models in animals are similar to the acute overdose in humans which may be lethal. Another model of administration in rodent is self-administration which more resembles to drug abuse in humans (Bernheim et al. 2016). However, the most relevant models to clinical MA abuse are defined by initially taking small doses of the drug followed by the use of progressively larger doses (Madden et al. 2005). In modeling such pattern of drug abuse in animals, some researchers believe that exposure to nontoxic amounts of MA will protect against toxic effects of later larger doses, referred to as MA preconditioning (El Ayadi and Zigmond 2011; Hodges et al. 2011; Cadet et al. 2011).
With this background, we selected three different regimens of MA exposure including two multiple increasing-doses regimens and one intermittent toxic regimen to compare the effect of these different exposure patterns on rat performance in novel object recognition, Y-maze and elevated plus maze tests.
Materials and methods
Animals
Adult male Wistar rats, weighing 220–270 g, were obtained from our breeding colony (Neuroscience Research Center), housed with food and water ad libitum, under a standard 12-h light/12-h dark cycle (lights on 7:00–19:00 h), temperature of 23 ± 2 °C and a relative humidity of 50 ± 5%. All the experiments followed the “Principles of Laboratory Animal Care” (NIH Publication No. 85-23, revised 1996) and were approved by the ethics committee for animal research of the Shahid Beheshti University of Medical Sciences.
Methamphetamine administration
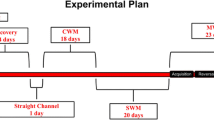
Methamphetamine (MA) hydrochloride (synthesized and analyzed by laboratory of Medicinal Chemistry, School of Pharmacy, Tehran University of Medical Sciences, Iran) was freshly dissolved in normal saline prior to intraperitoneal (i.p.) injections. Rats in three groups were treated with MA at three different regimens. The first group, (MA (1–4), n = 16), received escalating doses of MA: 1‚ 1.5‚ 2‚ 2.5‚ 3‚ 3.5 and 4 mg/kg, twice a day, every day over 7 days. The second group, (MA (1–10), n = 16), received escalating doses of MA: 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 mg/kg, twice a day, every day over 10 days. The third group, (MA (15–15), n = 14), received constant doses of MA: 15 mg/kg, twice a day, every other day over 14 days. MA injections were performed at 4-h intervals between 10 am and 15 pm. After MA injections, rats were kept individually in plastic home-cages with wood-chips bedding for a duration of 2 h after lower doses of MA (≤ 3 mg/kg) and 4 h after higher doses of MA (> 3 mg/kg) and thereafter returned to their shared home-cages. Three control groups, consisting of same animal numbers as MA groups, were also assigned, treated with normal saline (1 ml/kg) at the same time schedules as the MA-treated groups. In MA (15–15) group, two rats died during the course of drug administration and one died at the day 24 of experiment. Body weights were randomly measured for 7 rats per group on the first day of experiments, the day after last MA or saline injections and 1 week later before the beginning of behavioral tests. Schematic illustration summarizing MA treatment regimens and later behavioral tests is provided in Fig. 1.

Experimental protocol. Timelines for the administrations of MA or vehicle in a MA (1–4), b MA (1–10), and c MA (15–15) groups and behavioral tests including: spontaneous alternation behavior; SAB, novel object recognition; NOR and elevated plus maze; EPM
Spontaneous alternation behavior (SAB) test
One week after last MA injections, rats were subjected to the test of Y-maze spontaneous alternation behavior (SAB) as a measure of the spatial working memory (Hughes 2004). The Plexiglas Y-maze consisted of three arms (61 × 14 × 35 cm3), labeled A, B, C, and positioned at an equal angle. Rats were placed at the end of one arm and allowed to explore freely through the maze for 8 min. The sequences of arms entries, as defined as the entry of four paws into one arm, were recorded manually (i.e., ABCABACACBCACAC, etc.). The parameters measured included total number of arm entries, spontaneous alternation performance (SAP; defined as consecutive entries into the three different arms), same arm returns (SAR), and alternate arm returns (AAR). The percent of SAP was calculated as the ratio of absolute number of spontaneous alternations to possible number of alternations (= total arm entries − 2) × 100.
