
Abstract
Dunlin (Calidris alpina) females migrate ahead of males towards their wintering grounds, but the factors influencing the timing of this migration is incompletely understood. In our long-term (1991–2000) study of the autumn migration phenology of male and female dunlins, as adult and immature birds separately, we noted that the difference in mean migration dates between sexes was the greatest in years of high breeding success, reaching 5 days in immature birds and 7 days in adult birds. In other, relatively less successful years, this difference could reach 2 days in immature birds and 3 days in adult birds. As Dunlin females are not fully involved in parental care, the timing of their migration appears to be determined mainly by the timing of the start of the breeding season. In males, the timing of the autumn migration appears to depend primarily on the time spent on breeding and parental care. Hence, in years of high breeding success, with a high number of offspring, the males stayed longer at the breeding grounds. In the years with a high and moderate production of offspring, immature birds departed ahead of adults, probably due to their very low breeding success. A novel finding of our study is that in the years of overall low breeding success the opposite was true: immature birds migrated later than adults and much later than in other years. The plausible explanation is that lack of experience may result in greater tendency of immature birds to renest after failure of the first clutch, especially when high breeding failures of earlier arriving adult Dunlins may lead to decreasing competition at the breeding ground. However, in years of low breeding success, there was no difference in mean migration date between immature males and females, indicating that although immature birds stayed longer at the breeding grounds, they had a very low breeding success.
Zusammenfassung
Immature Alpenstrandläufer Calidris alpina ziehen in Jahren mit niedrigem Bruterfolg später in die Überwinterungsgebiete als Altvögel
Weibliche Alpenstrandläufer ziehen vor den Männchen in die Überwinterungsgebiete. Dieser Geschlechtsunterschied im mittleren Zugdatum war in Jahren mit hohem Bruterfolg am größten und betrug fünf Tage bei immaturen Tieren und sieben Tage bei Altvögeln. In anderen Brutsaisons betrug dieser Unterschied bis zu zwei Tagen bei immaturen Tieren und drei Tagen bei Altvögeln. Da sich weibliche Alpenstrandläufer weniger stark an der Brutpflege beteiligen als Männchen, wird ihr Zugzeitpunkt hauptsächlich durch den Beginn der Brutsaison bestimmt. Bei Männchen hingegen hängt der Beginn des Herbstzuges hauptsächlich von der für die Brutpflege benötigten Zeit ab. Daher bleiben Mannchen in Jahren mit vielen Nachkommen länger im Brutgebiet. In Jahren mit hohen und mittleren Nachkommenzahlen zogen immature Tiere vor den Altvögeln ab, wahrscheinlich aufgrund ihres sehr niedrigen Bruterfolgs. Ein neuer Befund ist, dass in Saisons mit insgesamt niedrigem Bruterfolg das Gegenteil der Fall war, d. h. immature Tiere zogen später als Altvögel und deutlich später als in anderen Saisons ab. Eine plausible Erklärung hierfür ist, dass immature Tiere aus Mangel an Erfahrung eher dazu neigen, nach Verlust des ersten Geleges erneut zu brüten, besonders wenn hohe Brutverluste der früher im Brutgebiet eintreffenden Altvögel die Konkurrenz im Brutgebiet verringern. In Jahren mit niedrigem Bruterfolg bestand jedoch kein Unterschied im mittleren Zugdatum zwischen immaturen Männchen und Weibchen, was darauf hindeutet, dass der Bruterfolg der immaturen Tiere sehr niedrig war, obwohl sie länger im Brutgebiet blieben.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Dunlin (Calidris alpina) is a migratory wader species of the southern Baltic Sea area during the autumn migration (Kube et al. 1994; Meissner et al. 2009). It is estimated that about 1–2 % of adults and more than 10 % of juveniles of the East Atlantic migratory population used stop-over sites located on the southern Baltic coast (Kube et al. 1994). The published data currently available on the migration phenology of the Dunlin in the Baltic region provide only a general description of the migration pattern of this species (Brenning 1987; Pettersson 1994; Meissner and Strzałkowska 2006). It is known that adults arrive at the Baltic Sea stop-over locations earlier than first-year birds (juveniles) and that from mid-August the proportion of juveniles in staging flocks gradually increases (Brenning 1987; Meissner and Strzałkowska 2006). Moreover, males migrate somewhat later than females because in this particular species they take care of chicks and thus stay longer at the breeding grounds (Soikkeli 1967; Zając 1980). Although a conspicuous annual variability in the migration timing of this species has been noted (Brenning 1987; Meissner and Sikora 1995), a long-term analysis of the autumn migration phenology of male and female Dunlins separately, both as adults and juveniles, is lacking.
