
Abstract
For irruptive or facultative migrants in the subfamily Carduelinae, the winter finches, seasonal movements are extensive but have been poorly described in North America. We used stable hydrogen isotope measurements of feathers (δ2Hf) collected in three regions in Ontario, Canada, during the winters of 2014–15 and 2015–16 to infer potential breeding origins of four species. We also deployed color bands to better understand migratory movements within and across winters. Values of δ2Hf differed by age, sex, and capture location both within and among species. Predicted breeding origins of wintering evening grosbeaks (Coccothraustes vespertinus) and pine grosbeaks (Pinicola enucleator) were largely from northwestern North America, suggesting a predominately west–east pattern of seasonal movements. Common redpolls (Acanthis flammea) and hoary redpolls (A. hornemanni) largely originated from the High Arctic or western Canada, suggesting predominantly north–south seasonal movements. Grosbeaks were most often recaptured or re-sighted at the same capture location between winters. Recaptures and observations of color-banded redpolls only occurred within a winter and mostly at the same capture location. Our results document the first linkages between potential breeding and wintering sites for a large number of individual winter finches during two successive winters. Observed variation in breeding origins and movement patterns suggests each species employs a different migration strategy; therefore, future research and conservation efforts might be more beneficial if directed at individual species of winter finch.
Zusammenfassung
Gleich und gleich gesellt sich nicht immer gern: unterschiedliche Ursprungsmausergebiete und Zugmuster bei Winterfinken in Ontario
Die sogenannten "Winterfinken", also invasionsartig beziehungsweise fakultativ ziehende Vogelarten aus der Unterfamilie Carduelinae, zeigen ausgedehnte saisonale Zugbewegungen, welche allerdings für Nordamerika kaum erforscht sind. Mittels stabiler Wasserstoff-Isotopendaten (δ2Hf) von Federn, welche während der Winter 2014-15 und 2015-16 in drei Regionen Ontarios (Kanada) gesammelt wurden, versuchten wir, Rückschlüsse auf die potenziellen Ursprungsbrutgebiete von vier Arten zu ziehen. Außerdem setzten wir Farbringe ein, um Zugbewegungen während des Winters und zwischen Wintern besser nachvollziehen zu können. Die δ2Hf-Werte unterschieden sich nach Alter, Geschlecht und Fangort sowohl innerhalb als auch zwischen Arten. Die angenommenen Ursprungsbrutgebiete überwinternder Abendkernbeißer (Coccothraustes vespertinus) und Hakengimpel (Pinicola enucleator) lagen hauptsächlich im nordwestlichen Nordamerika, was eine überwiegende West-Ost-Ausrichtung der saisonalen Zugbewegungen vermuten lässt. Taigabirkenzeisige (Acanthis flammea) und Polarbirkenzeisige (A. hornemanni) stammten überwiegend aus hohen arktischen Breiten oder aus Westkanada, was auf saisonale Bewegungen hauptsächlich in Nord-Süd-Richtung schließen lässt. Bei den Kernbeißern und Gimpeln erfolgten die meisten Wiederfänge oder Wiedersichtungen in verschiedenen Wintern am selben Ort. Wiederfänge und Sichtungen von farbberingten Birkenzeisigen traten nur innerhalb eines Winters und zumeist am selben Fangort auf. Unsere Ergebnisse dokumentieren die ersten Zusammenhänge zwischen potenziellen Brut- und Überwinterungsgebieten für eine große Anzahl individueller Winterfinken während zweier aufeinanderfolgender Winter. Die festgestellte Variation bezüglich der Ursprungsbrutgebiete und der Zugmuster legt nahe, dass eine jede Art eine andere Zugstrategie anwendet; daher könnte es für zukünftige Untersuchungen und Schutzmaßnahmen sinnvoll sein, diese individuell auf einzelne Winterfinkenarten auszurichten.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
For most migratory species, seasonal movements typically involve consistent and predictable timing and routes of return migration from a breeding site to another location where conditions are more favorable for survival (Berthold 2001; Newton 2008, 2012). In other species, migratory movements are irruptive and less predictable in time and space (Newton 2008). In general, irruptive migrants exhibit highly variable migratory behaviors within individuals, between sexes, and within and among populations, often resulting in drastic differences in timing, orientation, and distances of movements within and across seasons and years (Prescott 1991; Newton 2012; Fudikar et al. 2013).
In North America, northern-breeding seed-eating birds in the subfamily Carduelinae, referred to as winter finches, show spectacular periodic irruptions between late fall and early spring, presumably in response to the variable nature of tree and shrub mast abundance (Hochachka et al. 1999; Koenig 2001; Koenig and Knops 2001; Strong et al. 2015). Winter finches breed throughout the boreal forest, Subarctic, and most Arctic regions. They leave their breeding grounds following severe seed crop declines (Bock and Lepthien 1976), in response to high bird population densities following years of banner seed crops (Widrlechner and Dragula 1984), or both (Koenig and Knops 2001). Evidence from band-recapture data suggests that several species of winter finch in North America may undertake longitudinal migrations, often covering thousands of kilometers between breeding and wintering sites (Troy 1983; Brewer et al. 2000; Strong et al. 2015).