Novel object recognition (NOR) test
The day after Y-maze test, animals were subjected to a novel object recognition (NOR) task, for 4 days. The test is based on the tendency of rat to interact more with a novel object within a familiar environment and therefore assesses rats’ ability to distinguish between novel and familiar objects considered as recognition memory (Ennaceur 2010; Antunes and Biala 2012). NOR test was accomplished in a wooden open field box (40 cm × 40 cm × 38 cm high). All animals were habituated to open field arena by allowing to individually explore the empty arena for 10 min, at the first 2 days of experiment. Twenty-four hours after second habituation phase, the familiarization phase was performed by exposing the animals to the familiar arena with two identical objects (object A1 and object A2) placed in two adjacent corners, for 3 min. Ninety min after familiarization phase, short-term memory (STM) was tested by replacing one of familiar objects with one novel object (B). On the day four of experiment, long-term memory (LTM) was tested by replacing the object (B) with another novel one (C). During the STM and LTM phases, each rat was allowed to explore the objects for 3 min. All the objects were plastic Lego, consistent in height and volume, but different in shape and appearance. Objects were selected after a preliminary experiment with a distinct group of animals to be sure that all the objects are equally preferred at the first encounter.
All trials were videotaped, and an experimenter blind to the treatment conditions reported the total exploration time for each object. Exploration was considered as sniffing or touching the objects with the nose and sitting on the object was not considered as exploration. Rats that spent a total of less than 12 s exploring two objects during the familiarization phase were excluded from the experiment. The novel object preference index percentage was calculated as the ratio of time spent exploring new objects in STM or LTM tests (B or C) over the total time spent exploring both objects, i.e., (A + B) or (A + C) multiplied 100.
Elevated plus maze (EPM) test
The day after NOR test completed, rats were subjected to EPM test, as a widely used behavioral test to assess the anxiety-related behavior in rodents (Walf and Frye 2007). The Plexiglas plus maze apparatus was consisted of a plus-shaped platform with two open (50 × 10 cm) and two closed arms (50 × 10 × 40 cm). The whole apparatus was set 50 cm above the test room floor. Rats were individually placed on the center of the platform with the head facing one of the open arms and allowed to freely explore the maze for a testing period of 5 min. The number of entries into the open and closed arms and the total time spent in the open and closed arms were recorded. An arm entry was defined when all four paws of the rat were in the arm. Time spent in the central area was not included in either open arm time or closed arm time. The open arms entries percentage (OAE% = the ratio of entries into open arms to the total entries × 100) and the percentage of time spent in open arms [OAT% = the ratio of times spent in the open arms to total time spent in any arms × 100) were reported as the criteria of open space-induced anxiety-like behavior.
Statistical analysis
An overall two-way analysis of variance (ANOVA) was used to evaluate the effect of MA and different regimens of MA exposure over the control groups on the behavioral parameters. The Student’s t test was applied to compare each MA group versus the corresponding control values. In the familiarization phase of NOR test, a two-tailed paired t test was employed to compare time spent exploring the identical objects. A two-way repeated measures analysis of variance (ANOVA) was also used to compare the means of repeated body weight measurements. Data are presented as mean ± SEM, and all statistical significant differences are defined at p < 0.05 using SPSS v. 21.
Results
In this study, the effect of three different regiments of MA including different doses over different time durations were studied on the rats’ spatial working memory, recognition memory and anxiety-related behavior. Regiments were consisted of 36 mg/kg of MA over 7 days in group MA (1–4), 110 mg/kg over 10 days in group MA (1–10) and 210 mg/kg over 14 days in MA (15–15) group. Inhibitory effects of MA on food intake and body weight changes could be confounding variables. However, it has been shown that MA discontinuation is accompanied with a dramatically increase in food intake (Saito et al. 1995). Therefore, in all groups, behavioral tests were started after a 1 week free of drug to reduce the effect of immediate behavioral responses to MA on the tests parameters and therefore better focus on the probable long-lasting effects of MA on cognition. Body weight data obtained at the first day of experiments, a day after last MA or saline injections and before the beginning of behavioral tests were analyzed with two-way ANOVA for repeated measures. Results revealed that the main effect of time was significant (p < 0.001); however, the main effect of MA treatment was not significant (p > 0.05) and therefore there was no significant difference between body weights in MA-treated groups and corresponding controls at the beginning of behavioral tests (Fig. 2).