Several factors may influence the timing of wader migration towards their wintering grounds (Blomqvist et al. 2002; Anthes 2004). In the Arctic, environmental conditions constrain the time available for breeding, and waders nesting there only replace a clutch after failure of a previous one at the early stage of incubation (Jamieson 2011). The timing of breeding of Arctic waders depends on a fine balance between many factors, of which the most important are food abundance, extent of snow cover early in the season and the abundance of predators (Meltofte et al. 2007; Smith et al. 2010). Hence, a late spring can delay clutch initiation and may result in the later departure of birds from the breeding grounds. Alternatively, in seasons of high predation pressure, waders tend to nest early in the season, which reduces the risk of predation, and also provides the advantage of an increased opportunity for renesting (Smith et al. 2010). Another factor influencing the timing of waders’ departure towards the wintering areas is breeding success (Meissner 2005; Barshep et al. 2012). In years when there has been no time for replacement of the clutch or when the predation pressure has been high, adult birds have been observed to depart relatively earlier on their autumn migration and to arrive earlier at stop-over sites (Blomqvist et al. 2002; Meissner 2005). However, it remains unknown if high predation pressure, which is the main factor leading to low breeding success in Arctic waders (Underhill et al. 1993), differentially affects adult birds and inexperienced immature birds breeding for the first time. This aspect has not been difficult to analyse, because in the majority of Arctic waders first-year birds stay at the wintering grounds until the next breeding season (Summers et al. 1995; Hockey et al. 1998). One exception is the Dunlin. Individuals of this species in their second calendar year of life (immatures) migrate to breeding grounds (Holmes 1966; Soikkeli 1967). This behaviour provides the opportunity to analyse autumn migration phenology of males and females separately, as both adult and immature birds, in years of high and low breeding success, which is the main aim of this study.
Materials and methods
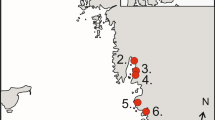
The studies were conducted in the vicinity of the mouth of the Reda river (Puck Bay, southern Baltic coast) between 1991 and 2000. In 1991–1995 the fieldwork was carried out mainly on a sandy spit near Rewa village and in 1996–2000 at a location about 2.5 km westwards, at the mouth of the Reda river (Fig. 1). Dunlins were on occasion trapped at these two sites simultaneously. During the study period, foraging waders were observed at several sites along the coast of a small bay situated between Rewa village and the river mouth. These observations and the high number of retraps showed that the whole area could be treated as one feeding site, and I consequently pooled data from all of the study for analysis. During the entire study period counts of waders were made every day between early July and the end of September. Wader trapping was conducted using walk-in traps (Meissner 1998). Wing length, total head length and bill length of each trapped Dunlin were measured (Meissner 2000). The age of the captured birds was determined according to the descriptions proposed by Meissner and Skakuj (2009), which categorises the Dunlins into three age classes: juveniles, immatures (birds in the second calendar year) and adults (>2 years). Juveniles were excluded from the main analyses because they migrate towards their overwintering sites until late October, and only about 70 % of juveniles migrate in the period covered by the fieldwork in our study area (Meissner and Sikora 1995). In terms of counts it was impossible to distinguish between adult and second-year birds; therefore, we classified the birds into only two age classes, i.e., juveniles and other birds based on breeding plumage. Although the proportion of juveniles based on the daily counts and trapping results were strongly correlated (r = 0.81) during the study period, we only used the results of counts to assess the overall breeding success in a given year (see Minton 2003 for details). The proportion of juveniles based on regular counts at stop-over sites during the autumn migration is considered to be the better method for assessing wader productivity than the results of walk-in trap catching in this period (Meissner 2007). The general linear model (GLM) was used to check differences in the proportion of juveniles in all counts, with year as the fixed factor. The proportions were arcus sinus square root transformed before the analysis.