Evening grosbeaks and Pine grosbeaks forage on a variety of large seeds and berries during winter (Adkisson 1999; Gillihan and Byers 2001). In contrast, common redpolls and Hoary redpolls forage on small seeds and grasses during winter (Knox and Lowther 2000a, b), suggesting each species group tracks and responds to variation in different food resources. All four species are sexually dimorphic, based on plumage, linear measurements (e.g., wing chord), and mass with females generally 3–5% smaller than males (Knox and Lowther 2000a, b; Gillihan and Byers 2001). These morphological and dietary differences could result in some population structuring during winter. Previous work on winter finches suggested differential migration between sexes resulted in latitudinal variation of sex ratios during winter (Prescott 1991, 1994), with larger-bodied males wintering further north than females. Populations may also be spatially structured during winter if migratory distances are limited by morphological characteristics (e.g., wing length; Winkler and Leisler 1992).
Several species of winter finches are purportedly declining (Bonter and Harvey 2008; COSEWIC 2016). Determining potential causes and subsequently applying conservation actions are complicated by their irruptive and generally nomadic movements. Although continental-scale winter surveys (e.g., christmas bird count, project feeder watch) may be sufficient for assessing general trends of these species (Lepage and Francis 2002), lack of knowledge of factors influencing trends at discrete spatial units, particularly on the breeding grounds, reduces the utility of these trends for conservation and management (Ralston et al. 2015). Current monitoring programs (e.g., Breeding Bird Survey) do not cover adequate portions of the breeding range of some species in the boreal forest and cover only minimal parts of breeding range of the Arctic-breeding species, mostly in Alaska (Dunn 2005). Development of monitoring programs for these regions is difficult given the lack of roads and low human density (Roy et al. 2019).
Recent technological advances, largely in the development and novel use of extrinsic markers, such as miniaturized radio transmitters and data logging devices (e.g., automated radio telemetry, archival light-level geolocators) have vastly improved our understanding of movement in many bird species (Rubenstein and Hobson 2004; Stutchbury et al. 2009; Taylor et al. 2017). Despite these advances, such approaches have limited application for species that exhibit irregular or unpredictable movements, especially if tagged birds must be recaptured to recover logged data, or if birds have to pass through specific areas to be detected (e.g., coded VHF tags). Likewise, band-recapture data have provided valuable information on distances and directions of movements and breeding site fidelity of some irruptive species; however, such information typically must be obtained over many years and there are obvious limitations when using those data for understanding connectivity (e.g., spatial and temporal bias, low band recaptures; Prescott 1991; Brewer et al. 2000).
The use of natural, intrinsic biological or biogeochemical markers, such as naturally occurring stable isotopes of key elements are well suited to the study of irruptive species because only one capture is required. The measurement of stable hydrogen isotope ratios in bird feathers (δ2Hf) has proven particularly effective as a means of inferring molt or natal origin (Hobson and Wassenaar 2019). This is due to the fact that hydrogen in feathers is ultimately derived from precipitation which shows continent-wide amount-weighted pattern in δ2H (i.e., isoscapes; Bowen et al. 2005). Once formed, feathers and other metabolically inert tissues ‘lock in’ these isotopic signals enabling estimates of likely regions of origin using probabilistic approaches (Hobson et al. 2014). This method has been successfully applied to some European species with irruptive or irregular movements (Newton et al. 2006; Marquiss et al. 2012; Fox and Hobson 2010) and presents a promising means of linking birds between winter locations and their breeding grounds for North American species of finch with similar behaviors.
The overall objective of this study was to estimate the molt origins and movement patterns of four winter finch species, evening grosbeak (Coccothraustes vespertinus), pine grosbeak (Pinicola enucleator), common redpoll (Acanthis flammea), and hoary Redpoll (A. hornemanni), as a means of improving our understanding of the linkages between breeding and wintering sites. To do this, we measured δ2Hf from birds captured during winter near residential bird feeders at sites across three regions in Ontario, Canada. Specifically, our goals were to: (1) improve our understanding of linkages between breeding/natal and wintering sites, (2) determine if demographic or spatial patterns exist in the winter movements of finches, and (3) use winter site fidelity as a means of evaluating the timing and consistency (regular vs. irregular) of movement patterns among these northern breeding species. Based on movement patterns from historical Nearctic and Palearctic banding recovery data (Brewer et al. 2000), we predicted that winter finches would undertake primarily longitudinal migrations. Based on the body size hypothesis (Prescott 1991, 1994; Macdonald et al. 2016), we predicted that smaller-bodied females would winter at lower latitudes than males, and should have lower mean δ2Hf values than males wintering at sites of similar latitude (i.e. more distant breeding grounds).