Body weight of rats treated with three different regimens of MA and the corresponding controls. Body weight was measured at the first day of experiment, the day after last MA or saline injection, and 1 week later, before the behavioral tests in a MA (1–4), b MA (1–10), and c MA (15–15) groups. n = 7 per group
Spatial working memory
Y-maze data were first analyzed by an overall multivariate two-way ANOVA, with MA treatment and regimens as the two independent variables. The results showed a significant overall effect of MA treatment on SAP (F1,84 = 23.044, p < 0.001), SAR (F1,84 = 13.516, p < 0.001) and SAP % (F1,84 = 42.586, p < 0.001). This is while MA treatment had no significant effect on the total number of arm entries as a measure of general locomotor activity (F1,84 = 1.975, p > 0.05) and also on AAR (F1,84 = 3.840, p > 0.05). The main effect of MA regimen was not significant at all measured parameters (p > 0.05). Comparing each MA regimen with the corresponding control revealed that MA treatment in (1–4) and (15–15) groups significantly decreased the SAP number (p < 0.01, t test) and SAP % (p < 0.001, t test) and in parallel increased the SAR number (p < 0.05, t test). This working memory impairment manifested by the decrease in SAP and SAP % and increase in SAR and AAR was also observed in MA (1–10) group although it was statistically significant only for the SAP % parameter (p < 0.01, t test) (Fig. 3).

Effects of three different regimens of MA on spontaneous alternation behavior in Y-maze. The total number of arm entries (a), number of spontaneous alternation performance (SAP) (b), number of same arm returns (SAR) (c), alternate arm returns (AAR) (d) and the percentage of SAP (e) are presented as mean ± SEM. *p < 0.05, **p < 0.01 and ***p < 0.001 versus corresponding controls (Ctrl) (t test). n = 12–16 per group
Novel object recognition memory
In familiarization phase of novel object recognition task, paired t test analysis showed that there was no significant difference in the absolute time spent exploring each individual object in each experimental group (Fig. 4a), suggesting that there was not a place bias during learning. In assessing short-term recognition memory, 90 min after the familiarization phase, an overall two-way ANOVA showed that MA treatment has a significant main effect on reducing the novel object preference index (F1,66 = 6.930, p < 0.05) although the main effect of MA regimen was not significant (F2,66 = 2.202, p > 0.05). Further independent t test analysis to compare each MA-treated group with the corresponding control revealed that although the novel object preference is reduced in MA exposed rats, it does not reach statistically significant (Fig. 4b).

Effects of three different regimens of MA on novel object recognition memory. Total time spent exploring identical objects during the familiarization phase (a), and the preferences indices for the novel objects introduced 90 min (short-term memory; STM) (b) and 24 h (long-term memory; LTM) (c) after familiarization phase are presented as mean ± SEM. *p < 0.05 and ***p < 0.001 versus corresponding controls (t test). n = 9–15 per group
Analysis of the preference index for another novel object introduced 24 h after familiarization, as a measure of long-term recognition memory, showed a significant main effect of MA treatment (F1,66 = 22.282, p < 0.001) and also a significant effect of MA regimen (F2,66 = 6.305, p < 0.01). Further t test analysis indicated that MA-treated rats exhibit recognition memory deficits manifested by significant decrease in novel object preference index in MA (1–10) and MA (15–15) groups (p < 0.001 and p < 0.05, respectively) (Fig. 4c).
Anxiety-related behavior
The EPM test was used to assess anxiety in rats following MA-induced spatial working memory and recognition memory impairments. Two-way ANOVA revealed an overall significant effect of MA treatment on OAE% (F1,53 = 36.492, p < 0.001) and OAT% (F1,53 = 53.530, p < 0.001) parameters. The main effect of MA regimens was not significant on OAE% (F2,53 = 0.466, p > 0.05) but was significant on OAT% (F2,53 = 4.204, p < 0.05). Further t test analysis of OAE% and OAT% in MA-treated groups, each with its own control also showed the significant increase of anxiety-related behavior manifested as fewer entries into and lest time spent in open arms of plus maze in all MA-treated groups (Fig. 5a, b).