The study area. Filled circles two ringing sites
The timing of the fieldwork differed slightly among seasons. Therefore, to standardise data, we defined a reference period—16 July to 28 September—when the counts and wader trapping activities were conducted every day in all years. This period covers the entire migration period of adults and immatures at southern Baltic stop-over sites (Meissner and Sikora 1995). All daily counts within the reference period were pooled to give one overall proportion of juvenile birds for each year of the study. We assumed that the differences in migration timing of juveniles in different years had only a slight impact on their proportion in the reference period. In each year of the study period, we noted that the peak of the number of migrating juvenile Dunlins occurred before the end of the reference period, following which there was a rapid decline in the number of migrating birds. The same pattern was observed in a previous study at other stop-over sites located near the study area (Meissner and Sikora 1995). Only in 1996 was the number of migrating juveniles still high on the last day of the reference period, with a sharp decrease occurring 2 days later. Hence, the proportion of juveniles in this year might be slightly underestimated.
The data on captured birds were used to compare the migration phenology of males and females within age groups. The sex of trapped birds was assessed using the discriminant functions based on the bill and wing lengths (Meissner and Pilacka 2008). In these functions the “0” value separates both sexes. Although the classification accuracy of these discriminant functions is quite high (over 97 % for females and 100 % for males), to increase the reliability of sexing, we treated birds with discriminant values ranging between −0.5 and 0.5 as unsexed. A total of 3,220 adults and 1,186 immatures with identified sex were included in further analyses.
A GLM [GLZ in Statistica (StatSoft, Tulsa, OK)] with a logarithm link function and normal error distribution (McCullagh and Nelder 1983) was used to account simultaneously for the effects of all independent variables. The trapping date of each bird (the day number in the season) was linked to sex, age and years of low, moderate and high breeding success, respectively. As a covariate, I used these latter three arbitrarily distinguished categories of breeding success of the breeding season—and not the proportion of juveniles—because this approach allows for the direct comparison of the mean migration date of adult and immature males and females in years of different breeding success/offspring production. Low breeding success was assigned to years in which juveniles comprised <40 % of all counted Dunlins, while high breeding success was assigned to those seasons in which juveniles comprised >60 % of all counted Dunlins; the category of moderate breeding success included all other years, i.e. when the proportion of juveniles was equal to 40–60 %. The Wald χ2 statistic was used to test for significant differences between groups. Mean migration date was calculated as an average and determined separately for males and females of both age categories.
The Akaike information criterion (AIC) was applied to evaluate which of the GLZ models explained the most variance and had the highest level of data support (Burnham and Anderson 2002; Burnham et al. 2011). The differences in AIC (∆AIC) values were calculated by subtracting the minimum AIC for the best-fitted model from the AIC of all subsequent candidate models. These differences were used to determine, which model provided the best description of the data on the basis of the fewest model parameters. Only models with a ∆AIC value of <9 were taken into account because these were considered to be acceptably similar in their ability to describe the data (Burnham et al. 2011) and models with larger ∆AIC have considerably less support (Burnham and Anderson 2002). The Akaike weight (AICw), which gives the relative support of the data for each model, was reported for all models to determine each model’s relative likelihood (Burnham and Anderson 2002; Burnham et al. 2011). All statistical procedures were performed using Statistica 10 software (StatSoft Inc. 2011).
Results
Breeding success in consecutive years
The differences in the proportion of juveniles between years were statistically significant (GLM: F 9,473 = 2.57, p = 0.007). Juveniles comprised >60 % of all counted Dunlins in 1991, 1993 and 1999, and these years were therefore considered to be years of high breeding success. In contrast, in 1992 and 1995 juvenile birds represented <40 % of all migrating Dunlins counted, and these years were therefore categorised as years of low breeding success. The remaining years of the study period were categorised as years of moderate breeding success (Fig. 2).