Methods
Study area and population
We conducted our study during the winters (December–April) of 2014–15 and 2015–16 in three regions, clustered at similar longitudes, in Ontario, Canada, within the non-breeding range of our study species (Fig. 1). Within each region, we visited an average of six local residential bird feeding or semi-permanent banding sites (n = 18, range 3–8) to increase sample sizes of each target species given the high variation in the presence and abundance of winter finch flocks within and between successive winters. Some locations or sites within each region were not revisited in both winters, given the absence or low abundance of target species between years. Given high variation in winter weather and the resulting logistical challenges to accessing some sites frequently or reliably, we were unable to standardize daily or seasonal trapping effort.

Location of sampling sites and study regions in Ontario where winter finches were trapped and feathers sampled for isotope analysis during the winters of 2014–15 and 2015–16. Triangles indicate sampling sites in the western study region; stars indicate sites in the central study region; squares indicate sites in the eastern study region
Birds were captured using mist nets and baited ground and feeder traps at residential bird feeders or established banding sites. Once captured, we aged and sexed birds based on plumage (Pyle 2008), measured wing chord, and fitted each individual with a uniquely numbered US Geological Survey metal band. Prior to release, a single feather (P1), grown during the post-juvenile or pre-basic molt on the natal or breeding grounds (Pyle 2008), was pulled for isotope analysis. We inferred site fidelity and fine-scale movement patterns of birds, both within and between years, through recaptures of banded birds or re-sightings of color-banded birds. Captured birds were marked using a single darvic color band unique to each cohort (e.g., species × site × month) and were not individually marked, so we were only able to estimate site fidelity of re-sighted birds by taking the single largest count of marked birds by cohort observed at any one time. Semi-continuous banding operations (e.g., at least one bi-weekly trapping event) were conducted at three sampling sites (two in the eastern region, and one in central region) during both winters where birds were routinely recaptured.
Sample preparation and stable isotope analysis
Prior to isotope analysis, individual feathers were cleaned of surface oils and contaminants using a 2:1 chloroform:methanol solvent rinse. Samples were weighed (350 μg) into silver capsules and loaded into a Eurovector Uni-Prep carousel (60 °C) interfaced with a Eurovector elemental analyser (Eurovector, Milan, Italy). Following pyrolytic combustion on glassy carbon at 1350 °C, H2 gas was introduced into a Thermo (Breman, Germany) Delta V Plus isotope-ratio mass spectrometer under continuous flow. Analysis of δ2H was conducted using the comparative equilibration method of Wassenaar and Hobson (2003) with three calibrated keratin reference materials (CBS, δ2H = − 197‰; KHS, δ2H = − 54.1‰; SPK, δ2H = − 121.6‰) corrected for linear instrumental drift. Based on within-run analyses (n = 5 each) of the keratin standards, measurement error was approximately ± 2‰ for hydrogen isotopes in feather (δ2Hf). All δ2H values are reported relative to the Vienna Standard Mean Ocean Water (VSMOW)—Standard Light Antarctic Precipitation (SLAP) scale.
Statistical analysis
We grouped hatch year (HY, before 31 Dec) and second year (SY, after 31 Dec), and after hatch year (AHY, before 31 Dec) and after second year (ASY, after 31 Dec) age classes into ‘HY’ and ‘AHY’, respectively (Macdonald et al. 2016). We used multi-way analysis of variance (ANOVA) in a general linear model (GLM) to test for differences in δ2Hf by age, sex, and region for each species, separately. We found no significant differences in δ2Hf for each species between years (p > 0.2), so we used data for both years combined. Our interaction terms were not statistically significant (p > 0.1), so we used type II sums of squares. To determine if wing chord could also be used as a proxy for migration distance (Winkler and Leisler 1992), we examined the relationship between δ2Hf and wing chord using a separate one-way ANOVA for each species. Analyses were performed in the R statistical computing environment version 3.5.0 (R Development Core Team 2017) using the ‘car’ package. The level of significance for all tests was set at α = 0.05.
Assignment to origin
Individual birds of each species were assigned to their respective breeding ranges using a likelihood-based assignment method (Royle and Rubenstein 2004). We employed an algorithm relating variation in δ2Hf to differences in amount-weighted growing season average precipitation δ2H (δ2Hp; Hobson et al. 2012; Bowen et al. 2005). We created a δ2Hf isoscape surface for non-ground foraging, short-distance migrants using the algorithm from Hobson et al. (2012): δ2Hf = − 22.98 + 0.95 × δ2Hp. We then used the standard deviation of the residuals from the regression (SD = 9.7 ‰) in the assignments. To limit assignments to biologically plausible origins, we clipped the calibrated δ2Hf isoscape to the respective digital breeding ranges for three of the species from BirdLife International and NatureServe (2011) in a Geographic Information System (GIS). For hoary redpoll, we modified the species’ digital range from Ridgely et al. (2012) by including approximate breeding, over-wintering and year-round ranges in Greenland based on Knox and Lowther (2000a, b), since Ontario is a plausible wintering area for birds from that region. We assumed that three species with circumpolar distributions captured at our study sites were unlikely to originate from Eurasian breeding grounds. Therefore, we removed those regions from the assignment analysis. Band recovery data have shown that winter finch species can traverse the continental divide (i.e., Rocky Mountains; Brewer et al. 2000), so we retained all breeding areas for all species in North America in our analyses.