Effects of three different regimens of MA on anxiety-related behavior. The percent of open arm entries, OAE% (a) and open arm time, OAT% (b) are indices of anxiety-related behavior. *p < 0.05, **p < 0.01 and ***p < 0.001 versus corresponding controls (t test). Data are presented as mean ± SEM, n = 9–10 per group
Discussion
The present study revealed that MA exposure at three different regimens varying in average daily doses, total doses and exposure duration, impairs rats’ cognitive function after 1 week of drug-free period. The impaired cognitive function can be attributed to several neurotoxic mechanisms depending on the drug dose and the exposure time. Besides acute increases in dopamine and serotonin levels following lower doses of MA, the hyperthermia produced by high doses of MA is also a cause of nerve terminal damages, breakdowns in the blood–brain barrier, neuroinflammation and neurodegeneration (Bowyer and Hanig 2014; Yu et al. 2015). However, giving a 1-week drug-free period in the current study, we aimed to focus on the long-lasting cognitive effects of MA regardless of direct or indirect underlying mechanisms.
Spontaneous alternation performance (SAP) is considered as a measure of spatial working memory or short-term memory based on the innate tendency of rats to enter a prior unexplored arm of Y-maze with the advantage of avoiding stressful handling of animals and also providing useful measure of locomotor activity (Hughes 2004; Bizon et al. 2012). Although spontaneous alternation is considered as a hippocampal-dependent task, lesions of other structures of the brain are also shown to impair it (Ainge et al. 2007; Bizon et al. 2012). Previous studies show that though acute low doses of MA (2 mg/kg) do not induce memory impairment (Lee et al. 2011), acute large doses (30 mg/kg) impair rat’s spatial memory in Y-maze a day after drug exposure (Simões et al. 2007). Regarding the effect of repeated MA exposure on spatial memory, there are lots of controversial data obtained from experiments using different MA doses and regimens in mice or rats, measuring memory at different time points in the course of MA exposure or following a period of drug-free. For instance, it is shown that sensitization to MA following repeated low doses (2 mg/kg) is accompanied by reduced SAP and decreased NMDA receptor expression in mice prefrontal cortex and hippocampus (Lee et al. 2011). Adolescent male mice exposed to repeated neurotoxic doses of MA (24 mg/kg/daily for 14 days) though is shown to exhibit normal short-term memory performance during active drug exposure and exhibit impaired hippocampal synaptic plasticity and short-term memory after days of drug abstinence (North et al. 2013). Unexpectedly, in another study, it is revealed that adolescent mice treated with two weekly injections of MA (30 mg/kg) exhibit an improved performance in radial arm maze working memory at 1 week following the first MA injection. This is while these two MA doses result in impaired working memory performance across 5 weeks of study after drug abstinence, attributed to alteration of the expression of glutamate and dopamine signaling elements in the hippocampus and dorsal striatum (Braren et al. 2014). In a rat model mimicking, the common human “abuser” condition, which is characterized by compulsive consumption of growing doses, it is shown that repeated MA (10, 15, 15, 20, 20, 25 and 30 mg/kg/day) exposure over 7 days does not cause a mnemonic deficit in Y-maze test suggesting a kind of precondition/protection provided by early exposure to lower doses (Simões et al. 2007). However, Simões et al. only measures spatial working memory on day 7 and such kind of protection cannot be expanded to the time after drug abstinence. However, it is repeatedly reported that working memory deficits is one of the main neurocognitive consequences of MA abuse in humans which may be even followed by years of abstinence (Hart et al. 2012; Rusyniak 2013). Consistently, assessing three different regimens of MA in rats we showed that low and high multiple increasing-doses regimens as well as intermittent toxic doses regimen all result in a significant spatial working memory impairment following 1 week drug abstinence.