The proportion of juvenile Dunlins (Calidris alpina) (black bars) among all Dunlins counted in the reference period of each year of the study (1991–2000). H, M, L Years of high, moderate and low breeding success, respectively, based on the proportion of juvenile Dunlins counted
Phenology of autumn migration
The best GLZ model indicated a significant effect of breeding success, age and sex on the migration date, with a significant interaction of breeding success on age and sex (Table 1: model 1). This model was better supported by the data by more than twofold compared to the next best model, which included the interaction between age and sex (Table 1: model 2), as well as the model including the interaction between all three factors (Table 1: model 3). Immature Dunlins migrated earlier (regardless of sex and breeding success) than adult birds (Table 2: Wald χ2 = 40.3, p < 0.0001) and females migrated earlier than males (Table 2: Wald χ 2 = 50.0, p < 0.0001). However, breeding success had a significant effect on the mean migration date of Dunlins in terms of both age and sex. The largest difference in mean migration dates between adult males and adult females occurred in years of high breeding success, when it could reach 7 days; in other years, this difference reached a maximum of only 3 days (Cochran–Cox test: p < 0.05 in all three cases) (Fig. 3). The migration date of immature females preceded that of immature males by 2 and 5 days in years of moderate and high breeding success, respectively (Cochran–Cox test, p < 0.004 in both cases), whereas in years of low breeding success there was no statistically significant difference in the mean migration date between immature males and females (Cochran–Cox test, t′ = 0.547, p = 0.586).

Difference in mean migration dates of Dunlin males (M; circles and solid line) and females (F; rectangles and dashed line) in years of low, moderate and high breeding success, respectively. Thick line Immature birds (Imm), thin line adult birds (Ad). Symbols Mean migration date, whiskers 95 % confidence intervals
In years of high and moderate breeding success/high and moderate number of offspring, immature birds migrated earlier than adult birds. However, in seasons of low breeding success/low numbers of offspring the opposite behaviour was observed, with immature birds migrating much later than in other years and later than the adults (Fig. 3). Differences in mean migration date between adult and immature birds within each sex were statistically significant (Cochran–Cox test, p < 0.003 in all cases).
Mean migration dates of adult and immature birdss in consecutive seasons were not correlated (Kendall tau = 0.20, p = 0.42). Over the entire study period, the mean migration dates of immature birds were the latest in both years of low productivity (1992 and 1995), being even later than those of adults in the same seasons (Fig. 4).

Relationship between mean migration dates of adult and immature Dunlins in consecutive years. Black circles Years of low breeding success, grey circles years of moderate breeding success, white circles years of high breeding success. Diagonal line shows the equality of mean dates of migration of both age groups
Discussion
Sex differences in the timing of the autumn migration are common among species of subfamily Calidrinae, with the parent involved in chick rearing remaining longer at the breeding grounds—and hence migrating later—than their mate (Butler et al. 1987; Butler and Kaiser 1995; Summers and Nicoll 2004; Meissner 2005; Barshep et al. 2012). This is also the case in the Dunlin, a small wader which breeds in Arctic and subarctic regions: females take care of their broods for much less time than males and typically depart during the 10–12 days immediately following the hatch (Soikkeli 1967; Jamieson 2011). As a result, males stay longer at the breeding site and start on their autumn migration at various lengths of time after the departure of the females (Zając 1980). In years of low breeding success many males—most likely those which failed to breed successfully—are likely to depart with females, resulting in a smaller difference between the mean migration date of each sex. When breeding success is high, the majority of males stay longer at the breeding grounds, with the difference in migration dates being the greatest in both adult and immature birds. The influence of breeding success on migration phenology has also been described in the Knot (Calidris canutus) and Curlew Sandpiper (Calidris ferruginea), with various research groups reporting that in these species the parent taking care of chicks also migrate earlier than usual in years when the number of offspring is low (Meissner 2005; Figureola 2006; Barshep et al. 2012).