For each individual feather sample (bird), we assessed the likelihood that each cell in the calibrated isoscape represented a potential origin for the individual using the normal probability density function and arbitrarily applied a 2:1 odds ratio criterion to delimit whether a pixel was a potential area of origin (1) or not (0). This odds ratio provides a reasonable tradeoff between depicting the correct origins with precision of the assignments (Hobson et al. 2012; Hobson and Wassenaar 2019). The resulting binary surfaces were summed for all individuals in a ‘population’ where pixel-level data reflect the number of individuals potentially originating at a given pixel were included in a final population depiction. We conducted multiple assignment to origin analyses for each species that assess differences in δ2Hf between ages, sexes, and regions based on initial GLM results. Manipulation of digital files and statistical analyses were conducted using functions in the ‘raster’ package (Hijmans et al. 2015) in the R statistical computing environment version 3.5.0 (R Development Core Team 2017).
Results
Isotopic composition
We collected feather samples from 814 individual birds from four species of winter finches within the subfamily Carduelinae (Table 1). We detected differences in δ2Hf values between species, with evening grosbeaks having the highest overall mean values consistent with more southern origins and common redpolls the lowest representing more northern origins. Mean δ2Hf values within species were similar between years, suggesting movement patterns between breeding and wintering sites were consistent during the 2 years of our study (Fig. 2). We found differences in δ2Hf values by sex, age, and sampling region, though these differences were not consistent between species (Table 2). We found a significant negative relationship between δ2Hf values and wing chord in common redpolls (F1,372 = 7.48, p < 0.01) but not for hoary redpolls (F1,26 = 1.31, p = 0.26), evening grosbeaks (F1,75 = 0.26, p = 0.61), or pine grosbeaks (F1,139 = 0.11, p = 0.74).

Boxplot showing variation in isotope values of feathers (δ2Hf) for four winter finch species between the winters of 2014–15 and 2015–16
Assignment to origin
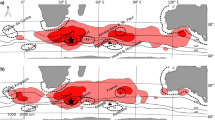
The probabilistic assignment analyses revealed substantial spatial structure of molt origins for winter finches sampled in Ontario. Both evening and pine grosbeaks had overall highest likelihoods of originating from the western portion of their breeding ranges in western Canada. Adult evening grosbeak had the highest probability of origins in the northern Rockies, central Manitoba and a lower probability of origin from some of the eastern portions of the breeding range (Fig. 3a). Likely areas of origin for hatch-year birds were constrained to western Manitoba, eastern Saskatchewan, and south-central British Columbia (Fig. 3b). Female pine grosbeak probability of origin included areas extending from the southern Northwest Territories, northern British Columbia, and much of the central and eastern part of the range in Alaska (Fig. 4a). Likely origins of male pine grosbeak were similar to females, though fewer males appeared to originate from the central Yukon and Northwest Territories (Fig. 4b). Likely origins of pine grosbeaks sampled in the western region of our study area included a large portion of the breeding range from the southern Northwest Territories, northern British Columbia, and much of the latitudinal range in eastern Alaska (Fig. 4c). Pine grosbeaks sampled in the central region had potential origins similar to birds sampled in the west, though fewer birds originated in the central Yukon and Northwest Territories than birds from the western region (Fig. 4d). Results for evening and pine grosbeaks suggest birds wintering in Ontario generally migrate longitudinally between breeding or natal sites and wintering areas.

Assignment of probable breeding or natal origins of evening grosbeaks in North America using feather δ2H of adult (AHY) and immature (HY) birds. Assignments were restricted to the known breeding range (black outline). Legend values indicate the potential number of individuals originating from a single cell in the raster

Assignment of probable breeding or natal origins of pine grosbeaks in North America using feather δ2H of females, males, birds from the western study region, and birds from the central study region. Assignments were restricted to the known breeding range (black outline). Legend values indicate the potential number of individuals originating from a single cell in the raster
Based on the highest probability of origin, the provenance of common and hoary redpolls was concentrated on Ellesmere Island in the Canadian high Arctic and/or northwest Greenland (Fig. 5a, b). This suggests the majority of redpolls wintering in Ontario generally moved latitudinally between breeding or natal sites and wintering areas for the years we sampled. Common redpolls wintering in central and western regions of our study area largely originated from the Canadian high Arctic. Some of these birds may have also originated from higher elevations in the western part of the range, including the Mackenzie Mountains in the central Yukon and western Northwest Territories and the St. Elias Mountains in southwest Yukon (Fig. 6a). Common redpolls in the eastern region of our study area appeared to have breeding origins from more southern and western breeding locations, such as Baffin Island, central Nunavut, and higher elevation sites in the Northwest and Yukon Territories, and eastern Alaska (Fig. 6b).