To further compare the regimens selected for modeling repeated MA exposure in rats, object recognition task was used to test episodic memory (Ennaceur 2010; Dickerson and Eichenbaum 2010) which is particularly compromised in MA abusers with deficits observed in multiple aspects of recognition memory (Rusyniak 2013). Memory for object novelty recognition is primarily dependent on the perirhinal cortex and interactions with the hippocampus (Aggleton et al. 2010; Antunes and Biala 2012). In the present study, MA exposure had no effect on rat’s preference to one of two identical objects in familiarization phase which is similar with previous data confirming no effect of MA on initial exploratory behavior of rats in NOR test (Reichel et al. 2011). In recognizing novel object shortly after familiarization, though there were signs of memory deficit manifested by no significant difference with the preference indexes in the familiarization phase, there were also no significant differences with the corresponding controls. Consistently, it has been shown that multiple escalating doses of MA in a course of 14 days do not impair recognition memory in rats assessed 3 h post-familiarization on day 6 of drug abstinence (Clark et al. 2007). However, as mentioned above, searching the effects of repeated MA exposure on recognition memory reveals controversial data in rodents. There are several documents showing that repeated neurotoxic doses, multiple sort-term neurotoxic doses and even sensitizing doses of MA impair the short-term recognition memory in rats and mice (Bisagno et al. 2002; Belcher et al. 2005; He et al. 2006; Marshall et al. 2007; Reichel et al. 2011). The retrieval of short-term recognition memory in the three regimens of MA studied herein shows that MA toxicity in these regimes is not sufficient to account for the impaired recognition memory in short term. It is also not reasonable to suggest a protective role for the adaptations associated with early exposure to lower doses of MA since the preserved short-term memory was recorded not only in two multiple increasing-doses regimens but also in the intermittent toxic doses regimen.
The next finding of the NOR test was that the recognition memory is impaired a day after familiarization to objects in MA (1–10) and MA (15–15) groups. This is in apparently consistent with some previous reports (Kamei et al. 2006; Gonçalves et al. 2012) and highlights the effect of a threshold dose of MA in producing long-lasting recognition memory retrieval deficits.
MA-induced neuronal toxicity and plasticity along with memory impairment leads to some other persistent behavioral changes, such as increased anxiety-related behaviors (Glasner-Edwards et al. 2010; Darke et al. 2008). Both in rodents and humans, anxiety is a common symptom in physical dependence and withdrawal of MA (McGregor et al. 2005; Hajheidari et al. 2015). It has been reportedly shown that both acute single dose and chronic repeated doses of MA cause anxiety-related behavioral symptoms in rodents, at different time points of drug abstinence (Hayase et al. 2005; Pometlová et al. 2012; Šlamberová et al. 2015). There are also some reports showing no effects of escalating doses of MA on anxiety parameters in mice (Kitanaka et al. 2010) or even anxiolytic-like responses in rats (Miladi-Gorji et al. 2015). This discrepancy among different reports might be attributed to the doses used, the route and number of injections, time between injections and testing time points (Kitanaka et al. 2008). Although the precise mechanisms by which MA affects anxiety-related behavior are far from being completely understood (Glasner-Edwards et al. 2010), in simulating a model for the study of long-lasting neuropsychiatric effects of MA, co-occurrence of the anxiety may serve as an index of suitability. The data of the present study showed that MA in all three regiments modeling repeated drug intake increase the avoidance to open arms of elevated plus maze, while the locomotor activity was unchanged.
In summary, MA at total doses of 36, 110 and 210 mg/kg induces spatial working memory impairment along with anxiety-related behavioral responses. MA deleterious effects in novel object recognition memory are somehow dose dependent and finally preconditioning effect of increasing doses may be ruled out at least in the case of parameters measured here. Present result emphasize that the regimen 1–10 mg/kg serves as a good model in rat for the repeated MA exposure inducing spatial working memory and novel object recognition memory impairments, with no mortality compared with repeated toxic doses.