Females of many waders are not fully involved in parental care; consequently, the timing of their migration is determined mainly by the date of the start of the breeding season. In contrast, the beginning of the autumn migration in males depends mostly on the time spent on breeding and taking care of chicks. This behaviour may explain why—in our study—adult male dunlins migrated on average 4 days later in the years of high breeding success than in the years of low and moderate breeding success, while the mean departure date of the adult females’ migration varied only by 1 day (Fig. 3).
Adult (=experienced) Dunlins arrive at the breeding grounds somewhat earlier than immature birds nesting for the first time (Soikkeli 1967). This behaviour has also been observed in the Spotted Sandpiper (Actitis macularia) and Purple Sandpiper (Calidris maritima) with first-year birds nesting, on average, later than older ones (Oring et al. 1991; Summers and Nicoll 2004). The later arrival and earlier departure of immature dunlins from nesting areas has even led to the suggestion that they do not take part in breeding at all (Kozlova 1962). Wader nests which are initiated early in the season and chicks which hatch early in the season have a greater probability of survival than those hatched later in the breeding season (Hartman and Oring 2009). Birds which start to breed later in the season may therefore be under higher predation pressure because predator encounter rates and activity have been found to be positively affected by the cumulative number of initiated clutches (Sandercock 1998). Thus, immature Dunlins arriving later than adult birds are expected to have a low breeding success, especially as waders which breed for the first time have much more breeding failures than older, more experienced birds (Oring et al. 1991), which is also common in other bird species (Ollason and Dunnet 1978; Pugesek 1983; Forslund and Pärt 1995). The earlier departure of immature Dunlins from the breeding grounds is likely directly associated with nest failure, which is consistent with earlier observations that in Arctic waders, failed breeders migrate toward wintering grounds first (Oring and Lank 1982; Morrison 1984; Syroechkovski and Lappo 1994). It cannot be excluded that a fraction of immature males may not take part in breeding, being competitively inferior to older birds in terms of territory defence (Lozano et al. 1996), and hence depart earlier from the breeding grounds.
In general, immature Dunlins migrate earlier toward wintering grounds than adult birds (Gromadzka 1985; Meissner and Strzałkowska 2006), but the results of this study show that in years of low breeding success immature birds migrate later than adult birds. In the 2 years of low breeding success in 1992 and 1995, the abundance of rodents at the western Siberian breeding grounds was low and predation pressure was high (Blomqvist et al. 2002; http://www.arcticbirds.net). As a result, the number of waders’ and geese offspring was very low, particularly in 1992 (Ganter and Boyd 2000; Blomqvist et al. 2002).
Dunlins are capable of laying a replacement clutch after the failure of a previous nest (Soikkeli 1967; Jönsson 1991; Naves et al. 2008). However, there is a seasonal decline in the probability that a failed clutch would be replaced, and replacement has been observed only in the early phase of the breeding period (Jamieson 2011). Hence, a lack of experience may result in the greater tendency of immature birds to renest after the failure of the first clutch. High breeding failures of earlier arriving adult Dunlins may lead to decreasing competition for breeding sites and increase the opportunity for immatures to nest. However, in years of low breeding success, there was no difference in mean migration date of immature males and females, which indicates that although immatures stayed longer at the breeding grounds, they had a very low breeding success.