Assignment of probable breeding or natal origins using feather δ2H of common redpolls, and hoary redpolls, in North America. Assignments were restricted to the known breeding range (black outline). Legend values indicate the potential number of individuals originating from a single cell in the raster

Assignment of probable breeding or natal origins of common redpolls in North America using feather δ2H of birds from western and central study regions, and birds from the eastern study region. Assignments were restricted to the known breeding range (black outline). Legend values indicate the potential number of individuals originating from a single cell in the raster
Site fidelity and movement
Recaptures of banded birds or re-sightings of color-marked birds, both within and between winters, suggests overall fidelity of finches to our capture sites was low, ranging from 0 to 21.1% (Table 3). Grosbeaks were seldom encountered at sites during the same winter and were often recaptured or re-sighted at sites between winters. The mean duration between recaptures or re-sightings was 208.7 days (n = 11) for evening grosbeaks and 313.8 days (n = 10) for pine grosbeaks. Evening grosbeaks were recaptured or re-sighted at the same sites late in the first winter and early the subsequent winter. Evening grosbeaks were more likely to move between sites within the same region, either within or between winters, with mean distances between encounters of 37.9 km (n = 11). Mean distances between encounter sites in this species was strongly influenced by a single color-banded Evening Grosbeak re-sighted at a feeder 310 km from the original banding location between winters. Therefore, with the exception of this individual, most recaptures or re-sightings were at small spatial scales, often among closely adjacent sites in the same region. Pine grosbeak showed no apparent movement among sites and were only recaptured or re-sighted for brief periods at their original capture sites, often within a few weeks of the original capture date the following winter. Redpolls were only re-sighted or recaptured within the same winter and remained at original capture sites for an average of 23.8 days (n = 207) for common redpolls and 8.5 days (n = 8) for hoary redpolls. Redpolls were rarely detected moving among sites in a region within the same winter, though two color-banded common redpolls were encountered at distances of 4.9 and 20.5 km from their original sites during the same winter 67 and 24 days, respectively, between encounters.
Discussion
Our investigation into probable origins of four species of winter finch captured at sites in Ontario during two winters revealed some consistency in the spatial and temporal movement patterns within species, but less consistent patterns among species. We identified broad potential breeding range catchment areas for each species with a probabilistic assignment to origin approach using δ2Hf, which to our knowledge is the first application of this technique to species of seed-eating birds in the subfamily Carduelinae in North America. Using recaptures of banded birds and re-sightings of color-marked birds, we observed subtle differences in the spatial and temporal patterns of both within- and between-year movements between species. Redpolls, with limited fidelity to our study sites, both within- and between-years, had less apparent structure to their movements than grosbeaks.
For both species of grosbeak, our results suggested that the greatest number of birds across study sites originated from western portions of the breeding range, although some individuals may have derived from eastern portions of the breeding range. This was consistent with our hypothesis of birds moving longitudinally (west–east) between breeding and wintering locations. Brewer et al. (2000) suggested a similar longitudinal pattern in evening grosbeak movements based on several decades of band recovery data in Canada. Band recoveries of pine grosbeaks in North America have been limited, so our results provide the first direct evidence of large-scale longitudinal movements between breeding and wintering sites in Nearctic populations of this species. Adkisson (1999) suggested that pine grosbeaks exhibit limited annual movements, except during infrequent irruption events, when eastern birds were thought to make southward movements along a broad front during winter in response to limited availability of mast-seed crops at higher latitudes. Given the similarity of patterns observed within this species between winters, these longitudinal (e.g., west–east) seasonal movements between breeding and wintering sites may be more common for this species in North America. Given our breeding origin assignments and the apparent structure of birds at winter capture locations in this species, there is some evidence of a chain migration pattern (Berthold 2001). In this case, the longitudinal order of breeding populations was similar on the wintering grounds, with birds from western breeding areas occupying more western wintering sites.
For redpolls, our results indicate that birds sampled at wintering sites in Ontario potentially originated from a broad catchment, including western, central, and northern portions of the breeding ranges. This result was inconsistent with our hypothesis of longitudinal movements between breeding and wintering locations. However, our results represent some of the only known linkages between redpoll breeding and wintering sites during several successive seasons. Previous studies of banded redpolls concluded that longitudinal movements were indicative of shifts in wintering areas, with an east–west displacement of birds alternating between successive winters, rather than movements within a single season (Troy 1983; Brewer et al. 2000; Wilson 2008). While redpolls apparently alternate their southward movements between longitudinal extremes of their range in North America, they do not appear to entirely vacate their normal winter range, even in major irruption years (Hochachka et al. 1999). The winter of 2014–15 was likely an irruption year for redpolls into our study area, with > 85% of total redpoll captures occurring during this first winter, despite comparable sampling effort between years. In the winter of 2015–16, far fewer birds were encountered and captures were generally limited to the most northerly (western region) study sites. The ultimate driver of this presumed biannual pattern remains unclear and is likely not limited to large-scale variation in food resources or environmental variables (Koenig 2001; Koenig and Knops 2001; Strong et al. 2015). Expanding winter feather sampling for isotope analysis across broader spatial scales and longer time scales will provide a better understanding of spatiotemporal movement dynamics of winter finches.