References
Aggleton JP, Albasser MM, Aggleton DJ, Poirier GL, Pearce JM (2010) Lesions of the rat perirhinal cortex spare the acquisition of a complex configural visual discrimination yet impair object recognition. Behav Neurosci 124(1):55–68. doi:10.1037/a0018320
Ainge JA, van der Meer MA, Langston RF, Wood ER (2007) Exploring the role of context-dependent hippocampal activity in spatial alternation behavior. Hippocampus 17(10):988–1002
Antunes M, Biala G (2012) The novel object recognition memory: neurobiology, test procedure, and its modifications. Cogn Process 13(2):93–110. doi:10.1007/s10339-011-0430-z
Belcher AM, O’Dell SJ, Marshall JF (2005) Impaired object recognition memory following methamphetamine, but not p-chloroamphetamine- or d-amphetamine-induced neurotoxicity. Neuropsychopharmacology 30(11):2026–2034
Bernheim A, See RE, Reichel CM (2016) Chronic methamphetamine self-administration disrupts cortical control of cognition. Neurosci Biobehav Rev 69:36–48. doi:10.1016/j.neubiorev.2016.07.020
Bisagno V, Ferguson D, Luine VN (2002) Short toxic methamphetamine schedule impairs object recognition task in male rats. Brain Res 940(1–2):95–101
Bizon JL, Foster TC, Alexander GE, Glisky EL (2012) Characterizing cognitive aging of working memory and executive function in animal models. Front Aging Neurosci 4:19 (eCollection 2012)
Bowyer JF, Hanig JP (2014) Amphetamine- and methamphetamine-induced hyperthermia: implications of the effects produced in brain vasculature and peripheral organs to forebrain neurotoxicity. Temperature (Austin) 1(3):172–182. doi:10.4161/23328940.2014.982049
Braren SH, Drapala D, Tulloch IK, Serrano PA (2014) Methamphetamine-induced short-term increase and long-term decrease in spatial working memory affects protein Kinase M zeta (PKMζ), dopamine, and glutamate receptors. Front Behav Neurosci 8:438. doi:10.3389/fnbeh.2014.00438
Cadet JL, Bisagno V (2016) Neuropsychological consequences of chronic drug use: relevance to treatment approaches. Front Psychiatry 6:189. doi:10.3389/fpsyt.2015.00189 (eCollection 2015)
Cadet JL, Brannock C, Ladenheim B, McCoy MT, Beauvais G, Hodges AB, Lehrmann E, Wood WH 3rd, Becker KG, Krasnova IN (2011) Methamphetamine preconditioning causes differential changes in striatal transcriptional responses to large doses of the drug. Dose Response 9(2):165–181
Clark RE, Kuczenski R, Segal DS (2007) Escalating dose, multiple binge methamphetamine regimen does not impair recognition memory in rats. Synapse 61(7):515–522
Darke S, Kaye S, McKetin R, Duflou J (2008) Major physical and psychological harms of methamphetamine use. Drug Alcohol Rev 27(3):253–262. doi:10.1080/09595230801923702
Davidson C, Lee TH, Ellinwood EH (2005) Acute and chronic continuous methamphetamine have different long-term behavioral and neurochemical consequences. Neurochem Int 46(3):189–203
Dickerson BC, Eichenbaum H (2010) The episodic memory system: neurocircuitry and disorders. Neuropsychopharmacology 35(1):86–104. doi:10.1038/npp.2009.126
El Ayadi A, Zigmond MJ (2011) Low concentrations of methamphetamine can protect dopaminergic cells against a larger oxidative stress injury: mechanistic study. PLoS ONE 6(10):e24722
Ennaceur A (2010) One-trial object recognition in rats and mice: methodological and theoretical issues. Behav Brain Res 215(2):244–254. doi:10.1016/j.bbr.2009.12.036
Ghazvini H, Khaksari M, Esmaeilpour K, Shabani M, Asadi-Shekaari M, Khodamoradi M, Sheibani V (2016) Effects of treatment with estrogen and progesterone on the methamphetamine-induced cognitive impairment in ovariectomized rats. Neurosci Lett 619:60–67. doi:10.1016/j.neulet.2016.02.057
Glasner-Edwards S, Mooney LJ, Marinelli-Casey P, Hillhouse M, Ang A, Rawson R, Methamphetamine Treatment Project Corporate Authors (2010) Anxiety disorders among methamphetamine dependent adults: association with post-treatment functioning. Am J Addict 19(5):385–390. doi:10.1111/j.1521-0391.2010.00061.x