References
Anthes N (2004) Long-distance migration timing of Tringa sandpipers adjusted to recent climate change. Bird Study 51:203–211
Barshep Y, Meissner W, Underhill LG (2012) Timing of migration of the curlew sandpiper (Calidris ferruginea) through Poland in relation to Arctic breeding conditions. Ornis Fennica 89:120–129
Blomqvist S, Holmgren N, Åkesson S, Hedenström A, Pettersson J (2002) Indirect effects of lemming cycles on sandpiper dynamics: 50 years of counts from southern Sweden. Oecologia 133:146–158
Brenning U (1987) Der herbstliche Zug des Alpenstrandlaufers (Calidris alpina) im Naturschutzgebiet Langenwerder (Wismar-Bucht/Poel). Ber Vogelwarte Hiddensee 8:4–19
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Burnham KP, Anderson DR, Huyvaert KP (2011) AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav Ecol Sociobiol 65:23–35
Butler RW, Kaiser GW (1995) Migration chronology, sex ratio, and body mass of Least sandpipers in British Columbia. Wilson Bull 107:413–422
Butler RW, Kaiser GW, Smith GEJ (1987) Migration chronology, length of stay, sex ratio, and weight of Western sandpipers, (Calidris mauri) on the south coast of British Columbia. J Field Ornithol 58:103–111
Figureola J (2006) The role of breeding success and parental care in determining sexual differences in the timing of southward migration of the curlew sandpiper Calidris ferruginea through Tunisia. Int Wader Stud 19:148–150
Forslund P, Pärt T (1995) Age and reproduction in birds––hypotheses and tests. Trends Ecol Evol 10:374–378
Ganter B, Boyd H (2000) A tropical volcano, high predation pressure, and the breeding biology of Arctic waterbirds: a circumpolar review of breeding failure in the summer of 1992. Arctic 53:289–305
Gromadzka J (1985) Dunlin––Calidris alpina (L.). In: Viksne IA, Mihelson HA (eds) Migrations of birds of Eastern Europe and Northern Asia. Gruiformes–Charadriiformes. Nauka, Moscow, pp 193–220 (in Russian)
Hartman CA, Oring LW (2009) Reproductive success of long-billed curlews (Numenius americanus) in northeastern Nevada hay fields. Auk 126:420–430
Hockey PAR, Turpie JK, Velásquez CR (1998) What selective pressures have driven the evolution of deferred northward migration by juvenile waders? J Avian Biol 29:325–330
Holmes RT (1966) Breeding ecology and annual cycle adaptations of red-backed sandpipers (Calidris alpina) in northern Alaska. Condor 68:3–46
Jamieson SE (2011) Pacific dunlin Calidris alpina pacifica show a high propensity for second clutch production. J Ornithol 152:1013–1021
Jönsson PE (1991) Reproduction and survival in a declining population of the southern dunlin Calidris alpina schinzii. Wader Study Group Bull 61:56–68
Kozlova EV (1962) Fauna of USSR: Charadriiformes, Limicoiae, vol. 2, sec. I, part 3. Zoolological Institute of the Russian Academy of Sciences, Moscow/St. Petersburg (in Russian)
Kube J, Rösner H-U, Behmann H, Brenning U, Gromadzka J (1994) Der Zug des Alpenstrandläufers (Calidris alpina) an der südlichen Ostseeküste und im Schleswig-Holsteinischen Wattermeer in Sommer und Herbst 1991. Corax 15:73–82
Lozano GA, Perreault S, Lemon RE (1996) Age, arrival date and reproductive success of male American Redstarts Setophaga ruticilla. J Avian Biol 27:164–170
McCullagh P, Nelder JA (1983) Generalized linear models. Chapman and Hall, London
Meissner W (1998) Some notes on using walk-in traps. Wader Study Group Bull 86:33–35
Meissner W (2000) The wader station. In: Busse P (ed) Bird station manual. SE European Bird Migration Network, Gdansk University Press, Gdansk, pp. 89–102
Meissner W (2005) Variation in timing of the Siberian knot Calidris c. canutus autumn migration in the Puck Bay region (southern Baltic). Acta Ornithol 40:95–101
Meissner W (2007) To count or to catch: do walk-in traps lead to a biased measure of wader productivity? Wader Study Group Bull 113:53–56
Meissner W, Pilacka L (2008) Sex identification of adult dunlins Calidris alpina alpina migrating in autumn through Baltic region. Ornis Fennica 85:135–138
Meissner W, Sikora A (1995) Spring and autumn migration of waders (Charadrii) on the Hel Peninsula. Not Ornithol 36:205–239 (in Polish with English summary)