We detected differences in δ2Hf based on sex, age, and region, though these differences were not consistent among species. Values of δ2Hf and the probable origins of males and females captured at wintering sites in Ontario differed for all species except evening grosbeak. As predicted, female pine grosbeaks and common redpolls had significantly lower δ2Hf values than males, suggesting they travelled greater distances between breeding and wintering sites than males. Differential migration between sexes has been demonstrated in evening grosbeaks in previous studies (Prescott 1991, 1994), though, despite females having slightly higher mean δ2Hf values, differences between sexes were not statistically significant in our study. Male hoary redpolls had significantly lower δ2Hf values than females, suggesting they travelled greater distances between breeding and wintering sites than females. However, differences in δ2Hf between sexes within species were typically small (< 6‰) and may result from differences in molt timing or location, body size and/or nutrient allocation for sympatrically breeding birds (but see Nordell et al. 2016; Rushing et al. 2014). Therefore, we provide weak evidence for differential migration by sex for three species of winter finch and further research is required.
We found no apparent relationship between δ2Hf values and wing chord in all species except common redpoll, for which we found a statistically significant negative relationship. The lack of spatial structure in wing chord for most of our study species is expected given the irruptive and nomadic nature of winter finches (Winkler and Leisler 1992). The result for common redpoll was not expected, since this species is thought to be more irruptive than the other species (Troy 1983) but might suggest more consistent spatial movement and segregation patterns for this species even during irruption events. However, we sampled for only 2 years and future research should investigate this pattern over multiple irruption and non-irruption years. Evening grosbeaks were also the only species in which we detected differences in origins of birds based on age, with HY birds having lower δ2Hf values than AHY birds. Prescott (1991) did not detect differences in the distribution of age classes in evening grosbeaks during winter, but banding records were distributed across large temporal and spatial ranges, which may not reflect the full extent of winter movements in this species. Differences in δ2Hf have been found for adult and juvenile birds from the same breeding site potentially due to discrepancies in diet and stress (Haché et al. 2012; Langin et al. 2007) and interpretations of this result should, therefore, take this into account.
Site fidelity during winter, based on both recaptures of banded birds and re-sightings of color-marked birds, was low in all four species. For evening grosbeaks, fidelity was comparable to rates reported elsewhere in North America (2–14%; Clinch 1956; Balph and Lindahl 1978; Yunick 1983). Movement among sites in evening grosbeaks were mostly local, occurring between adjacent sampling sites in the same region. Similarly, birds were re-sighted within a few days of original capture at the same sites, or were re-sighted in the same area early the following winter. Winter site fidelity in pine grosbeaks averaged lower in our study than for other published accounts (e.g., 43%; Adkisson 1999). Pine grosbeaks were re-sighted at capture sites, generally in the same month as the original capture date from the previous winter, suggesting that, for a portion of the population, they followed the same routes and general timing between breeding and wintering locations in subsequent winters. Site fidelity in redpolls was also generally low, though in contrast to grosbeaks, recaptures or re-sightings of color-banded birds were only within the same winter and typically at original capture sites. Overall site fidelity, based on within-year recaptures and re-sightings, in common redpolls (7.6%) was lower than the rate observed by Yunick (1983) of 34% in Schenectady, NY. Redpolls continue moving southwards as winter proceeds (Wilson 2008), stopping opportunistically as they encounter food resources (Hochachka et al. 1999), which was generally supported by the relatively short residency time at our study sites. Despite overall low rates of fidelity, the different residency times and movement patterns both within and between winters exhibited by each species suggests they employ different strategies which warrant further study.
References
Adkisson CS (1999) Pine grosbeak (Pinicola enucleator), version 2.0. In: Poole AF, Gill FB (eds) The birds of North America. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.456
Balph MH, Lindahl AM (1978) Winter philopatry of Evening grosbeaks in northern Utah. N Am Bird Bander 3:149–151
Berthold P (2001) Bird migration: a general survey. Oxford University Press, New York, p 272
Birdlife International and NatureServe (2011) Bird species distribution maps of the world. Birdlife International, Cambridge
Bock CE, Lepthien LW (1976) Synchronous eruptions of boreal seed-eating birds. Am Nat 110:559–571