Gonçalves J, Baptista S, Olesen MV, Fontes-Ribeiro C, Malva JO, Woldbye DP, Silva AP (2012) Methamphetamine-induced changes in the mice hippocampal neuropeptide Y system: implications for memory impairment. J Neurochem 123(6):1041–1053. doi:10.1111/jnc.12052
Gonzalez R, Bechara A, Martin EM (2007) Executive functions among individuals with methamphetamine or alcohol as drugs of choice: preliminary observations. J Clin Exp Neuropsychol 29(2):155–159
Grace CE, Schaefer TL, Herring NR, Graham DL, Skelton MR, Gudelsky GA, Williams MT, Vorhees CV (2010) Effect of a neurotoxic dose regimen of (+)-methamphetamine on behavior, plasma corticosterone, and brain monoamines in adult C57BL/6 mice. Neurotoxicol Teratol 32(3):346–355
Hajheidari S, Miladi-Gorji H, Bigdeli I (2015) Effect of the environmental enrichment on the severity of psychological dependence and voluntary methamphetamine consumption in methamphetamine withdrawn rats. Neurosci Lett 584:151–155. doi:10.1016/j.neulet.2014.10.017
Hart CL, Marvin CB, Silver R, Smith EE (2012) Is cognitive functioning impaired in methamphetamine users? A critical review. Neuropsychopharmacology 37(3):586–608
Hayase T, Yamamoto Y, Yamamoto K (2005) Persistent anxiogenic effects of a single or repeated doses of cocaine and methamphetamine: interactions with endogenous cannabinoid receptor ligands. Behav Pharmacol 16(5–6):395–404
He J, Yang Y, Yu Y, Li X, Li XM (2006) The effects of chronic administration of quetiapine on the methamphetamine-induced recognition memory impairment and dopaminergic terminal deficit in rats. Behav Brain Res 172(1):39–45
Hodges AB, Ladenheim B, McCoy MT, Beauvais G, Cai N, Krasnova IN, Cadet JL (2011) Long-term protective effects of methamphetamine preconditioning against single-day methamphetamine toxic challenges. Curr Neuropharmacol 9(1):35–39
Hoffman WF, Moore M, Templin R, McFarland B, Hitzemann RJ, Mitchell SH (2006) Neuropsychological function and delay discounting in methamphetamine-dependent individuals. Psychopharmacology 188(2):162–170
Hughes RN (2004) The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci Biobehav Rev 28(5):497–505
Kamei H, Nagai T, Nakano H, Togan Y, Takayanagi M, Takahashi K, Kobayashi K, Yoshida S, Maeda K, Takuma K, Nabeshima T, Yamada K (2006) Repeated methamphetamine treatment impairs recognition memory through a failure of novelty-induced ERK1/2 activation in the prefrontal cortex of mice. Biol Psychiatry 59(1):75–84
Kitanaka J, Kitanaka N, Takemura M (2008) Neurochemical consequences of dysphoric state during amphetamine withdrawal in animal models: a review. Neurochem Res 33(1):204–219 (Epub 2007 Jun 29)
Kitanaka N, Kitanaka J, Tatsuta T, Tanaka K, Watabe K, Nishiyama N, Morita Y, Takemura M (2010) Withdrawal from fixed-dose injection of methamphetamine decreases cerebral levels of 3-methoxy-4-hydroxyphenylglycol and induces the expression of anxiety-related behavior in mice. Neurochem Res 35(5):749–760. doi:10.1007/s11064-010-0132-4
Lee KW, Kim HC, Lee SY, Jang CG (2011) Methamphetamine-sensitized mice are accompanied by memory impairment and reduction of N-methyl-d-aspartate receptor ligand binding in the prefrontal cortex and hippocampus. Neuroscience 178:101–107. doi:10.1016/j.neuroscience.2011.01.025 (Epub 2011 Jan 19)
London ED, Berman SM, Voytek B, Simon SL, Mandelkern MA, Monterosso J, Thompson PM, Brody AL, Geaga JA, Hong MS, Hayashi KM, Rawson RA, Ling W (2005) Cerebral metabolic dysfunction and impaired vigilance in recently abstinent methamphetamine abusers. Biol Psychiatry 58(10):770–778
Madden LJ, Flynn CT, Zandonatti MA, May M, Parsons LH, Katner SN, Henriksen SJ, Fox HS (2005) Modeling human methamphetamine exposure in nonhuman primates: chronic dosing in the rhesus macaque leads to behavioral and physiological abnormalities. Neuropsychopharmacology 30(2):350–359
Marshall JF, Belcher AM, Feinstein EM, O’Dell SJ (2007) Methamphetamine-induced neural and cognitive changes in rodents. Addiction 102(Suppl 1):61–69
McGregor C, Srisurapanont M, Jittiwutikarn J, Laobhripatr S, Wongtan T, White JM (2005) The nature, time course and severity of methamphetamine withdrawal. Addiction 100(9):1320–1329