Meissner W, Skakuj M (2009) Ageing and sexing series 4: ageing and sexing the dunlin Calidris alpina migrating through Europe. Wader Study Group Bull 116:35–38
Meissner W, Strzałkowska M (2006) Autumn migration dynamics of the dunlin (Calidris alpina) at the Reda Mouth (southern Baltic). Ring 28:33–43
Meissner W, Włodarczak-Komosińska A, Górecki D, Wójcik C, Ściborski M, Krupa R, Zięcik P, Kozakiewicz M, Rydzkowski P, Remisiewicz M (2009) Autumn migration of waders (Charadrii) at the Reda mouth (N Poland). Ring 31:23–39
Meltofte H, Høye TT, Schmidt NM, Forchhammer MC (2007) Differences in food abundance cause inter-annual variation in the breeding phenology of High Arctic waders. Polar Biol 30:601–606
Minton C (2003) The importance of long-term monitoring of reproduction rates in waders. Wader Study Group Bull 100:178–182
Morrison RIG (1984) Migration systems of some New World shorebirds. In: Burger J, Olla B (eds). Behavior of marine animals, vol 6: shorebirds: migration and foraging behavior. Plenum, New York, pp 125–202
Naves LC, Lanctot RB, Taylor AR, Coutsoubos NP (2008) How often do Arctic shorebirds lay replacement clutches? Wader Study Group Bull 115:2–9
Ollason JC, Dunnet GM (1978) Age, experience and other factors affecting the breeding success of the Fulmar, Fulmarus glacialis, in Orkney. J Anim Ecol 47:961–976
Oring LW, Lank DB (1982) Sexual selection, arrival times, philopatry, and site-fidelity in the polyandrous spotted sandpiper. Behav Ecol Sociobiol 10:185–191
Oring LW, Reed JM, Colwell MA, Lank DB, Maxson SJ (1991) Factors regulating mating success and reproductive success in spotted sandpipers (Actitis macularia). Behav Ecol Sociobiol 28:433–442
Pettersson J (1994) Ottenby Fågelstationen med det stora kärrsnäppematerialet. Calidris 3:92–97
Pugesek BH (1983) The relationship between parental age and reproductive effort in the California Gull (Larus callifornicus). Behav Ecol Sociobiol 13:161–171
Sandercock BK (1998) Chronology of nesting events in western and semipalmated sandpipers near the Arctic circle. J Field Ornithol 69:235–243
Smith PA, Grant Gilchrist H, Forbes MR, Martin J-L, Allard K (2010) Inter-annual variation in the breeding chronology of arctic shorebirds: effects of weather, snow melt and predators. J Avian Biol 41:292–304
Soikkeli N (1967) Breeding cycle and population dynamics in the dunlin (Calidris alpina). Ann Zool Fenn 4:158–198
StatSoft Inc (2011) Statistica (data analysis software system), version 10. Available at: www.statsoft.com
Summers RW, Nicoll M (2004) Geographical variation in the breeding biology of the purple sandpiper Calidris maritima. Ibis 146:303–313
Summers RW, Underhill LG, Prys-Jones RP (1995) Why do young waders in southern Africa delay their first return migration to the breeding grounds? Ardea 83:351–357
Syroechkovski EE Jr, Lappo EG (1994) Migration phenology of waders (Charadrii) on the Taimyr Peninsula, northern Russia. Ostrich 65:181–190
Underhill LG, Prys-Jones RP, Syroechkovski EE, Groen NM, Karpov V, Lappo HG, van Roomen MWJ, Rybkin A, Schekkerman H, Summers RW (1993) Breeding of waders (Charadrii) and brent geese Branta bernicla at Pronchishcheva Lake, northeastern Taimyr, Russia, in a peak and a decreasing lemming year. Ibis 135:277–292
Zając R (1980) Different autumn migration rates of sexes in the dunlin Calidris a. alpina as determined by means of normal probability paper. Acta Ornithol 17:1–12
Acknowledgments
I would like to thank all ringers and volunteers working at ringing stations in Rewa and at the mouth of the Reda river. Special thanks to Agnieszka Ożarowska for improving the English and providing helpful comments on the manuscript, and to Jenny Gill and an anonymous reviewer for valuable comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by N. Chernetsov.
This is publication no. 149 of the Waterbird Research Group KULING.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 License (http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article

Cite this article
Meissner, W. Immature dunlins Calidris alpina migrate towards wintering grounds later than adults in years of low breeding success. J Ornithol 156, 47–53 (2015). https://doi.org/10.1007/s10336-014-1132-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-014-1132-y