Bonter DN, Harvey MG (2008) Winter survey data reveal rangewide population decline in Evening grosbeak populations. Condor 110(2):376–381
Bowen GJ, Wassenaar LI, Hobson KA (2005) Global application of stable hydrogen and oxygen isotopes to wildlife forensics. Oecologia 143(3):337–348
Brewer D, Diamond AW, Woodsworth AJ, Collins BT, Dunn EH (2000) Canadian Atlas of bird banding: Doves, Cuckoos, and Hummingbirds, through Passerines, 1921–1995, 1st edn. Canadian Wildlife Service, Ottawa
Clinch FA (1956) Evening grosbeak banding at Watertown, New York. Kingbird 6:77–80
COSEWIC (2016) COSEWIC assessment and status report on the Evening grosbeak, Coccothraustes vespertinus, in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, p 64
Dunn EH, Altman BL, Bart J, Beardmore CJ, Berlanga H, Blancher PJ, Butcher GS, Demarest DW, Dettmers R, Hunter WC, Iñigo-Elias EE, Panjabi AO, Pashley DN, Ralph CJ, Rich TD, Rosenberg KV, Rustay CM, Ruth JM, and Will TC (2005). High priority needs for range-wide monitoring of North American landbirds. Partners in Flight Technical Series No. 2, Laurel, Maryland, USA. https://www.partnersinflight.org/wp-content/uploads/2017/03/PIF-Technical-Series-02-Monitoring-Needs.pdf. Accessed 14 May 2019
Fox A, Hobson KA (2010) Linking breeding and wintering areas of ‘northern trumpeting’ bullfinches Pyrrhula p. pyrrhula by color rings and deuterium stable isotopes. Bird Study 57:125–127
Fudikar AM, Schmidt A, Hau M, Quetting M, Partecke J (2013) Female-biased obligate strategies in a partially migratory population. J Anim Ecol 82(4):863–871
Gilihan SW, Byers BE (2001) Evening grosbeak (Coccothraustes vespertinus) version 2.0. In: Poole AF, Gill FB (eds) The birds of North America. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.599
Haché S, Hobson KA, Villard MA, Bayne EM (2012) Assigning birds to geographic origin using feather hydrogen isotope ratios (δ2H):importance of year, age, and habitat. Can J Zool 90:722–728
Hijmans RJ, van Etten J, Cheng J, Mattiuzzi M, Sumner M, Greenberg JA, Perpinan Lamigueiro O, Bevan A, Racine EB, Shortridge A, Ghosh A (2015) Raster: geographic data analysis and modeling. Version 2.5–2. https://cran.r-project.org/package=raster. Accessed 21 Aug 2019
Hobson KA, Wassenaar LI (2019) Tracking animal migration with stable isotopes, 2nd edn. Academic Press, San Diego, p 268
Hobson KA, Van Wilgenburg SL, Wassenaar LI, Larsen K (2012) Linking hydrogen (δ2H) isotopes in feathers and precipitation: sources of variance and consequences for assignment to isoscapes. PLoS ONE 7(4):e35137
Hobson KA, Van Wilgenburg SL, Faaborg J, Toms J, Rengifo C, Llanes Sosa A, Aubrey Y, Brito Aguilar R (2014) Connecting breeding and wintering grounds of neotropical migrant songbirds using stable hydrogen isotopes: a call for an isotopic atlas of migratory connectivity. J Field Ornithol 85(3):237–257
Hochachka WM, Wells JV, Rosenberg KV, Tessaglia-Holmes DL, Dhondt AA (1999) Irruptive migration of Common redpolls. Condor 101:195–204
Knox AG, Lowther PA (2000a) Common Redpoll (Acanthis flammea), version 2.0. In: Poole AF, Gill FB (eds) The birds of North America. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.543
Knox AG, Lowther PE (2000b) Hoary Redpoll (Acanthis hornemanni), version 2.0. In: Poole AF, Gill FB (eds) The birds of North America. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.544
Koenig WD (2001) Synchrony and periodicity of eruptions by boreal birds. Condor 103(4):725–735
Koenig WD, Knops JMH (2001) Seed-crop size and eruptions of North American boreal seed-eating birds. J Anim Ecol 70:609–620
Langin KM, Reudink MW, Marra PP, Norris DR, Kyser TK, Ratcliffe LM (2007) Hydrogen isotopic variation in migratory bird tissues of known origin: implications for geographic assignment. Oecologia 152:449–457
Lepage D, Francis CM (2002) Do feeder counts reliably indicate bird population changes? 21 years of winter bird counts in Ontario. Can Condor 104(2):255–270
Macdonald CA, McKinnon EA, Gilchrist HG, Love OP (2016) Cold tolerance, and not earlier arrival on breeding grounds, explains why males winter further north in an Arctic-breeding songbird. J Avian Biol 47:7–15
Marquiss M, Newton I, Hobson KA, Kolbeinsson Y (2012) Origins of irruptive migrations by common Crossbills Loxia curvirostra into northwestern Europe revealed by stable isotope analysis. Ibis 154(2):400–409
Newton I (2008) The migration ecology of birds. Academic Press, London, p 984
Newton I (2012) Obligate and facultative migration in birds: ecological aspects. J Ornithol 153:171–180
Newton I, Hobson KA, Fox AD, Marquiss M (2006) An investigation into the provenance of Northern Bullfinches Pyrrhula p. pyrrhula found in winter in Scotland and Denmark. J Avian Biol 37:431–435
Nordell CJ, Haché S, Bayne EM, Sólymos FKR, Godwin CM, Krikun R, Pyly P, Hobson KA (2016) Within-site variation is feather stable hydrogen isotope (δ2Hf) values of boreal songbirds: implications for assignment to origin. PLoS ONE 11:e0163957
Prescott DRC (1991) Winter distribution of age and sex classes in an irruptive migrant, the Evening grosbeak. Condor 93:694–700