Miladi-Gorji H, Fadaei A, Bigdeli I (2015) Anxiety assessment in methamphetamine—sensitized and withdrawn rats: immediate and delayed effects. Iran J Psychiatry 10(3):150–157
North A, Swant J, Salvatore MF, Gamble-George J, Prins P, Butler B, Mittal MK, Heltsley R, Clark JT, Khoshbouei H (2013) Chronic methamphetamine exposure produces a delayed, long-lasting memory deficit. Synapse 67(5):245–257. doi:10.1002/syn.21635
Paulus MP, Hozack NE, Zauscher BE, Frank L, Brown GG, Braff DL, Schuckit MA (2002) Behavioral and functional neuroimaging evidence for prefrontal dysfunction in methamphetamine-dependent subjects. Neuropsychopharmacology 26(1):53–63
Pometlová M, Nohejlová-Deykun K, Slamberová R (2012) Anxiogenic effect of low-dose methamphetamine in the test of elevated plus-maze. Prague Med Rep 113(3):223–230
Reichel CM, Schwendt M, McGinty JF, Olive MF, See RE (2011) Loss of object recognition memory produced by extended access to methamphetamine self-administration is reversed by positive allosteric modulation of metabotropic glutamate receptor 5. Neuropsychopharmacology 36(4):782–792. doi:10.1038/npp.2010.212
Rusyniak DE (2013) Neurologic manifestations of chronic methamphetamine abuse. Psychiatr Clin N Am 36(2):261–275. doi:10.1016/j.psc.2013.02.005
Saito M, Terada M, Saito TR, Takahashi KW (1995) Effects of the long-term administration of methamphetamine on body weight, food intake, blood biochemistry and estrous cycle in rats. Exp Anim 43(5):747–754
Simões PF, Silva AP, Pereira FC, Marques E, Grade S, Milhazes N, Borges F, Ribeiro CF, Macedo TR (2007) Methamphetamine induces alterations on hippocampal NMDA and AMPA receptor subunit levels and impairs spatial working memory. Neuroscience 150(2):433–441
Simon SL, Domier C, Carnell J, Brethen P, Rawson R, Ling W (2000) Cognitive impairment in individuals currently using methamphetamine. Am J Addict 9(3):222–231
Simon SL, Dacey J, Glynn S, Rawson R, Ling W (2004) The effect of relapse on cognition in abstinent methamphetamine abusers. J Subst Abuse Treat 27(1):59–66
Šlamberová R, Pometlová M, Macúchová E, Nohejlová K, Stuchlík A, Valeš K (2015) Do the effects of prenatal exposure and acute treatment of methamphetamine on anxiety vary depending on the animal model used? Behav Brain Res 292:361–369. doi:10.1016/j.bbr.2015.07.001
Walf AA, Frye CA (2007) The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat Protoc 2(2):322–328
Woods SP, Rippeth JD, Conover E, Gongvatana A, Gonzalez R, Carey CL, Cherner M, Heaton RK, Grant I, HIV Neurobehavioral Research Center Group (2005) Deficient strategic control of verbal encoding and retrieval in individuals with methamphetamine dependence. Neuropsychology 19(1):35–43
Yu S, Zhu L, Shen Q, Bai X, Di X (2015) Recent advances in methamphetamine neurotoxicity mechanisms and its molecular pathophysiology. Behav Neurol 2015:103969. doi:10.1155/2015/103969
Zhong N, Jiang H, Du J, Zhao Y, Sun H, Xu D, Li C, Zhuang W, Li X, Hashimoto K, Zhao M (2016) The cognitive impairments and psychological wellbeing of methamphetamine dependent patients compared with health controls. Prog Neuropsychopharmacol Biol Psychiatry 69:31–37. doi:10.1016/j.pnpbp.2016.04.005
Acknowledgements
The authors are thankful to the Neuroscience Research Center of Shahid Beheshti University of Medical Sciences for funding this research.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Handling editor: Maria Gulinello (Albert Einstein College of Medicine, New York).
Reviewers: Charles Vorhees (University of Cincinnati), Jun-Xu Li (University at Buffalo).
Rights and permissions
About this article

Cite this article
Seyedhosseini Tamijani, S.M., Beirami, E., Ahmadiani, A. et al. Effect of three different regimens of repeated methamphetamine on rats’ cognitive performance. Cogn Process 19, 107–115 (2018). https://doi.org/10.1007/s10339-017-0839-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10339-017-0839-0