Prescott DRC (1994) Intraspecific and geographical trends in body size of a differential migrant, the Evening grosbeak. Auk 111:693–702
Pyle P (2008) Identification guide to North American birds, part I: Columbidae to Ploceidae. Slate Creek Press, Bolinas
R Development Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ralston J, King DI, DeLuca WV, Niemi GJ, Glennon MJ, Scarl JC, Lambert JD (2015) Analysis of combined data sets yields trend estimates for vulnerable spruce-fir birds in northern United States. Biol Conserv 187:270–278
Ridgely RS, Allnutt TF, Brooks T, McNicol DK, Mehlman DW, Young BE, Zook JR, BirdLife International (2012) Digital distribution maps of the birds of the western hemisphere, version 5.0. In: International BirdLife (ed) Bird species distribution maps of the world, version 2.0. BirdLife International, Cambridge
Roy C, Michel NL, Handel CM, Van Wilgenburg SL, Burkhalter JC, Gurney KEB, Messmer DJ, Princé K, Rushing CS, Saracco JF, Schuster R, Smith AC, Smith PA, Sólymos P, Venier LA, Zuckerberg B (2019) Monitoring boreal avian populations: how can we estimate trends and trajectories from noisy data? Avian Conserv Ecol 14(2):8. https://doi.org/10.5751/ACE-01397-140208
Royle JA, Rubenstein DR (2004) The role of species abundance in determining breeding origins of migratory birds with stable isotopes. Ecol Appl 14(6):1780–1788
Rubenstein DR, Hobson KA (2004) From birds to butterflies: animal movement patterns and stable isotopes. Trends Ecol Evol 19(5):256–263
Rushing CS, Ryder TB, Saracco JF, Marra PP (2014) Assessing migratory connectivity for a long-distance migratory bird using multiple intrinsic markers. Ecol Appl 24:445–456
Strong C, Zuckerberg B, Betancourt JL, Koenig WD (2015) Climatic dipoles drive two principal modes of Norther American boreal bird irruption. Proc Natl Acad Sci USA 112:E2795–E2802
Stutchbury BJM, Taylor SA, Done T, Gow E, Kramer PM, Tautin J, Fox JW, Afanasyev V (2009) Tracking long-distance songbird migration by using geolocators. Science 323(5916):896
Taylor PD, Crewe TL, Mackenzie SA, Lepage D, Aubry Y, Crysler Z, Finney G, Francis CM, Guglielmo CG, Hamilton DJ, Holberton RL, Loring PH, Mitchell GW, Norris DR, Paquet J, Ronconi RA, Smetzer JR, Smith PA, Welch LJ, Woodworth BK (2017) The Motus wildlife tracking system: a collaborative research network to enhance the understanding of wildlife movement. Avian Conserv Ecol 12(1):8
Troy DM (1983) Recaptures of redpolls: movements of an irruptive species. J Field Ornithol 54(2):146–151
Wassenaar LI, Hobson KA (2003) Comparative equilibration and online technique for determination of non-exchangeable hydrogen of keratins for use in animal migration studies. Isot Environ Health Stud 39:211–217
Widrlechner MP, Dragula SK (1984) Relation of cone-crop size to irruptions of four seed-eating birds in California. Am Birds 38:840–846
Wilson WH Jr (2008) Winter movements of Common redpolls and Pine Siskins within and between winters. N Am Bird Bander 33(4):169–174
Winkler H, Leisler B (1992) On the ecomorphology of migrants. Ibis 134(1):21–28
Yunick RP (1983) Winter site fidelity of some northern finches (Fringillidae). J Field Ornithol 54(3):254–258
Acknowledgements
Many thanks to M. Caskey, B. Fried, B. Murphy, and C. Sharp, who assisted with capturing, banding, and collecting feather samples for this study. We thank R. Tafel and B. Ratcliff for providing logistical support during feather collection. Thanks also to the many homeowners, including L. Anderson, D. Brunet, D. Chalifoux, S. Chalifoux, M. Desjardins, B. Fagan, S. Fagan, K. Gowing, B. Horth, G. Kideres, N. Maurice, M. Maurice, B. McCracken, J. McCracken, S. Momy, R. Sharp, and M. Young, who generously allowed us to collect feather samples at their feeders. We thank C. Gryba and G. Koehler at the Stable Isotope Hydrology and Ecology Lab of Environment and Climate Change Canada, and F. Longstaffe and T. Plint at the Laboratory of Stable Isotope Science at the University of Western Ontario for preparing and analyzing feather samples. Thanks to E. Howat, J. Put, and N. Spencer for assisting with data management, analysis, and mapping. R. Weeber and two anonymous reviewers provided valuable input and comments on a previous version of the manuscript. This work was supported by an operating grant to KCH from Environment and Climate Change Canada, which required approval of the manuscript before publication. All methods were in compliance with Canadian law and conducted with approval under appropriate permits.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical standards
All procedures were approved by the Wildlife Eastern Animal Care Committee of Environment and Climate Change Canada (14KH01, 15KH01) and the Canadian Bird Banding Office (#10746).
Additional information
Communicated by C. G. Guglielmo.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hannah, K.C., Kardynal, K.J. & Hobson, K.A. Birds of a feather don’t always flock together: variation in molt origins and movement patterns of winter finches in Ontario. J Ornithol 161, 609–620 (2020). https://doi.org/10.1007/s10336-020-01760-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-020-01760-1